
Аксонный транспорт в нервной системе
Аксонный транспорт. Быстрый аксонный транспорт. Медленный аксонный транспорт.
Процессы внутриклеточного транспорта наиболее ярко могут быть продемонстрированы на аксоне нервной клетки. Аксонный транспорт рассматривается здесь подробно, чтобы проиллюстрировать события, которые, вероятно, сходным образом происходят в большинстве клеток. Аксон, диаметр которого составляет всего несколько микронов, может достигать длины одного метра и более, и движение белков путем диффузии от ядра к дисталь-ному концу аксона заняло бы годы. Давно известно, что когда какой-либо из участков аксона подвергается констрикции, часть аксона, расположенная проксимальнее, расширяется. Это выглядит так, как будто в аксоне блокирован центробежный поток. Такой поток быстрый аксонный транспорт-может быть продемонстрирован движением радиоактивных маркеров, как в эксперименте, показанном на рис. 1.14. Лейцин, меченный радиоактивной меткой, инъецировали в ганглий дорсального корешка, и затем со 2-го по 10-й час измеряли радиоактивность в седалищном нерве на расстоянии 166 мм от тел нейронов. За 10 часов пик радиоактивности в месте инъекции менялся незначительно. Но волна радиоактивности распространялась по аксону с постоянной скоростью около 34 мм за 2 ч, или 410 мм/сут. Показано, что во всех нейронах гомойотермных животных быстрый аксонный транспорт осуществляется с такой же скоростью, причем ощутимых различий между тонкими, безмиелиновыми волокнами и наиболее толстыми аксонами, а также между моторными и сенсорными волокнами не наблюдается. Тип радиоактивного маркера также не влияет на скорость быстрого аксонного транспорта; маркерами могут служить разнообразные радиоактивные молекулы, такие, как различные аминокислоты, включающиеся в белки тела нейрона. Если проанализировать периферическую часть нерва, чтобы определить природу переносчиков транспортированной сюда радиоактивности, то такие переносчики обнаруживаются главным образом во фракции белков, но также в составе медиаторов и свободных аминокислот. Зная, что свойства этих веществ различны и особенно различны размеры их молекул, постоянную скорость транспорта мы можем объяснять только общим для всех них транспортным механизмом.
Быстрый аксонный транспорт может участвовать и в патологических процессах. Некоторые нейротропные вирусы (например, вирусы герпеса или полиомиелита) проникают в аксон на периферии и движутся с помощью ретроградного транспорта к телу нейрона, где размножаются и оказывают свое токсическое действие. Токсин столбняка-белок, который продуцируется бактериями, попадающими в организм при повреждениях кожи, захватывается нервными окончаниями и транспортируется к телу нейрона, где он вызывает характерные мышечные спазмы.
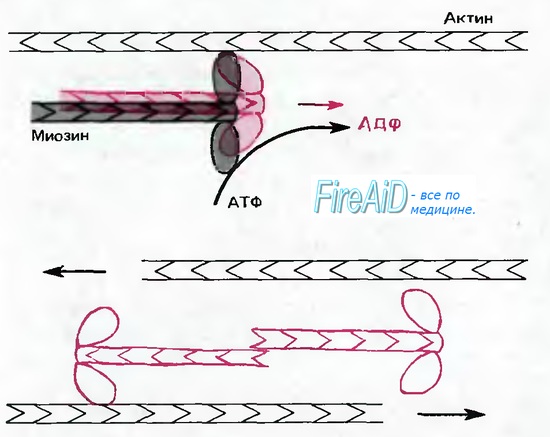
Рис. 1.13. Немышечный миозиновый комплекс при определенной ориентации может связываться с актиновыми филаментами различной полярности и, используя энергию АТФ, смещать их относительно друг друга
Помимо быстрого аксонного транспорта в клетке существует и довольно интенсивный медленный аксонный транспорт. Тубулин движется по аксону со скоростью около 1 мм/сут, а актин быстрее-до 5 мм/сут. С этими компонентами цитоскелета мигрируют и другие белки; например, ферменты, по-видимому, связаны с актином или тубулином. Скорости перемещения тубулина и актина примерно согласуются со скоростью роста, обнаруженной для механизма, описанного ранее, когда молекулы включаются в активный конец микротрубочки или мик-рофиламента. Следовательно, этот механизм может лежать в основе медленного аксонного транспорта. Скорость медленного аксонного транспорта примерно соответствует также скорости роста аксона, что, по-видимому, указывает на ограничения, накладываемые структурой цитоскелета на второй процесс.
Завершая данный раздел, следует подчеркнуть, что клетки ни в коем случае не являются статичными структурами, каковыми они кажутся, например, на электронно-микроскопических фотографиях. Плазматическая мембрана и особенно органеллы находятся в постоянном быстром движении и постоянной перестройке; только поэтому они способны функционировать. Далее, это не простые камеры, в которых протекают химические реакции, а высокоорганизованные конгломераты мембран и волокон, в которых реакции протекают в оптимально организованной последовательности.
Аксонный транспорт - это перемещение по аксону нервной клетки различного биологического материала. Аксональные отростки нейронов отвечают за передачу потенциала действия от тела нейрона к синапсу. Аксон представляет собой путь, по которому осуществляется транспорт необходимых биологических материалов между телом нейрона и синапсом, необходимый для функционирования нервной клетки. По аксону из области синтеза в теле нейрона транспортируются мембранные органел- лы (митохондрии), различные везикулы, сигнальные молекулы, ростовые факторы, белковые комплексы, компоненты цитоскелета и Na + - и К + - каналы. Конечными пунктами этого транспорта служат определенные области аксона и синаптической бляшки. В свою очередь, нейротрофические сигналы транспортируются из области синапса к телу клетки. Такова обратная связь, сообщающая о состоянии иннервации мишени.
По дендритам импульсы следуют к телу клетки, по аксону - от тела клетки к другим нейронам, мышцам или железам. Благодаря отросткам нейроны контактируют друг с другом и образуют нейронные сети и круги, по которым циркулируют нервные импульсы.
Специфическая функция аксона - проведение потенциала действия от тела клетки к другим клеткам или периферическим органам. Развитие аксона начинается с формирования конуса роста у нейрона. Конус роста проходит сквозь базальную мембрану, окружающую нервную трубку, и направляется через соединительную ткань зародыша к специфическим областям - мишеням. Конусы роста движутся по строго определенным путям, о чем свидетельствует точное подобие расположения нервов на обеих сторонах тела. Даже чужеродные аксоны, в экспериментальных условиях врастающие в конечность в местах нормальной иннервации, используют почти в точности тот же стандартный набор путей, по которым могут свободно передвигаться конусы роста. Очевидно, эти пути определяются внутренней структурой самой конечности, но молекулярная основа такой направляющей системы неизвестна. Видимо, по таким же предопределенным путям растут аксоны и в ЦНС, где эти пути, вероятно, определяются местными особенностями глиальных клеток эмбриона.
Классификация аксонного транспорта. Белки цитоскелета доставляются из тела клетки, двигаясь по аксону со скоростью от 1 до 5 мм в сутки. Это медленный аксонный транспорт (похожий на него транспорт имеется и в дендритах). Многие ферменты и другие белки цитозоля также переносятся при помощи этого типа транспорта. Медленный аксонный транспорт переносит крупные молекулы, в этом случае, сам транспортный механизм не является более медленным, но переносимые вещества время от времени попадают в клеточные компартаменты, не участвующие в транспорте. Так, митохондрии перемещаются иногда со скоростью быстрого транспорта, затем останавливаются или меняют направление движения, в результате получается медленный транспорт.
Нецитозольные материалы, которые необходимы в синапсе (секрети- руемые белки и мембраносвязанные молекулы), двигаются по аксону с гораздо большей скоростью. Эти вещества переносятся из места их синтеза, эндоплазматического ретикулума, к аппарату Гольджи, который часто располагается у основания аксона. Затем эти молекулы, упакованные в мембранные пузырьки, переносятся вдоль рельсов-микротрубочек путем быстрого аксонного транспорта со скоростью до 400 мм в сутки. Таким образом, по аксону транспортируются митохондрии, различные белки, включая нейропептиды (нейромедиаторы пептидной природы), непептидные нейромедиаторы.
Транспорт материалов от тела нейрона к синапсу называется антероградным, а в обратном направлении - ретроградным.
Транспорт по аксону на большие расстояния происходит с участием микротрубочек. Микротрубочки в аксоне обладают присущей им полярностью и ориентированы быстрорастущим (плюс) концом к синапсу, а медленнорастущим (минус) - к телу нейрона. Белки-моторы аксонного транспорта принадлежат к кинезиновому и динеиновому суперсемействам.
Кинезины являются плюс-концевыми моторными белками, транспортирующими такие грузы, как предшественники синаптических везикул и мембранные органеллы. Этот транспорт идет в направлении к синапсу (антероградно). Цитоплазматические динеины - это минус-концевые моторные белки, транспортирующие нейротрофные сигналы, эндосомы и другие грузы ретроградно к телу нейрона. Ретроградный транспорт осуществляется динеинами неэксклюзивно: обнаружены несколько кинези- нов, перемещающихся в ретроградном направлении.
5.2.5. АКСОННЫЙ ТРАНСПОРТ
Наличие у нейрона отростков, длина которых может достигать 1 м (например, аксоны, ин-нервирующие мускулатуру конечностей), создает серьезную проблему внутриклеточной связи между различными участками нейрона и ликвидации возможных повреждений его отростков. Основная масса веществ (структурных белков, ферментов, полисахаридов, липидов и др.) образуется в трофическом центре (теле) нейрона, расположенном преимущественно около ядра, а используются они в различных участках нейрона, включая его отростки. Хотя в аксонных окончаниях существуют синтез медиаторов, АТФ и повторное использование мембраны пузырьков после выделения медиатора, все же необходима постоянная доставка ферментов и фрагментов мембран из тела клетки. Для транспорта этих веществ (например, белков) путем диффузии на расстояние, равное максимальной длине аксона (около 1 м), потребовалось бы 50 лет! Для решения этой задачи эволюция сформировала специальный вид транспорта в пределах отростков нейрона, который более хорошо изучен в аксонах и получил название аксонного транспорта. С помощью этого процесса осуществляется трофическое влияние не только в пределах различных участков нейрона, но и на иннервируе-
мые клетки. В последнее время появились данные о существовании нейроплазматичес-кого транспорта в дендритах, который осуществляется из тела клетки со скоростью около 3 мм в сутки. Различают быстрый и медленный аксонный транспорт.
Быстрый аксонный транспорт осуществляется с помощью специальных структурных элементов нейрона: микротрубочек и микрофиламентов, часть которых представляет собой актиновые нити (актин составляет 10—15 % белков нейрона). Для транспорта необходима энергия АТФ. Разрушение микротрубочек (например, колхицином) и микрофиламентов (цитохолазином В), снижение уровня АТФ в аксоне более чем в 2 раза и падение концентрации Са 2+ блокируют аксонный транспорт.
рушается при разрушении микротрубочек, но прекращается при отделении аксона от тела нейрона, что свидетельствует о разных механизмах быстрого и медленного аксонного транспорта.
В. Функциональная роль аксонного транспорта. 1. Антеградный и ретроградный транспорт белков и других веществ необходимы для поддержания структуры и функции аксона и его пресинаптических окончаний, а также для таких процессов, как аксонный рост и образование синаптических контактов.
2. Аксонный транспорт участвует в трофическом влиянии нейрона на иннервируемую клетку, так как часть транспортируемых веществ выделяется в синаптическую щель и действует на рецепторы постсинаптической мембраны и близлежащих участков мембраны иннервируемой клетки. Эти вещества участвуют в регуляции обмена веществ, процессов размножения и дифференцировки ин-нервируемых клеток, формируя их функциональную специфику. Например, в опытах с перекрестной иннервацией быстрых и медленных мышц показано, что свойства мышц меняются в зависимости от типа иннервиру-ющего нейрона, его нейротрофического воздействия. Передатчики трофических влияний нейрона до сих пор точно не определены, важное значение в этом плане придается полипептидам и нуклеиновым кислотам.
3. Роль аксонного транспорта особенно ярко выявляется при повреждении нерва. Если нервное волокно на каком-либо участке прервано, его периферический отрезок, лишенный контакта с телом нейрона, подвергается разрушению, которое называется валле-ровской дегенерацией. В течение 2—3 сут наступает распад нейрофибрилл, митохондрий, миелина и синаптических окончаний. Надо отметить, что распаду подвергается участок волокна, снабжение которого кислородом и питательными веществами с кровотоком не прекращается. Считают, что решающим механизмом дегенерации является прекращение аксонного транспорта веществ от тела клетки до синаптических окончаний.
4. Аксонный транспорт играет важную роль и при регенерации нервных волокон.
Аксональный транспорт (аксоток) – это перемещение веществ от тела нейрона в отростки (антероградный аксоток) и в обратном направлении (ретроградный аксоток). Различают медленный аксональный ток веществ (1-5мм в сутки) и быстрый (до 1-5м в сутки). Обе транспортные системы присутствуют как в аксонах, так и в дендритах. Аксональный транспорт обеспечивает единство нейрона. Он создаёт постоянную связь между телом нейрона (трофическим центром) и отростками. Основные синтетические процессы идут в перикарионе. Здесь сосредоточены необходимые для этого органеллы. В отростках синтетические процессы протекают слабо.
Антероградная быстрая система транспортирует к нервным окончаниям белки и органеллы, необходимые для синаптических функций (митохондрии, фрагменты мембран, пузырьки, белки-ферменты, участвующие в обмене нейромедиаторов, а также предшественники нейромедиаторов). Ретроградная система возвращает в перикарион использованные и поврежденные мембраны и белки для деградации в лизосомах и обновления, приносит информацию о состоянии периферии, факторы роста нервов. Медленный транспорт – это антероградная система, проводящая белки и другие вещества для обновления аксоплазмы зрелых нейронов и обеспечения роста отростков при их развитии и регенерации.
Ретроградный транспорт может иметь значение в патологии. За счёт него нейротропные вирусы (герпеса, бешенства, полиомиелита) могут перемещаться с периферии в центральную нервную систему.
Нейроглия (глиоциты)
Глиоциты выполняют в нервной ткани вспомогательные функции: опорную, разграничительную, трофическую, секреторную и защитную. Они поддерживают постоянство среды вокруг нейронов. Клетки нейроглии делятся на две группы: макроглию и микроглию. Клетки макроглии бывают трёх типов: эпендимоциты, астроциты и олигодендро циты.
Эпендимоциты. Выстилают каналы и желудочки спинного и головного мозга, по которым циркулирует спинномозговая жидкость (ликвор). Эти клетки напоминают одно слойный призматический эпителий. На апикальных концах эпендимоцитов расположены реснички, помогающие движению спинномозговой жидкости. Через апикальные концы эпендимоциты могут выделять биологически активные вещества, которые с ликвором разносятся по всему мозгу. От ба зальных концов эпендимоцитов отходят отростки, которые могут идти через весь мозг. В желудочках мозга находятся сосудистые сплетения. Они покрыты специализированными эпен димо цитами, участвующими в образовании ликвора.
Астроциты. Различают протоплазматические и волокнистые астроциты. Протоплазматические астроциты имеют короткие толстые отростки. Они расположены в сером веществе мозга, выполняют разграничительную и трофическую функции. Волокнистые астроциты находятся в белом веществе, имеют многочисленные тонкие длинные от ростки, которые оплетают кровеносные сосуды мозга, образуя периваскулярные глиальные пограничные мембраны. Их отростки также изолируют синапсы. Таким образом, они изолируют нейроны и кровеносные сосуды и участвуют в образовании гемато-энцефалического барьера, обеспечивают обмен веществ между кровью и нейронами. Они также участвуют в образовании оболочек мозга и выполняют опорную функцию (образуют каркас мозга).
Олигодендроциты имеют мало отростков, окружают нейроны, выполняя трофическую (участие в пита нии нейронов) и разграничительную функции. Олигодендроциты, расположенные вокруг тел нейронов, называются мантийными глиоцитами. Олигодендроциты, расположенные в периферической нервной системе и образующие оболочки вокруг отростков нейронов, называют леммоцитами (шванновскими клетками).
Источники развития – нервная трубка, нервный гребень (ганглиозные пластинки) и плакоды. Нервная трубка образуется в результате смыкания краёв нервного желобка, развивающегося из эктодермы. Нервная трубка состоит из трёх концентрических слоёв клеток: внутренний –вентрикулярный (эпендимный), средний – мантийный и наружный – краевая вуаль. В центре нервной трубки находится полость, которая сохраняется у взрослых в виде каналов и желудочков мозга, по которым циркулирует спинномозговая жидкость. Вентрикулярный слой состоит из делящихся матричных клеток. Часть клеток из вентрикулярного слоя мигрирует в мантийный слой, где превращается в клетки двух типов: нейробластыи глиобласты(спонгиобласты). Нейробласты в процессе дифференцировки превращаются в нейроны, а спонгиобласты – в глиальные клетки. Нейробласты постепенно утрачивают способность к делению, приобретают отростки и органоиды специального назначения (нейрофибриллы) и дифференцируются в нейроны. Отростки нейронов прорастают в кравевую вуаль и образуют белое вещество нервной системы. Растущий аксон нейрона имеет конус роста, состоящийиз булавовидного утолщения, от которого отходят тонкие пальцевидные выросты – филоподии. Он растёт со скоростью 1 мм/сутки в сторону иннервируемого им органа-мишени, навстречу выделяемым этим органом химическим веществам (хемотаксис).
Нервные гребни расположены между нервной трубкой и эктодермой. Они образуются в результате выселения клеток из утолщенных краев нервного желобка – нервных валиков. Плакоды представляют собой утолщения эктодермы по бокам нервной трубки на головном конце зародыша. Нейробласты нервной трубки дают начало нервным клеткам, а глиобласты – глиальным клеткам головного и спинного мозга. Из клеток нервного гребня происходят нейроны и нейроглия всех нервных ганглиев, а из пла код – рецепторные (нейросенсорные) клетки органа обоняния, нейроны слухового и вестибулярного ганглиев. Клетки микроглии образуются из моноцитов крови.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Волокна группы А альфа
(диаметр -13-22 мкм, скорость – 60- 120 м/с, длительность ПД- 0,4-0,5 мс)
1). эфферентные волокна, проводящие
возбуждение к скелетным мышцам от альфа-мотонейронов
2) афферентные волокна, проводящие возбуждение от мышечных рецепторов в ЦНС
Волокна группы А бета
(диаметр – 8-13- мкм, скорость – 40- 70 м/с, длительность ПД- 0,4 -0,6 мс)
1. Афферентные волокна, проводящие
возбуждение от рецепторов прикосновения и сухожильных рецепторов в ЦНС
Волокна группы А гамма
(диаметр – 4-8 мкм, скорость – 15- 40 м/с, длительность ПД- 0,5 -0,7 мс)
1) эфферентные волокна к мышечным веретенам от гамма-мотонейрнов
2). афферентные волокна, проводящие
возбуждение от рецепторов прикосновения и давления в ЦНС
Волокна группы В
(диаметр – 1-3 мкм, скорость -3-14 м/с, длительность ПД- 1,2 мс)
Это – преганглионарные волокна вегетативной нервной системы
Волокна группы С
(диаметр – 0,5—1,0 мкм, скорость -0,5-2,0 м/с, длительность ПД- 2,0 мс)
1.постганглионарные волокна ВНС
2.афферентные волокна, проводящие возбуждение от рецепторов боли, давления и тепла в ЦНС
Аксонный транспорт. Быстрый аксонный транспорт. Медленный аксонный транспорт.
Аксонный транспорт - это перемещение веществ по аксону . Белки, синтезированные в теле клетки, синаптические медиаторные вещества и низкомолекулярные соединения перемещаются по аксону вместе с клеточными органеллами, в частности, митохондриями . Для большинства веществ и органелл обнаружен также транспорт в обратном направлении. Вирусы и токсинымогут проникать в аксон на его периферии и перемещаться по нему. Аксонный транспорт - активный процесс. Различают
быстрый аксонный транспорт и медленный аксонный транспорт .
Медленный аксонный транспорт - транспорт крупных молекул, в этом случае, видимо, сам транспортный механизм не является более медленным, но переносимые вещества время от времени попадают в клеточные компартаменты, не участвующие в транспорте. Так, митохондрии перемещаются иногда со скоростью быстрого транспорта, затем останавливаются или меняют направление движения, в результате получается медленный транспорт.
Скорость быстрого аксонного транспорта составляет 410 мм/день. Такая скорость обнаружена во всех нейронах теплокровных животных, независимо от вида переносимых молекул.
Во многих случаях транспорт органелл в клетке зависит от микротрубочек Микротрубочки в аксоне характеризуются относительной стабильностью по сравнению с другими клетками. Вероятно, это происходит за счет высокого содержания MAP , которые способны стабилизировать микротрубочки. Кроме того, этому способствует образование пучков микротрубочек с помощью различных ассоциированных белков .
Выделяют два основных вида транспорта: прямой (антероградный) – от тела клетки по отросткам к их периферии и обратный (ретроградный) – по отросткам нейрона к телу клетки
В нейроне, как и в других клетках организма, постоянно происходят процессы распада молекул, органоидов, других компонентов клетки. Их необходимо постоянно обновлять. Нейроплазматический транспорт важен для обеспечения электрических и неэлектрических функций нейрона, для осуществления обратной связи между отростками и телом нейрона. При повреждении нервов необходима регенерация поврежденных участков и восстановление иннервации органов.
Разнообразные вещества транспортируются по отросткам нейрона с разной скоростью, в разных направлениях и с использованием разных механизмов транспорта. Выделяют два основных вида транспорта: прямой (антероградный) – от тела клетки по отросткам к их периферии и обратный (ретроградный) – по отросткам нейрона к телу клетки (табл. 1).
Механизмы аксонного и дендритного транспорта
Прямой аксональный транспорт осуществляют моторные молекулы, связанные с системой цитоскелета и плазматической мембраной. Моторная часть молекул кинезина или денеина связывается с микротрубочкой, а хвостовая ее часть – с транспортируемым материалом, с аксональной мембраной или с соседними элементами цитоскелета. В обеспечении транспорта по отросткам принимают участие и ряд вспомогательных белков (адапторов), ассоциированных с кинезином или денеином. Все процессы идут со значительной затратой энергии.
Обратный (ретроградный) транспорт.
В аксонах основным механизмом обратного транспорта является система денеиновых и миозиновых моторных белков. Морфологическим субстратом этого транспорта являются: в аксоне – мультивезикулярные тела и сигнальные эндосомы, в дендритах – мультивезикулярные и мультиламеллярные тела.
Большую роль в транспортных процессах в нейроне играет гладкий эндоплазматический ретикулум. Показано, что по всей длине отростков нейрона распространяется непрерывная разветвленная сеть цистерн гладкого ретикулума. Концевые ветвления этой сети проникают в пресинаптические участки синапсов, где от них отшнуровываются синаптические пузырьки. Именно по его цистернам быстро транспортируются многие медиаторы и нейромодуляторы, нейросекреты, ферменты их синтеза и распада, ионы кальция и другие компоненты аксотока. Молекулярные механизмы этой разновидности транспорта пока не ясны.
Аксональный транспорт
Мембранные и цитоплазматические компоненты, которые образуются в биосинтезирующем аппарате сомы и проксимальной части дендритов , должны распределяться по аксону (особенно важно их поступление в пресинаптические структуры синапсов ), чтобы восполнить потерю элементов, подвергшихся высвобождению или инактивации. Однако многие аксоны слишком длинны, чтобы материалы могли эффективно перемещаться из сомы к синаптическим окончаниям путем простой диффузии. Эту задачу выполняет особый механизм - аксональный транспорт.
Существует несколько его типов. Окруженные мембранами органоиды и митохондрии транспортируются с относительно большой скоростью посредством быстрого аксонального транспорта. Вещества, растворенные в цитоплазме (например, белки), перемещаются с помощью медленного аксонального транспорта. У млекопитающих быстрый аксональный транспорт обладает скоростью 400 мм/сут, а медленный - около 1 мм/сут. Синаптические пузырьки могут передвигаться с номощью быстрого аксонального транспорта из сомы мотонейрона спинного мозга человека к нервно-мышечному соединению стопы примерно за 2,5 сут. Сравним: доставка на такое же расстояние многих растворимых белков происходит примерно за 3 г.
Для работы аксонального транспорта требуются затрата метаболической энергии и присутствие внутриклеточного Са2+. Элементы цитоскелета (точнее, микротрубочки ) создают систему направляющих тяжей, вдоль которых передвигаются окруженные мембранами органоиды ( рис. 32.13 ). Эти органоиды прикрепляются к микротрубочкам аналогично тому, как это происходит между толстыми и тонкими филаментами волокон скелетных мышц ; движение органоидов вдоль микротрубочек запускается ионами Са2+.
Аксональный транспорт осуществляется в двух направлениях. Транспорт от сомы к аксональным терминалям, называемый антероградным аксональным транспортом ( рис. 32.14 , а), восполняет в пресинаптических окончаниях запас синаптических пузырьков и ферментов, ответственных за синтез нейромедиатора. Транспорт в противоположном направлении - ретроградный аксональный транспорт ( рис. 32.14 , б), возвращает опустошенные синаптические пузырьки в сому, где эти мембранные структуры деградируются лизосомами .
Посредством аксонального транспорта по периферическим нервам распространяются некоторые вирусы и токсины . Так, вирус, который может вызывать ветряную оспу ( varicella-zoster virus ), проникает в клетки спинальных ганглиев . Там он пребывает в неактивной форме иногда в течение многих лет, пока не изменится иммунный статус человека. Тогда вирус может транспортироваться по сенсорным аксонам к коже, и в дерматомах соответствующих спинальных нервов возникают болезненные высыпания - опоясывающий лишай (Herpes zoster) . Путем аксонального транспорта также переносится столбнячный токсин . Бактерии Clostridium tetani могут оказаться в загрязненной ране и, если человек не был вакцинирован, попасть путем ретроградного транспорта в мотонейроны. Если токсин выйдет во внеклеточное пространство передних рогов спинного мозга , он заблокирует активность синаптических рецепторов тормозных нейромедиаторных аминокислот и станет причиной тетанических судорог .
Аксон (греч. ἀξον — ось) — нейрит, осевой цилиндр, отросток нервной клетки, по которому нервные импульсы идут от тела клетки (сомы) к иннервируемым органам и другим нервным клеткам.
Нейрон состоит из одного аксона, тела и нескольких дендритов, в зависимости от числа которых нервные клетки делятся на униполярные, биполярные, мультиполярные. Передача нервного импульса происходит от дендритов (или от тела клетки) к аксону, а затем сгенерированный потенциал действия от начального сегмента аксона передается назад к дендритам [1]. Если аксон в нервной ткани соединяется с телом следующей нервной клетки, такой контакт называется аксо-соматическим, с дендритами — аксо-дендритический, с другим аксоном — аксо-аксональный (редкий тип соединения, встречается в ЦНС).
В месте соединения аксона с телом нейрона у наиболее крупных пирамидных клеток 5-ого слоя коры находится аксонный холмик. Ранее предполагалось, что здесь происходит преобразование постсинаптического потенциала нейрона в нервные импульсы, но экспериментальные данные это не подтвердили. Регистрация электрических потенциалов выявила, что нервный импульс генерируется в самом аксоне, а именно в начальном сегменте на расстоянии
50 мкм от тела нейрона [2]. Для генерации потенциала действия в начальном сегменте аксона требуется повышенная концентрация натриевых каналов (до ста раз по сравнению с телом нейрона[3]).
В протоплазме аксона — аксоплазме — имеются тончайшие волоконца — нейрофибриллы, а также микротрубочки, митохондрии и агранулярная (гладкая) эндоплазматическая сеть. В зависимости от того, покрыты ли аксоны миелиновой (мякотной) оболочкой или лишены её, они образуют мякотные или безмякотные нервные волокна.
Концевые участки аксона — терминали — ветвятся и контактируют с другими нервными, мышечными или железистыми клетками. На конце аксона находится синаптическое окончание — концевой участок терминали, контактирующий с клеткой-мишенью. Вместе с постсинаптической мембраной клетки-мишени синаптическое окончание образует синапс. Через синапсы передаётся возбуждение.
Аксо́нный тра́нспорт — это перемещение по аксону нервной клетки различного биологического материала.
Аксональные отростки нейронов отвечают за передачу потенциала действия от тела нейрона к синапсу. Также аксон представляет собой путь, по которому осуществляется транспорт необходимых биологических материалов между телом нейрона и синапсом, необходимый для функционирования нервной клетки. По аксону из области синтеза в теле нейрона транспортируются мембранные органеллы (митохондрии), различные везикулы, сигнальные молекулы, ростовые факторы, белковые комплексы, компоненты цитоскелета и даже Na+- и K+-каналы. Конечными пунктами этого транспорта служат определенные области аксона и синаптической бляшки. В свою очередь, нейротрофические сигналы транспортируются из области синапса к телу клетки. Это выполняет роль обратной связи, сообщающей о состоянии иннервации мишени.Длина аксона периферической нервной системы человека может превышать 1 м, а может быть и больше у крупных животных. Толщина большого мотонейрона человека составляет 15 мкм, что при длине в 1 м дает объём
0,2 мм³, а это почти в 10000 раз больше объёма клетки печени. Это делает нейроны зависимыми от эффективного и координированного физического транспорта веществ и органелл по аксонам.
Величины длин и диаметров аксонов, а также количества материала, транспортируемого по ним, безусловно, говорят о возможности возникновения сбоев и ошибок в системе транспорта. Многие нейродегенеративные заболевания непосредственно связаны с нарушениями в работе этой системы.
Белки цитоскелета доставляются из тела клетки, двигаясь по аксону со скоростью от 1 до 5 мм в сутки. Это медленный аксонный транспорт (похожий на него транспорт имеется и в дендритах). Многие ферменты и другие белки цитозоля также переносятся при помощи этого типа транспорта.Нецитозольные материалы, которые необходимы в синапсе, такие как секретируемые белки и мембраносвязанные молекулы, двигаются по аксону с гораздо большей скоростью. Эти вещества переносятся из места их синтеза, эндоплазматического ретикулума, к аппарату Гольджи, который часто располагается у основания аксона. Затем эти молекулы, упакованные в мембранные пузырьки, переносятся вдоль рельсов-микротрубочек путем быстрого аксонного транспорта со скоростью до 400 мм в сутки. Таким образом по аксону транспортируются митохондрии, различные белки, включая нейропептиды (нейромедиаторы пептидной природы), непептидные нейромедиаторы.Транспорт материалов от тела нейрона к синапсу называется антероградным, а в обратном направлении — ретроградным.Транспорт по аксону на большие расстояния происходит с участием микротрубочек. Микротрубочки в аксоне обладают присущей им полярностью и ориентированны быстрорастущим (плюс-)концом к синапсу, а медленнорастущим (минус-) — к телу нейрона. Белки-моторы аксонного транспорта принадлежат к кинезиновому и динеиновому суперсемействам.Кинезины являются, в основном, плюс-концевыми моторными белка́ми, транспортирующими такие грузы, как предшественники синаптических везикул и мембранные органеллы. Этот транспорт идет в направлению к синапсу (антероградно). Цитоплазматические динеины — это минус-концевые моторные белки, транспортирующие нейротрофные сигналы, эндосомы и другие грузы ретроградно к телу нейрона. Ретроградный транспорт осуществляется динеинами не эксклюзивно: обнаружены несколько кинезинов, перемещающихся в ретроградном направлении.
11.Миелинизированные и немиелинизированные волокна. Процесс миелинизации. Бол-во нервов содержит миелинизированные и немиелинизированные или слабо миелинизированные волокна. Клеточный состав эндоневральных пространств отражает уровень миелинизации. В норме 90% обнаруживаемых в этом пространстве клеточных ядер относится к клеткам Шванна (леммоцитам), а остальные принадлежат фибробластам и капилярному эндотелию. При 80% шванновских клеток окружают немиелинизированных аксоны; рядом с миелинизированными волокнами их количество уменьшено в 4 раза. Миелинизированные волокна большого диаметра проводят импульсы в значительно более быстром темпе, чем слабо миелинизированные или немиелинизированные. Выделяют три класса волокон: А, В и С. А-волокна – соматические афферентные и афферентные миелинизированные нервные волокна, В-волокна – миелинизированные преганглионарные вегетативные волокна, С-волокна – немиелинизированные вегетативные и сенсорные волокна. Миелин покрывает оболочку нервных стволов и обеспечивает более эффективную передачу нервного импульса. Процесс называется миелинизацией, так как в результате образуется чехол из вещества миелина, примерно на 2/3 состоящего из жира и являющегося хорошим электрическим изолятором. Исследователи придают очень большое значение процессу миелинизации в развитии мозга. Известно, что у новорожденного ребенка миелинизировано примерно 2/3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций. Оказывается, миелинизированные волокна в сотни раз быстрее проводят возбуждение, чем немиелинизированные, т. е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно.
12.Межнейрональные связи. Синапсы, их строение и функции. На более поздних стадиях филогенеза и прежде всего у человека связь между нервными клетками осуществляется специальными образованиями — синапсами. Синапс состоит из трех основных элементов: пресинаптической мембраны, синаптической щели и постсинаптической мембраны. Пресинаптическая мембрана представляет собой нейросекреторный аппарат, в котором синтезируется и выделяется медиатор, оказывающий тормозящее или возбуждающее действие на постсинаптическую мембрану иннервируемой клетки. Постсинаптическая мембрана обладает избирательной чувствительность к химическому агенту — медиатору и практически нечувствительна к раздражителю электрическим током. Наличие синапсов определяет одностороннее проведение нервного импульса (обратная передача возбуждения с постсинаптнческой на пресинаптическую мембрану невозможна), поскольку в нервных волокнах возбуждение может распространяться в обе стороны от стимула. Вместе с тем в синапсе замедляется скорость проведения. Длительность синаптической задержки варьирует в значительных пределах в зависимости от функционального назначения синапса и составляет 0,2 — 0,5 мс в межнейрональных и нервно- мышечных синапсах, тогда как в нервных окончаниях гладкой мускулатуры достигает 5 — 10 мс.
13.Типы синапсов (химические и электрические). Механизм синаптической передачи. Медиатор, находящийся в пузырьках, выделяется в синаптическую щель с помощью экзоцитоза (пузырьки подходят к мембране, сливаются с ней и разрываются, выпуская медиатор). Его выделение происходит небольшими порциями – квантами. Каждый квант содержит от 1.000 до 10.000 молекул нейромедиатора. Небольшое количество квантов выходит из окончания и в состоянии покоя. Когда нервный импульс, т.е. ПД, достигает пресинаптического окончания, происходит деполяризация его пресинаптической мембраны. Открываются ее кальциевые каналы и ионы кальция входят в синаптическую бляшку. Начинается выделение большого количества квантов нейромедиатора. Молекулы медиатора диффундируют через синаптическую щель к постсинаптической мембране и взаимодействуют с ее хеморецепторами. В результате образования комплексов медиатор-рецептор, в субсинаптической мембране начинается синтез так называемых вторичных посредников. В частности цАМФ. Эти посредники активируют ионные каналы постсинаптической мембраны. Поэтому такие каналы называют хемозависимыми или рецепторуправляемыми. Т.е. они открываются при действии ФАВ на хеморецепторы. В результате открывания каналов изменяется потенциал субсинаптической мембраны. Такое изменение называется постсинаптическим потенциалом. Электрические синапсы. представляет собой щелевидное образование (размеры щели до 2 нм) с ионными мостиками-каналами между двумя контактирующими клетками. Петли тока, в частности при наличии потенциала действия (ПД), почти беспрепятственно перескакивают через такой щелевидный контакт и возбуждают, т.е. индуцируют генерацию ПД второй клетки. В целом, такие синапсы (они называются эфапсами) обеспечивают очень быструю передачу возбуждения. Но в то же время с помощью этих синапсов нельзя обеспечить одностороннее проведение, т. к. большая часть таких синапсов обладает двусторонней проводимостью. Кроме того, с их помощью нельзя заставить эффекторную клетку (клетку, которая управляется через данный синапс) тормозить свою активность. Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются щелевые контакты типа нексуса. Химические синапсы. По строению химические синапсы представляют собой окончания аксона (терминальные синапсы) или его варикозную часть (проходящие синапсы), которая заполнена химическим веществом - медиатором. В синапсе различают пресинаптический элемент, который ограничен пресинаптической мембраной, постсинаптический элемент, который ограничен постсипаптической мембраной, а также внесинаптическую область и синаптическую щель, величина которой составляет в среднем 50 нм. В литературе существует большое разнообразие в названиях синапсов. Например, синаптическая бляшка - это синапс между нейронами, концевая пластинка - это постсинаптическая мембрана мионеврального синапса, моторная бляшка - это пресинаптичсское окончание аксона на мышечном волокне.
Дата добавления: 2015-02-16 ; просмотров: 32 | Нарушение авторских прав
Читайте также:
