
Анализатор высшей нервной деятельности
Физиология сенсорных систем
Аналитико-синтетическая деятельность мозга. Две составные части учения И.П.Павлова о физиологии высшей нервной деятельности: об анализаторах и условных рефлексах. Синтез-анализатор. Единство аналитической и синтетической деятельности головного мозга. Анализ и синтез сложных раздражителей. Условнорефлекторная деятельность и механизм высшего анализа и синтеза.
Методы исследования физиологии высшей нервной деятельности: метод условных рефлексов, полиграфическая регистрация реакций, электроэнцефалография, магнитоэнцефалография, термоэнцефалография, позитронно-эмиссионная томография, функциональная магнитно-резонансная томография, оптическая регистрация активности нейронов, экстраклеточная и внутриклеточная регистрация, электрическое раздражение мозга, экстирпация и функциональное выключение участков мозга, исследования в онтогенезе и филогенезе, клинический метод, метод моделирования. Автоматизация экспериментов с применением компьютера.
Методы измерения порогов. Рабочая характеристика приемника. Ранжирование раздражителей.Принципы переработки информации в нервной системе. Кодирование стимула номером канала. Нейроны-детекторы. Командные нейроны. Модулирующие нейроны. Гностические нейроны. Концептуальная рефлекторная дуга.
Общие принципы работы анализаторов. Структура и функция анализатора. Многоуровневость анализаторных систем. Понятия уровня, нейронного слоя и проводящей части анализатора. Прямые афферентные пути и ассоциативные системы. Параллельные нейронные каналы, их множественность. Иерархический принцип построения анализаторов. Неспецифические влияния ретикулярной формации.
Рецепторы. Рецепторный потенциал. Адаптация. Рецептивное поле нейронов. Латеральное торможение.Глаз мечехвоста как модель изучения механизма и функции латерального торможения. Латеральное торможение в анализаторах. Роль возвратного торможения и афферентного коллатерального торможения в переработке сенсорных сигналов. Обратная афферентация.
Восходящие пути анализатора. Корковый конец анализатора. Слои коры. Колончатая организация коры. Проекционные области коры. Вызванный потенциал мозга. Нисходящие влияния анализаторов. Двигательные компоненты анализаторов.
Зрительный анализатор. Фоторецепторы: палочки и колбочки. Зрительные пигменты. Квантовые механизмы фоторецепции. Ранний и поздний рецепторный потенциал. Строение сетчатки. Свойство биполярных, горизонтальных, амакриновых и ганглиозных клеток. Роль глиальных клеток. Электроретинограмма и ее анализ. Фотопическое и скотопическое зрение. Световая и темновая адаптация. Сдвиг Пуркинье.
Оптическая система глаза. Нормальная и аномальная рефракция. Острота зрения. Зрачковый рефлекс. Аккомодация. Глазодвигательный аппарат глаза. Саккады. Следящие движения. Компенсаторные движения. Вергентные движения. Фиксация. Оптокинетический нистагм. Вестибуляторный нистагм. Электроокулография. Нейронные механизмы движений глаз.
Организация рецептивных полей ганглиозных клеток сетчатки. Концентрические рецептивные поля ганглиозных клеток сетчатки. Ганглиозные клетки, реагирующие на изменение стимула. Рецептивные поля наружного коленчатого тела. Простые, сложные и гиперсложные рецептивные поля нейронов зрительной коры. Ретинотопическая проекция. Слоистое строение зрительной коры. Организация нейронов зрительной коры в микро-, макро- и гиперколонки.
Восприятие формы. Нейроны, избирательные к ориентации элементов изображения. Восприятие движения. Детекторы, селективные к скорости и направлению движения.
Стереоскопическое зрение. Проекции двух сетчаток на наружные коленчатые тела и зрительную кору мозга. Корреспондирующие и диспаратные точки сетчатки. Нейроны, селективные в отношении степени диспаратности. Восприятие удаленности. Нейронные механизмы константности восприятия пространства.
Цветовое зрение. Трехкомпонентная теория цветового зрения. Цветоприемники сетчатки. Оппонентные нейроны. Рецептивные поля цветоизбирательных нейронов. Детекторы цвета. Правила сложения цветов. Сферическая модель цветоразличения. Нарушения цветового зрения.
Слуховой анализатор. Строение наружного, среднего и внутреннего уха. Кортиев орган. Волосковые клетки. Механизм возбуждения волосковых клеток. Микрофонный эффект улитки. Нейроны спирального ганглия. Частотно-пороговые кривые волокон слухового нерва. Характеристическая частота. Функция латерального торможения. Кодирование частоты и интенсивности звуковых сигналов. Тонотопическая проекция. Восходящие и нисходящие пути.
Нейроны кохлеарных ядер. Нейроны внутреннего коленчатого тела. Нейроны слуховой коры. Нейроны, селективные в отношении звуковых комплексов. Нейроны, избирательные в отношении скорости и направления модуляции по частоте и амплитуде. Нейронные механизмы фонематического слуха.
Бинауральный слух. Бинауральная разность фаз и интенсивностей как факторы локализации звука. Бинауральные нейроны верхней оливы. Детекторы направления звука в заднем двухолмии. Детекторы движения источника звука в пространстве. Нейронные механизмы эхолокации летучих мышей.
Вестибулярный анализатор. Строение и функция вестибулярного анализатора. Оттолитовый аппарат. Статоцист как прототип оттолитового прибора. Саккулюс и утрикулюс. Нейронные механизмы кодирования направления вектора силы тяжести. Полукружные каналы. Рецепторы полукружных каналов. Нейроны мозжечка. Нейронные механизмы кодирования ускорений. Нейронные механизмы компенсаторных движений глаз. Нейронные механизмы поддержания позы.
Кожный и двигательный анализатор. Рецепторные образования кожи. Тельца Паччини, Мейснера, диски Меркеля, свободные нервные окончания. Мышечное веретено. Рецептор растяжения рака как модель механорецептора.
Тактильная, температурная, проприоцептивная, вибрационная, болевая чувствительность. Структура соматосенсорного анализатора. Нейронная организация спинного мозга. Соматотопическая проекция. Таламические нейроны соматосенсорной системы. Колончатая организация соматосенсорной коры.
Вкусовой анализатор. Вкусовые рецепторы. Вкусовые луковицы. Вкусовые волокна барабанной струны и языкоглоточного нерва. Нейроны ядра одиночного пучка. Функция нейронов таламуса в детекции вкуса. Реакции нейронов гипоталамуса на вкусовые стимулы. Функция коры. Нейронные механизмы голода и жажды.
Обонятельный анализатор. Обонятельный эпителий. Электроольфактограмма. Обонятельные рецепторы. Обонятельные луковицы. Митральные клетки, кисточковые клетки. Обонятельный тракт. Первичная обонятельная кора. Нейронные механизмы кодирования запахов. Реакции нейронов гипоталамуса. Участие нейронов обонятельной системы в рефлекторном поведении.
Интероцепция. Рецепторы внутренних органов. Барорецепторы.Глюкорецепторы. Осморецепторы. Кора и внутренние органы.
- Адам Д. Восприятие, сознание, память. М.: Мир, 1983. С. 9—76.
- Батуев А.С., Куликов Г.А. Введение в физиологию сенсорных систем. М.: Изд-во Моск. ун-та, 1983. С. 4—136.
- Наатенен Р. Внимание и функции мозга. М.: Изд-во Моск. ун-та, 1997. 559 с.
- Основы сенсорной физиологии / Под ред. Р.М.Шмидта. М.: Мир, 1984. 287 с.
- Павлов И.П. Лекции о работе больших полушарий головного мозга.М.: Изд-во АМН СССР, 1952. С.73—98.
- Соколов Е.Н. Физиология высшей нервной деятельности.М.: Изд-во Моск. ун-та, 1974. Ч.1. С. 3—211.
- Физиология сенсорных систем. Л.: Наука, 1971. Ч.1. С. 88—111, 126—149, 246—299; Ч. 2. С. 130—137, 158—191, 299—307.
- Хьюбел Д. Глаз, мозг, зрение. М.: Мир, 1990. 239 с.
- Шеперд Г. Нейробиология: В 2 т. М.: Мир, 1987. Т.1. 368 с.;
Т.2. 454 с.
Физиология высшей нервной деятельности
Основные этапы формирования взглядов о поведении, функциях нервной системы и мозга. История развития понятия рефлекс: Р. Декарт, Й. Прохазка, И.М. Сеченов, И.П. Павлов. Предмет физиологии высшей нервной деятельности. Эволюционный подход к исследованию высшей нервной деятельности: взгляды Ч.Дарвина, И.П.Павлова, Л.А.Орбели. Физиология высшей нервной деятельности как составная часть нейронауки. Методы исследования физиологии высшей нервной деятельности.
Поведение, научение и память. Функциональная пластичность нервной ткани. Определение пластичности. Механизмы пластичности. Роль генома в пластических изменениях нервной ткани.
Рефлекторная основа поведения. Классификация врожденных форм поведения. Таксисы, безусловные рефлексы, инстинктивные формы поведения. Изменчивость врожденного поведения. Пластичность врожденного поведения. Ориентировочный рефлекс, его структура и нейронные механизмы. Нервная модель стимула. Угасание реакций. Гиппокамп и ориентировочный рефлекс. Физиологические механизмы внимания. Ориентировочно-исследовательская деятельность.
Научение. Классификация форм научения. Неассоциативные формы научения. Привыкание, его механизмы. Сенситизация, ее механизмы. Двухпроцессная теория привыкания. Посттетаническая потенциация.
Ассоциативное научение. Первая сигнальная система. Классический условный рефлекс. Виды классических условных рефлексов: пищевые, оборонительные, двигательные, вегетативные. Инструментальные условные рефлексы. Стадии генерализации и специализации условных рефлексов. Экстероцептивные, интероцептивные и проприоцептивные условные рефлексы. Правила образования условных рефлексов. Торможение условных рефлексов. Виды торможения. Механизмы внутреннего торможения. Сложные формы научения.Условные рефлексы на комплексные раздражители, на время, цепные, порядковые, подражательные и экстраполяционные. Импринтинг, его механизмы. Латентное научение. Психонервная деятельность.
Механизмы образования временной связи. Доминантный очаг. Нейрофизиологические механизмы доминанты. Дистантная синхронизация биопотенциалов головного мозга. Фазическая и тоническая активность мозга при выработке временной связи. Нейронные механизмы замыкания временных связей. Синапс Хебба. Роль постсинаптических рецепторов и экспрессии генов в замыкании временной связи.
Механизмы памяти. Виды памяти. Электрофизиологические корреляты памяти. Процессная негативность. Префронтальная кора и память. Гиппокамп и память. Молекулярные основы памяти.
Функциональные состояния, потребности, мотивации и эмоции. Функциональные состояния и модулирующие системы мозга. Нейрофизиологические механизмы функциональных состояний. Физиологические индикаторы функциональных состояний. Функциональное состояние и поведение. Уровни бодрствования. Сон. Структура состояния сна. Нейрофизиологические механизмы различных фаз сна. Теории сна. Сновидения. Условнорефлекторная деятельность во время сна. Патологические формы сна. Гипноз и его механизмы.
Потребности, мотивации и эмоции. Классификация потребностей. Роль критических периодов в формировании потребностей. Виды мотиваций. Мотивация как доминанта. ЭЭГ-корреляты мотивационных состояний. Нейроанатомия и нейрохимия мотиваций. Происхождение эмоций. Потребности и эмоции. Информационная теория эмоций. Мотивации и эмоции. Выражение эмоций. Нейроанатомия и нейрохимия эмоций. Эмоциональный анализатор. Семантическое эмоциональное пространство.
Интегративная деятельность мозга. Уровни интегративной деятельности (Ч. Шеррингтон). Концептуальная рефлекторная дуга (Е.Н.Соколов). Структура поведенческого акта — функциональная система П.К.Анохина. Механизмы управления движением: принцип сенсорных коррекций, принцип прямого программного управления движением. Спинальные генераторы локомоций. Центральные моторные программы. Принцип обратной связи в деятельности мозга. Механизмы непроизвольных и произвольных движений.
Высшие психические функции: речь и сознание. Вторая сигнальная система. Нейросемантический код. Речевые функции полушарий мозга. Развитие речи у ребенка. Нейронные механизмы восприятия и генерации речи. Взаимодействие первой и второй сигнальных систем. Метод позитронно-эмиссионной томографии и магнитного резонанса в исследовании речевых функций мозга. Определение сознания. Прожекторная теория сознания.
Индивидуальные различия высшей нервной деятельности. Теории индивидуальности. Общие типы высшей нервной деятельности (И.П.Павлов). Свойства нервной системы и индивидуальные различия. Генотип и фенотип в проявлениях высшей нервной деятельности человека. Темперамент в структуре индивидуальности. Биохимические переменные в исследованиях индивидуальных различий.
Основная
- Батуев А.С. Высшая нервная деятельность. М.: Высш. шк., 1991. 396 с.
- Данилова Н.Н., Крылова А.Л. Физиология высшей нервной деятельности. М.: Учебная лит-ра, 1997. 431 с.
- Данилова Н.Н. Психофизиологическая диагностика функциональных состояний. М.: Изд-во Моск. ун-та, 1992. 191 с.
- Павлов И.П. Лекции о работе больших полушарий головного мозга. М.: Изд-во АМН СССР, 1952. 285 с.
- Симонов П.В. Мотивированный мозг. М.: Наука, 1987. 265 с.
- Хорн Г. Память, импринтинг и мозг. М.: Мир, 1988. 344 с.
- Шеперд Г. Нейробиология: В 2 т. М.: Мир, 1987. Т.1. 454 с; Т.2. 368 с.
Дополнительная
- Адам Д. Восприятие, сознание, память. М.: Мир, 1981. С. 5—126.
- Анохин П.К. Биология и нейрофизиология условного рефлекса. М.: Медицина, 1968. С. 289—349, 381—405.
- Воронин Л.Г. Физиология высшей нервной деятельности. М.: Высш. шк., 1979. С. 3—210.
- Дарвин Ч. Выражение эмоций у человека и животных. Соч. М.: Изд-во АМН СССР, 1953. Т. 5.
- Шульговский В.В. Физиология целенаправленного поведения млекопитающих. М.: Изд-во Моск. ун-та, 1993.
- Эделмен Дж., Маунткасл В. Разумный мозг. М.: Мир, 1981.
С. 5—126.
Программу составили
Е.Н.Соколов, доктор биологических наук;
Л.К.Хлудова, кандидат психологических наук
Факультет психологии Московского государственного университета имени М.В. Ломоносова
125009, Москва, ул. Моховая, д. 11, стр. 9. Схема проезда. Телефонный справочник.
Дизайн и поддержка сайта 1997-2020: Станислав Козловский
Исследуя процессы образования и торможения условных рефлексов, Павлов пришел к выводу о том, что в коре больших полушарий происходят как аналититические, так и синтетические процессы. Так, сам факт образования условного рефлекса отражает процессы синтеза, то есть объединения в коре двух или нескольких возбужденных очагов, между которыми устанавливается временная связь. Выработка дифференцировочного торможения лежит в основе анализа (различения) близкородственных раздражителей. Выработка условных рефлексов улучшает аналитические способности мозга. Так, если человеку на ладонь вытянутой руки положить сначала грузик весом 30 г, а затем - в 31 г, то человек не ощущает разницы в массе грузиков. Но если грузик массой в 30 г подкреплять слабым электрическим током, то есть сделать его сигналом условного оборонительного рефлекса, то человек очень четко анализирует различие между грузиками.
На первых этапах образования условных рефлексов животное отвечает сходными реакциями на сигналы, похожие на условный раздражитель. Так, если условный пищевой рефлекс выработан на ноту ДО, то сначала любая другая нота вызывает этот же рефлекс. Эта стадия, получившая название стадии генерализации условного рефлекса, отражает синтетические процессы в коре больших полушарий. Но, поскольку едой подкрепляется только нота ДО, нес другие неподкрепляемые ноты перестают вызывать условное слюноотделение. Эта стадия получила название стадии специализации условного рефлекса, в основе которой лежат аналитические процессы в коре, обеспечивающие распознавание звуков.
Павлов создал учение о синтез-анализаторах, суть которого в том, что сложные процессы анализа и синтеза в высшей нервной деятельности обеспечиваются совместной работой основных анализаторов: зрительного, слухового, обонятельного, кожного, двигательного, вестибулярного, вкусового, а также анализаторов, рецепторы которых заложены во внутренних органах.
Каждый анализатор состоит из трех частей:
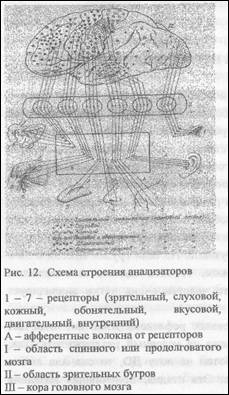
периферического, проводникового и мозгового отделов (рис.12). Периферический отдел анализатора представлен рецепторами, которые воспринимают внешние и внутренние сигналы. Рецепторами зрительного анализатора являются палочки и колбочки, расположенные в сетчатке глаза, рецепторами слухового анализатора -чувствительные клетки кортиева органа, расположенного в улитке, рецепторами кожного анализатора -многочисленные рецепторы кожи всей поверхности тела. Рецепторы двигательного анализатора расположены в мышцах и сухожилиях, составляя группу проприорецепторов. Вкусовые рецепторы покрывают поверхность языка и ротовой полости и раздражаются при пережевывании пищи. Рецепторы вестибулярного анализатора, отвечающего за правильное положение тела в пространстве, расположены в полукружных каналах и в преддверии внутреннего уха. Проводниковый отдел каждого анализатора представлен соответствующими чувствительными (афферентными, центростремительными) нервами: зрительными, слуховыми, обонятельными, вестибулярными. Часть центростремительных нервов не имеет специальных названий, входя в состав лицевого, языкоглоточного, подъязычного нервов, которые несут информацию
от вкусовых рецепторов. Информация от кожных рецепторов, проприорецепторов и интерорецепторов идет к головному мозгу по нервным волокнам мощных восходящих путей в составе задних и частично боковых канатиков спинного мозга. Характерной особенностью всех чувствительных нервов у позвоночных является их полное или частичное перекрещивание, в результате чего у млекопитающих информация о раздражений рецепторов левой половины туловища поступает в правое полушарие, а информация о раздражении рецепторов правой половины тела - в левое полушарие.
Скорость проведения возбуждения от рецепторов и органов чувств в головной мозг по чувствительным нервам составляет 100-120 м/сек и обеспечивается самыми быстродействующими миелиновыми нервными волокнами типа А. В результате столь быстрого проведения информации от рецепторов к Головному мозгу организм в состоянии быстро оценить характер и силу действующих стимулов и организовать адекватную ответную реакцию.
Организация ответных реакций организма на различные внешние и внутренние стимулы обеспечивается мозговым отделом анализатора. У низших позвоночных с неразвитыми полушариями мозга главную роль в организации реакций на зрительные, звуковые и вестибулярные раздражители играют средний мозг и мозжечок. У высших позвоночных, млекопитающих, вся информация о раздражении органов чувств и рецепторов поступает в кору больших полушарий, наиболее молодой в филогенетическом плане и наиболее сложный по строению отдел головного мозга. Поэтому мозговой отдел анализаторов у млекопитающих называется корковым отделом. Процесс перемещения всех чувствительных функций в одну плоскость - коры больших полушарий, получивший название кортикализации функций, привел к качественному скачку в аналитико-синтетической функции мозга. Действительно, кора больших полушарий представляет не что иное, как совокупность всех анализаторов, которые взаимодействуют между собой за счет процессов иррадиации возбуждения и образования временных связей, обеспечивая высший; корковый анализ и синтез внешних и внутренних раздражителей.
Аналитические процессы возможны и на уровне периферических отделов анализаторов, то есть на уровне рецепторов, органов чувств. Это так называемый периферический анализ. Уже на уровне сетчатки глаза за счет специализации в процессе эволюции ее рецепторов и разделения их на палочки и колбочки происходит первичный анализ зрительных сигналов. Палочки
Достаточно сложный периферический анализ происходит в сетчатке глаза лягушки. Здесь расположены светочувствительные клетки различных типов, одни из которых возбуждаются при смене освещенности, другие - при появлении быстро движущихся предметов (пролетающей бабочки, стрекозы), третьи - при шевелении очень мелких предметов, например кончика травинки. Информация от этих чувствительных клеток поступает в мозг лягушки по отдельным проводникам, что облегчает аналитико-синтетическую деятельность мозга лягушки.
Периферический анализ осуществляется и на уровне кожных рецепторов опять же в силу их специализации в ходе эволюции. Действительно, на поверхности кожи находятся высокочувствительные рецепторы: термо-, хемо-, барорецепторы, а также болевые рецепторы. Совместная работа всех этих групп рецепторов позволяет дифференцировать, то есть анализировать, различные свойства внешних контактных раздражителей. Слуховые рецепторы кортиева органа, расположенного в улитке, представлены десятками тысяч высокоспециализированных клеток, воспринимающих звуки разной частоты.
По мере эволюционного развития в жизни организма все большее значение стали приобретать высшие формы анализа и синтеза, осуществляемые в мозговых отделах анализаторов, у млекопитающих - в корковых отделах анализаторов.
Рассмотрим, как проявляются аналитико-синтетические процессы в высшей нервной деятельности млекопитающих при выработке сложных условных рефлексов, например, рефлексов на одновременный комплекс раздражителей. Напомним, как вырабатываются и проявляются такие рефлексы. Взятые в виде аккорда ноты ДО, МИ, СОЛЬ третьей октавы подкрепляются едой, в результате чего вырабатывается условный рефлекс на данный комплекс звуков. Однако, на первых этапах выработки каждая нота, взятая по отдельности, вызывает эту же реакцию, и лишь по мере укрепления условного рефлекса отдельные
раздражители теряют сигнальное значение, и условный рефлекс осуществляется лишь на комплекс данных звуков. Эти особенности при выработке условного рефлекса на комплексный раздражитель отражают сложные синтетические процессы в коре больших полушарий. На первых этапах выработки каждый из раздражителей связывается временной связью

с центром безусловного подкрепления, то есть с корковым представительством пищевого центра (рис. 13). Однако, поскольку эти частные связи не подкрепляются едой, то в конце концов они тормозятся. Центры, воспринимающие музыкальные звуки, объединяются в так называемый Функциональный комбинационный центр. Между этим единым функциональным центром и корковым представительством пищевого центра образуется временная связь, то есть вместо временных связей (а, б, в) устанавливается общая связь (г). Внешним выражением этих сложных процессов синтеза в коре больших полушарий и является тот факт, что сигнальное значение от отдельных раздражителей переходит к их комплексу. Подобные функциональные комбинационные центры образуются при выработке условных рефлексов на последовательные комплексы, на цепи раздражителей. При этом в качестве комплексов могут выступать раздражители разной модальности, например, свет, звук, кожное раздражение. В этих случаях функциональные комбинационные центры образуются между пространственно удаленными зонами коры больших полушарий - зрительной, слуховой, кожной чувствительности.
Сложные аналитико-синтетические процессы в коре больших полушарий обеспечивают системность в работе высших отделов мозга, что облегчает его
что можно было вместо девяти разных нот предъявить собаке первую ноту ДО и с привычными интервалами повторить ее 9 раз. В результате у собаки воспроизводится выработанный стереотип внешних ответов, то есть собаки будет реагировать на девятикратное повторение одной ноты как на чередование положительных и отрицательных условных сигналов. Следовательно, у собаки на внешний стереотип раздражителей выработался стереотип условнорефлекторных ответов, состоящий из чередования положительных (слюноотделение) и отрицательных (дифференцировочное торможение) реакций. Этот стереотип является достаточно прочным, поскольку

предъявление одного первого раздражителя девять раз не изменяет характерный стереотипный ответ, состоящий из положительных и отрицательных реакций. Более того, можно предъявить собаке только первый раздражитель - ноту ДО и наблюдать реализацию всей программы стереотипных ответов.
Каков же внутренний механизм динамического стереотипа? В его основе лежат сложные процессы синтеза между последовательно возбуждающимися центрами коры больших полушарий при стереотипном предъявлении условных раздражителей. В результате последовательно возбуждающиеся центры в слуховой зоне коры больших полушарий, ответственные за восприятие каждой из предъявляемых музыкальных нот, образуют цепочку за счет возникновения временных связей между этими центрами. В итоге, возбуждение первого центра при предъявлении первой ноты ДО3, вызывая свою ответную реакцию, вместе с тем служит сигналом для возбуждения второго центра, что приводит к реализации второго условнорефлекторного ответа и служит причиной возбуждения третьего центра и т.д. Таким образом, возбуждение первого центра служит сигналом последовательного возбуждения второго, третьего и последующих центров, результатом чего и является воспроизведение системы стереотипных условных ответов. Следовательно, причиной второй, третьей и последующих условных реакций может быть не условный раздражитель, а возбуждение предшествующих нервных центров. Такие условные рефлексы, укороченные с афферентного конца, получили название укороченных условных рефлексов 2 типа (Купалов). Большую роль в организации динамического стереотипа играют укороченные условные рефлексы 1 типа. Эти рефлексы связаны с мобилизующим действием обстановочных раздражителей, которые, не вызывая условных рефлексов, тем не менее настраивают определенные нервные центры на конкретную деятельность. В силу этого обстановочные раздражители могут играть роль переключателя условнорефлекторной деятельности. Так, мы знаем, что на один и тот же условный раздражитель, например, свет, в одной комнате можно выработать у животного условный слюноотделительный, в другой комнате - условный оборонительный рефлекс.
При выработке динамического стереотипа обстановочные раздражители также подготавливают, настраивают всю цепочку нервных центров на определенную деятельность, однако, вызываемое обстановкой возбуждение центров
внешне не проявляется, что позволяет говорить об укороченных с эфферентного конца условных рефлексах 1 типа.
Таким образом, в основе динамического стереотипа лежат сложные процессы объединения, синтеза возбужденных центров в коре с образованием временных связей между ними, что делает возможным реализацию динамического стереотипа через систему укороченных условных рефлексов 1 и 2 типа.
Если же слегка изменить порядок внешних раздражителей, например, поменять местами ноты ДО5 и ФА4 (см. протокол опыта), то это приведет к возникновению у животного отрицательных эмоций, что может закончиться срывом высшей нервной деятельности. Если все-таки выработать у собаки новую модификацию старого стереотипа, то животное опять успокаивается и реагирует системой ответов в соответствии с новым порядком условных раздражителей. Что будет, если вернуться к старому стереотипу, от которого животное с таким трудом отказалось? Оказалось, это опять сопровождается возникновением отрицательных эмоций. Все эти наблюдения свидетельствуют о том, что не только выработка динамического стереотипа, но и его переделка неизменно связаны с возникновением отрицательных эмоций у животных.
Явления динамического стереотипа сопровождают человека на протяжении всей его жизни. При этом для каждого возрастного периода характерны свои
стереотипы, то есть привычки бытового, социального плана. Так, у детей до 3 лет, находящихся в домашних условиях, вырабатываются детские стереотипы -реакции на постоянное общение с матерью к близкими, которые формируют у ребенка определенные навыки гигиены, питания, игры. Детские стереотипы отражают жизнь ребенка в соответствии с конкретным домашним укладом, где чередуются игры, прогулки, чтение детских книг и т.д. Поскольку образование ранних условных рефлексов идет по механизму импринтинга, детские стереотипы необыкновенно прочны, чем и объясняется присущий маленьким четям консерватизм и педантизм в поведении. Небольшой сбой во внешних условиях воспринимается ребенком болезненно, то есть сопровождается появлением отрицательных эмоций, которые недвусмысленно проявляются в форме плача, капризов, упрямства. Такое состояние наблюдается у малыша, если вместо матери другой человек начинает кормить его, читать ему книжку и т.д. Детские стереотипы, сменяясь взрослыми, тем не менее, никогда не разрушаются и могут всплыть при тяжелой болезни, в старости, когда человек становится раздражительным, капризным и эгоцентричным, то есть проявляет черты раннего детского возраста.
Первая серьезная ломка детского домашнего стереотипа происходит тогда, когда ребенка отдают в детский садик. Ребенка окружает все новое, к нему предъявляют новые требования, приспособление к которым, как правило, проходит у детей чрезвычайно болезненно. Старый стереотип должен уступить место новому, что приводит к возникновению сильных отрицательных эмоций. Нот почему психологи советуют родителям сначала приводить ребенка в детский садик только на несколько часов и постепенно увеличивать продолжительность его пребывания там. Тем самым снимается острота переживаний, то есть уменьшается психологическая нагрузка на ребенка, которая может привести к развитию неврозов, требующих уже специального лечения.
У обычных детей со здоровой психикой процесс перестройки, то есть выработки нового садикового стереотипа, заканчивается в среднем в течение одного месяца, и вот уже ребенок радостно идет в садик, где его ждут друзья, игры, прогулки, музыкальные занятия и прочие радости детской жизни. Сформировался новый жизненный стереотип на несколько лет, и жизнь по новому распорядку, новым социальным правилам сопровождается
положительными эмоциями, что проявляется в хорошем, ровном настроении ребенка, его физической и социальной активности, контактности.
Переход ребенка от садикового к школьному стереотипу тоже проходит очень болезненно, но устоявшийся стереотип привычной жизни и общения в школьном возрасте сопровождается положительными эмоциями, столь сильными, что подчас люди продолжают всю жизнь поддерживать дружбу со школьными друзьями.
Рассмотрение смены жизненных стереотипов у человека можно продолжить до наступления его старости, когда уход на пенсию становится чрезвычайно тяжелым для человека в психологическом плане, поскольку это связано с отказом от активной деятельности в привычном коллективе. Можно избежать тяжелых депрессивных состояний в пожилом возрасте, если заранее позаботится о выборе интересного дела - воспитания внуков, ухода за животными, цветами и т.д. Если же этого не сделать, то тяжелые психологические переживания могут привести к развитию депрессии и различных заболеваний.
Таким образом, в жизни человека все время происходит образование и смена стереотипов в соответствии с возрастными периодами. Отсюда название динамический стереотип, то есть стереотип, постоянно изменяющийся в течение долгой жизни, что обеспечивает адаптацию человека к новым требованиям его физической и социальной среды обитания.
Системность в работе коры больших полушарий, ярко проявляясь в динамическом стереотипе, имеет принципиальное значение для сохранения длительной работоспособности человека, его здоровья и долголетия. Павлов, отдавший науке 60 лет своей жизни, утверждал, что его высокая работоспособность поддерживалась за счет постоянного, неизменного в течение многих десятилетий распорядка дня, где разумно чередовались периоды интенсивной умственной деятельности и периоды активного отдыха. Жизнь по стереотипу легка, утверждал Павлов, поскольку каждая предыдущая деятельность является стимулом для последующей, создавая тем самым известный автоматизм в действиях и поступках, легкость переключения с одной формы деятельности на другую. Именно эта легкость, автоматизм при выполнении привычного набора разнообразных дел в течение дня и является основой тех положительных эмоций, которые неизменно сопутствуют устоявшемуся образу жизни, то есть устойчивому динамическому стереотипу.
Дата добавления: 2018-06-01 ; просмотров: 312 ;
Читайте также:
