
Биоэлектрический ток распространяющийся по нервным волокнам через специальные структуры
Электрические потенциалы, возникающие в тканях и отдельных клетках человека, животных и растений, важнейшие компоненты процессов возбуждения (См. Возбуждение) и торможения (См. Торможение). Исследование Б. п. имеет большое значение для понимания физико-химических и физиологических процессов в живых системах и применяется в клинике с диагностической целью (Электрокардиография, Электроэнцефалография, Электромиография и др.).
Различают следующие основные виды Б. п. нервных и мышечных клеток: потенциал покоя, потенциал действия, возбуждающие и тормозные постсинаптические потенциалы, генераторные потенциалы.
Потенциал покоя (ПП, мембранный потенциал покоя). У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует разность потенциалов (ПП) порядка 60—90мв, которая локализована на поверхностной мембране. Внутренняя сторона мембраны заряжена электроотрицательно по отношению к наружной (рис. 1). ПП обусловлен избирательной проницаемостью покоящейся мембраны для ионов К + (Ю. Бернштейн, 1902, 1912; А. Ходжкин и Б. Катц, 1947). Концентрация К + в протоплазме примерно в 50 раз выше, чем во внеклеточной жидкости, поэтому, диффундируя из клетки, ионы выносят на наружную сторону мембраны положительные заряды, при этом внутренняя сторона мембраны, практически не проницаемой для крупных органических анионов, приобретает отрицательный потенциал. Поскольку проницаемость мембраны в покое для Na + примерно в 100 раз ниже, чем для К + , диффузия натрия из внеклеточной жидкости (где он является основным катионом) в протоплазму мала и лишь незначительно снижает ПП, обусловленный ионами К + . В скелетных мышечных волокнах в возникновении потенциала покоя важную роль играют также ионы Cl - , диффундирующие внутрь клетки. Следствием ПП является ток покоя, регистрируемый между поврежденным и интактным участками нерва или мышцы при приложении отводящих электродов. Мембраны нервных и мышечных клеток (волокон) способны изменять ионную проницаемость в ответ на сдвиги мембранного потенциала. При увеличении ПП (гиперполяризация мембраны) проницаемость поверхностных клеточных мембран для Na + и К + падает, а при уменьшении ПП (деполяризация) она возрастает, причём скорость изменений проницаемости для Na + значительно превышает скорость увеличения проницаемости мембраны для К + .
Постсинаптические потенциалы (ПСП) возникают в участках мембраны нервных или мышечных клеток, непосредственно граничащих с синаптическими окончаниями. Они имеют амплитуду порядка нескольких мв и длительность 10—15 мсек. ПСП подразделяются на возбуждающие (ВПСП) и тормозные (ТПСП). ВПСП представляют собой местную деполяризацию постсинаптической мембраны, обусловленную действием соответствующего медиатора (например, ацетилхолина в нервно-мышечном соединении). При достижении ВПСП некоторого порогового (критического) значения в клетке возникает распространяющийся ПД (рис. 5, а, б). ТПСП выражается местной гиперполяризацией мембраны, обусловленной действием тормозного медиатора (рис. 5, в). В отличие от ПД, амплитуда ПСП постепенно увеличивается с увеличением количества выделившегося из нервного окончания медиатора. ВПСП и ТПСП суммируются друг с другом при одновременном или последовательном поступлении нервных импульсов к окончаниям, расположенным на мембране одной и той же клетки.
Генераторные потенциалы возникают в мембране чувствительных нервных окончаний — рецепторов. Они внешне сходны с ВПСП — их амплитуда порядка нескольких мв и зависит от силы приложенного к рецептору раздражения (рис. 6). Когда генераторный потенциал достигает порогового (критического) значения, в соседнем участке мембраны нервного волокна возникает распространяющийся ПД. Ионный механизм генераторных потенциалов ещё недостаточно изучен.
Наряду с перечисленными относительно быстро развивающимися Б. п., в нервных клетках, волокнах гладких мышц и некоторых растительных клетках регистрируются также очень медленные колебания мембранного потенциала неизвестной природы, причём на гребне волны деполяризации мембраны часто возникают разряды импульсов.
Все Б. п. могут быть зарегистрированы и точно измерены только с помощью внутриклеточных микроэлектродов, позволяющих отводить разности потенциалов между внутренней и наружной сторонами мембраны клетки. При отведении колебаний Б. п. от целых нервов, мышц или мозга с помощью поверхностных электродов регистрируется лишь суммарно потенциал множества синхронно или, чаще, асинхронно работающих клеток. Так, электромиограмма представляет собой результат сложения (интерференции) ПД множества скелетных мышечных волокон; электрокардиограмма — результирующая колебаний электрических потенциалов мышечных волокон различных отделов сердца; электроэнцефалограмма — результат суммации главным образом ВПСП и ТПСП множества клеток различных слоев коры больших полушарий. Регистрация таких интерференционных электрограмм, хотя и не позволяет анализировать колебания Б. п. отдельных клеток, имеет важное значение для суждения о состоянии исследуемого органа в целом. В клинической практике электромиограмму, электрокардиограмму и электроэнцефалограмму регистрируют с помощью электродов, расположенных на коже соответствующих частей тела. Оценка данных, полученных этими методами, основана на сопоставлении изменений характера соответствующей кривой с результатами клинических, физиологических и патологоанатомических исследований.
Лит.; Физиология человека, М., 1966; Гальвани Л. и Вольта А., Избр. работы о животном электричестве, М. — Л., 1937; Ходжкин А., Нервный импульс, пер. с англ., М., 1965; Экклс Дж., Физиология нервных клеток, пер. с англ., М., 1959; его же, Физиология синапсов, М., 1966; Катц Б., Нерв, мышца и синапс, пер. с англ., М., 1968; Ходоров Б. И., Проблема возбудимости, Л., 1969.
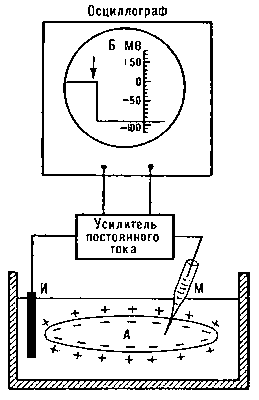
Рис. 1. Схема измерений мембранного потенциала покоя с помощью внутриклеточного стеклянного микроэлектрода (М). Второй электрод (И) помещен в омывающую клетку жидкость.
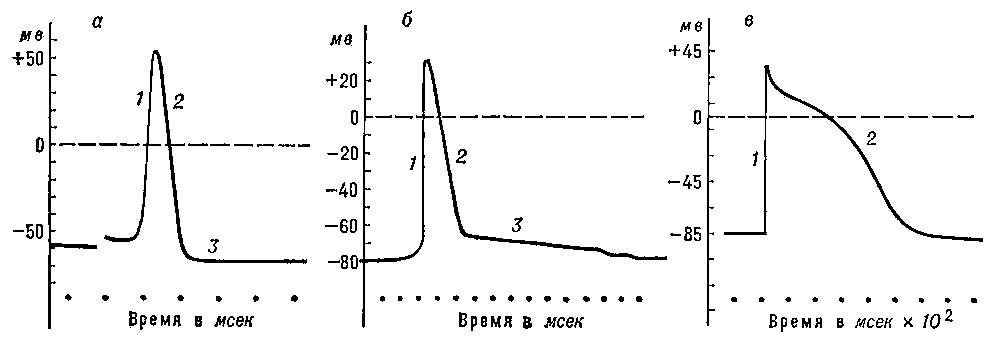
Рис. 2. Потенциалы действия, зарегистрированные с помощью внутриклеточных микроэлектродов: а — гигантского аксона кальмара; б — скелетного мышечного волокна; в — волокна мышцы сердца собаки; 1 — восходящая фаза ПД; 2 — нисходящая фаза; 3 — следовая гиперполяризация (а) и следовая деполяризация (б).
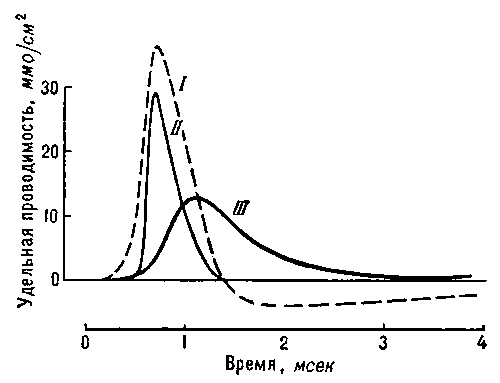
Рис. 3. Изменения натриевой и калиевой проводимости мембраны нервного волокна во время генерации потенциала действия (I). Изменения проводимости пропорциональны изменениям проницаемости для Na + (II) и К + (III).
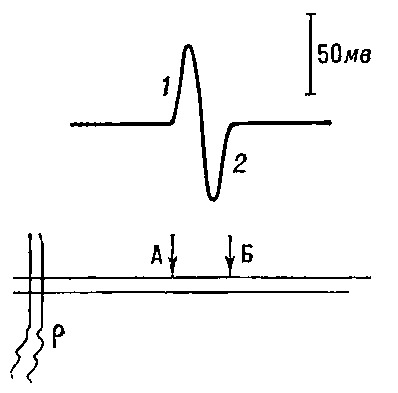
Рис. 4. Регистрация распространения потенциала действия вдоль нервного волокна. А, Б — внеклеточные электроды; р — раздражающие электроды. Вверху — отклонение луча осциллографа под влиянием волны возбуждения; 1 — волна возбуждения под электродом А; 2 — под электродом Б.
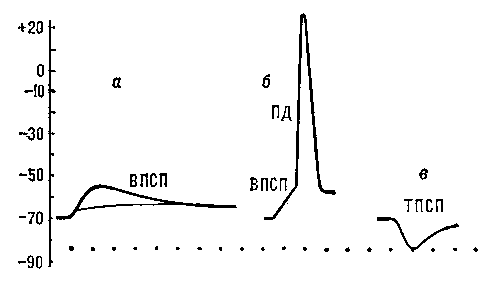
Рис. 5. Постсинаптические потенциалы: а — подпороговые ВПСП, возникающие в нервной клетке в ответ на раздражения соответствующих нервных волокон; б — ВПСП, достигший порогового значения, достаточного для возникновения ПД; в — ТПСП, вызванный раздражением тормозных нервных волокон.
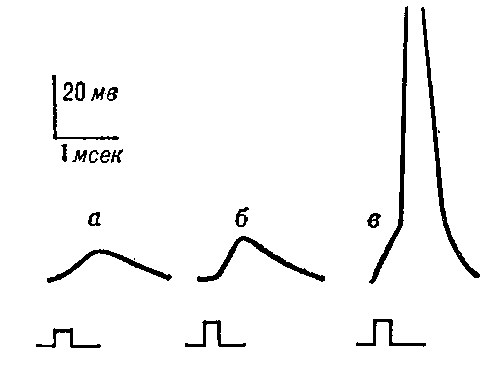
Рис. 6. Генераторные потенциалы: увеличение амплитуды при усилении раздражения рецептора (а — в). При достижении порогового значения (в) генераторный потенциал вызвал в чувствительном нервном волокне потенциал действия.
Возникновение и распространение возбуждения сопровождаются электрическими явлениями в тканях.
Рис. СХЕМА ОПЫТА ВТОРИЧНОГО СОКРАЩЕНИЯ (ОБЪЯСНЕНИЕ В ТЕКСТЕ).
Возражения, которые выдвигал Вольта, основывались на доказанном им факте, что при соединении двух разных металлов, в опыте Гальвани — меди и железа, возникает разность потенциалов, которая и вызывает сокращение мышцы.
Начался интересный научный спор между Гальвани и Вольта. По мере развертывания этого спора каждая из сторон прибегала к новым опытам для доказательства правоты своих взглядов. Решающим был опыт Гальвани, который он проводил без участия металлов. Этот опыт, получивший название второго опыта Гальвани, или сокращения без металлов, заключался в следующем: у лягушки отпрепаровывали седалищный нерв и набрасывали на обнаженные мышцы голени; при этом мышцы сокращались.
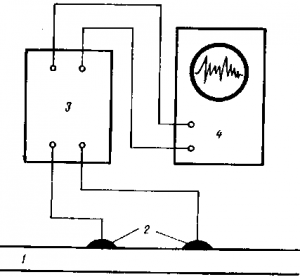
Рис. 2 Схема осциллографа для записи биопотенциалов, 1 — возбудимая ткань, 2 — электроды, 3- усилитель, 4- осцилограф
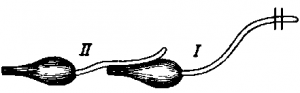
Особенно интересными были опыты, поставленные в середине прошлого века и получившие название вторичного тетануса, или вторичного сокращения. На мышцу одного нервно-мышечного препарата накладывали нерв другого нервно-мышечного препарата (рис. ). При раздражении индукционным током нерва первого нервно-мышечного препарата сокращалась и мышца второго препарата, нерв которого был наброшен на мышцу первого препарата. Это явление объяснялось тем, что при возбуждении в мышце возникали биоэлектрические явления, которые вызывали возбуждение второго нервно-мышечного препарата.
Исследования, проведенные уже во второй половине XIX века и в XX веке с использованием новейших физических приборов, внесли очень много в учение о биоэлектрических явлениях.
Очень ценные факты были получены крупнейшим русским физиологом Н Е. Введенским при помощи разработанной им методики телефонического выслушивания токов действия мышц и нервов. Выслушивание в телефонную трубку мышц и нервов при их возбуждении давало возможность улавливать особые звуки, связанные с возникновением токов действия и их ритмами.
Детальное изучение биоэлектрических явлений стало возможным благодаря тому, что был сконструирован и введен в физиологическую практику струнный гальванометр. Это настолько чувствительный прибор, что при его помощи удается обнаружить и зарегистрировать весьма малые электрические колебания в живых тканях.
Опишем схематически устройство гальванометра. Между двумя сильными электромагнитами протянута очень тонкая платиновая или кварцевая нить, покрытая золотом или серебром. Концы этой нити (струны) соединяются с электродами, которые прикладывают к нерву или мышце. Когда по струне проходит электрический ток, она отклоняется к одному из электромагнитов. Гальванометр устроен так, что при помощи специальных осветителей, струна освещается и ее движения фотографируются на фотографической бумаге. Колебания струны, сфотографированные на бумаге, дают возможность более детально изучить особенности биоэлектрических явлений. Гальванометр позволяет изучить весьма слабые токи. Струна гальванометра в новейших приборах может совершить до 1000 колебаний в секунду.
В настоящее время пользуются осциллографами разной конструкции, которые дают возможность еще более детально изучить биоэлектрические явления в живых тканях.
Наиболее чувствительным и не обладающим инертностью прибором является катодный осциллограф. Принцип устройства этого прибора заключается в том, что при помощи спе циального приспособления к так называемой катодной лампе пропускается тонкий пучок электронов. Этот пучок проходит между двумя металлическими пластинками, которые соединены с нервом или мышцей. Разности потенциалов, которые появляются в исследуемой живой ткани, передаются металлическим пластинкам, и катодный луч отклоняется к той пластинке, которая имеет положительный электрический заряд. Отклонение луча за писывается на фотографической пленке, которая непрерывно движется. Полученные таким образом осциллограммы дают возможность изучать малейшие колебания электрических зарядов в исследуемой ткани.
При записи биотоков, которые очень малы, между объектом и осциллографом включаются усилители (рис. 2), что дает возможность записать биопотенциалы в тысячные доли вольта и продолжительностью в тысячные доли секунды.
При записи биопотенциалов мозга — электроэнцефалограммы, электрокардиограммы и др., часто пользуются чернильнопишущим приспособлением, что создает возможность записать колебания биопотенциалов чернилами на бумаге.
Статья на тему Биоэлектрические явления в тканях
4.3. Свойства импульсов возбуждения в центральной нервной системе. Биоэлектрические явления
Главной причиной возникновения и распространения возбуждения является изменение электрического заряда на поверхности живой клетки, т. е. так называемые биоэлектрические явления.
По обе стороны поверхностной клеточной мембраны в состоянии покоя создается разность потенциалов, равная около -60-(-90) мВ, причем поверхность клетки заряжена электроположительно по отношению к цитоплазме. Эту разность потенциалов называют потенциалом покоя, или мембранным потенциалом. Величина мембранного потенциала для клеток различных тканей различна: чем выше функциональная специализация клетки, тем она больше. Например, для клеток нервной и мышечной тканей она составляет -80-(-90) мВ, для эпителиальной ткани -18-(-20) мВ.
Причиной возникновения биоэлектрических явлений служит избирательная проницаемость клеточной мембраны. Внутри клетки в цитоплазме ионов калия больше в 30–50 раз, чем вне клетки, ионов натрия меньше в 8-10 раз, в 50 раз меньше ионов хлора. В состоянии покоя клеточная мембрана более проницаема для ионов калия, чем для ионов натрия, и ионы калия выходят через поры в мембране наружу. Миграция положительно заряженных ионов калия из клетки сообщает наружной поверхности мембраны положительный заряд. Таким образом, поверхность клетки в покое несет положительный заряд, тогда как внутренняя сторона мембраны оказывается заряженной отрицательно за счет ионов хлора, аминокислот и других органических ионов, которые через мембрану практически не проникают.
Когда участок нервного или мышечного волокна подвергается действию раздражителя, в этом месте возникает возбуждение, проявляющееся в быстром колебании мембранного потенциала, называемого потенциалом действия.
Потенциал действия возникает из-за изменения ионной проницаемости мембраны. Происходит повышение проницаемости мембраны для катионов натрия. Ионы натрия поступают внутрь клетки под действием электростатических сил осмоса, тогда как в покое клеточная мембрана была малопроницаемой для этих ионов. При этом приток положительно заряженных ионов натрия из внешней среды клетки в цитоплазму значительно превышает поток ионов калия из клетки наружу. В результате происходит изменение мембранного потенциала (понижение мембранной разности потенциалов, а также возникновение разности потенциалов противоположного знака – фаза деполяризации). Внутренняя поверхность мембраны стала заряженной положительно, а наружная вследствие потери положительно заряженных ионов натрия – отрицательно, в этот момент регистрируется пик потенциала действия. Потенциал действия возникает в тот момент, когда деполяризация мембраны достигает критического (порогового) уровня.
Повышение проницаемости мембраны для ионов натрия продолжается короткое время. Затем в клетке возникают восстановительные процессы, приводящие к понижению проницаемости мембраны для ионов натрия и возрастанию для ионов калия. Поскольку ионы калия также заряжены положительно, то их выход из клетки восстанавливает исходные отношения потенциалов снаружи и внутри клетки (фаза реполяризации).
Изменение ионного состава внутри клетки и вне ее достигается несколькими путями: активным и пассивным трансмембранным переносом ионов. Пассивный транспорт обеспечивают присутствующие в мембране поры и селективные (избирательные) каналы для ионов (натрия, калия, хлора, кальция). Эти каналы обладают воротной системой и могут быть закрыты или открыты. Активный транспорт осуществляется по принципу натрийкалиевого насоса, который работает, потребляя энергию АТФ. Его основным компонентом является мембранная NA, КАТФаза.
Проведение возбуждения. Проведение возбуждения обусловлено тем, что потенциал действия, возникший в одной клетке (или в одном из ее участков), становится раздражителем, вызывающим возбуждение соседних участков.
По безмякотным нервным волокнам возбуждение распространяется медленно (от 1 до 30 м/с). Это связано с тем, что биоэлектрические процессы клеточной мембраны проходят на каждом участке волокна, по всей его длине.
Существует определенная зависимость между скоростью проведения возбуждения и диаметром нервного волокна: чем толще волокно, тем больше скорость проведения возбуждения.
Передача возбуждения в синапсах. Синапсом (от греч. synapsis – соединение) называется область контакта двух клеточных мембран, обеспечивающих переход возбуждения с нервных окончаний на возбуждаемые структуры. Возбуждение от одной нервной клетки к другой – процесс однонаправленный: импульс передается всегда с аксона одного нейрона на тело клетки и дендриты другого нейрона.
Аксоны большинства нейронов сильно ветвятся на конце и образуют многочисленные окончания на телах нервных клеток и их дендритах, а также на мышечных волокнах и на клетках желез. Количество синапсов на теле одного нейрона может достигать 100 и больше, а на дендритах одного нейрона – нескольких тысяч. Одно нервное волокно может образовать более 10 тыс. синапсов на многих нервных клетках.
Синапс имеет сложное строение. Он образован двумя мембранами – пресинаптической и постсинаптической, между которыми есть синаптическая щель. Пресинаптическая часть синапса находится на нервном окончании, постсинаптическая мембрана – на теле или дендритах нейрона, к которому передается нервный импульс. В пресинаптической области всегда наблюдаются большие скопления митохондрий.
Возбуждение через синапсы передается химическим путем с помощью особого вещества – посредника, или медиатора, находящегося в синаптических пузырьках, расположенных в пресинаптической терминали. В разных синапсах вырабатываются разные медиаторы. Чаще всего это ацетилхолин, адреналин или норадреналин.
Выделяют также электрические синапсы. Они отличаются узкой синаптической щелью и наличием поперечных каналов, пересекающих обе мембраны, т. е. между цитоплазмами обоих клеток есть прямая связь. Каналы образованы белковыми молекулами каждой из мембран, соединенных комплементарно. Схема передачи возбуждения в таком синапсе подобна схеме передачи потенциала действия в гомогенном нервном проводнике.
В химических синапсах механизм передачи импульса следующий. Приход нервного импульса в пресинаптическое окончание сопровождается синхронным выбросом в синаптическую щель медиатора из синаптических пузырьков, расположенных в непосредственной близости от нее. Обычно в пресинаптическое окончание приходит серия импульсов, частота их возрастает при увеличении силы раздражителя, приводя к увеличению выделения медиатора в синаптическую щель. Размеры синаптической щели очень малы, и медиатор, быстро достигая постсинаптической мембраны, взаимодействует с ее веществом. В результате этого взаимодействия структура постсинаптической мембраны временно изменяется, проницаемость ее для ионов натрия повышается, что приводит к перемещению ионов и, как следствие, возникновению возбуждающего постсинаптического потенциала. Когда этот потенциал достигает определенной величины, возникает распространяющееся возбуждение – потенциал действия. Через несколько миллисекунд медиатор разрушается специальными ферментами.
Выделяют также особые синапсы тормозного действия. Полагают, что в специализированных тормозящих нейронах, в нервных окончаниях аксонов вырабатывается особый медиатор, оказывающий тормозящее воздействие на последующий нейрон. В коре больших полушарий головного мозга таким медиатором считают гамма-аминомасляную кислоту. Структура и механизм работы синапсов тормозного действия аналогичны таковым у синапсов возбуждающего действия, только результатом их действия является гиперполяризация. Это ведет к возникновению тормозного постсинаптического потенциала, в результате чего наступает торможение.
На каждой нервной клетке расположено множество возбуждающих и тормозящих синапсов, что создает условия для различных ответов на прошедшие сигналы.
Данный текст является ознакомительным фрагментом.
Процессы жизнедеятельности неразрывно связаны с различными формами биоэлектрических явлений. В частности, биоэлектрические явления сопровождают возникновение возбуждения и его проведение по нервным волокнам, сокращение волокон скелетных, гладких мышц, миокарда, а также процессы всасывания в желудочно-кишечном тракте, восприятия, вкуса, запаха, деятельность анализаторов и т. д.
Причиной возникновения биоэлектрических явлений являются постоянно происходящие в процессе жизнедеятельности перераспределение ионов на мембранах и их трансмембранный транспорт [4,с.21]. На практике биоэлектрические явления регистрируют как разность электрически потенциалов между двумя точками живой ткани. Например, с помощью микро электродов можно измерить разность потенциалов между наружной и внутренней сторонами оболочки (мембраны) клетки.
Та разность потенциалов называется потенциалом покоя, или мембранным потенциалом. Наличие его обусловлено неравномерным распределением ионов (в первую очередь ионов натрия и калия) между внутренним содержимым клетки (ее цитоплазмой) и окружающей клетку средой.
Величина мембранного потенциала различна, причем для каждого типа клетки свойственна строго определенная величина потенциала покоя, которая отражает интенсивность обменных процессов, протекающих в них.
При возбуждении клетки возникает так называемый потенциал действия, который в отличие от потенциала покоя распространяется в виде волны возбуждения по поверхности клетки со скоростью нескольких десятков метров в секунду.
Существуют и другие виды потенциалов, например, потенциал повреждения.
Биоэлектрические явления исторически возникли как способ более совершенной связи между отдельными образованьями многоклеточного организма.
Биоэлектрические явления в деятельности ЦНС животных и человека
Существует два типа общения между клетками. Первый связан с химическим взаимодействием, при котором вещество, продуцируемое одной клеткой, достигает другой, вызывая в ней ответную реакцию.
Такой способ общения не обеспечивает быстрой передачи информации. Более совершенный способ передачи информации и сигнализации осуществляется при помощи нервных импульсов, одним из компонентов которых являются биоэлектрические явления.
Особенно отчетливо он выражен в деятельности центральной нервной системы животных и человека. Живой организм является не только генератором биопотенциалов, но и проводником электрического тока, причем изменение степени электропроводности живых тканей в зависимости от их жизнедеятельности может служить показателем жизнеспособности клеток или тканей [8, с. 83].
К наиболее важным функциям, присущим всем живым существам, можно отнести следующие:
- самовоспроизведение: запись, хранение и использование генетической информации;
- энергообеспечение: получение энергии за счет внешних энергетических ресурсов;
- превращение веществ: использование химических соединений окружающей среды для образования компонентов своего тела;
Раздражимость: способность принимать и обрабатывать сигналы, поступающие из внешней и внутренней сред организма, и адекватно отвечать на эти сигналы.
Биоэнергетика - это отрасль функциональной биологии, изучающая превращение энергии внешних ресурсов в биологически полезную работу.
Липидный компонент мембран более или менее стандартен: это фосфолипиды, реже глико - и сульфолипиды.
Белок - это тот компонент, который определяет специфику мембраны.
Важнейшей функцией многих мембран является превращение одной формы энергии в другую. Эта функция осуществляется особыми белками, встроенными в особого типа мембрану, носящую название энергопреобразующей. Такая мембрана непроницаема для большинства веществ, которые имеются в растворах по обе стороны.
Любая мембрана, выполняющая энергетическую функцию, способна к превращению химической энергии или энергии света в электрическую энергию, а именно в трансмембранную разность электрических потенциалов или в энергию разности концентраций веществ, содержащихся в разделенных мембраной растворах.
Некоторые энергообразующие мембраны могут превращать электрическую энергию в механическую. Определенные ткани теплокровных животных и человека превращают накопленную энергию в тепло в целях терморегуляции, что оказывается биологически оправданным при понижении окружающей температуры.
Таким образом, человеческий организм является генератором биопотенциалов, проводником электрического тока.
Биоэлектрические явления особенно ярко выражены в деятельности центральной нервной системы.
Любой агент, повышающий натриевую проницаемость мембраны, является раздражителем возбудимой ткани. Раздражителями нервных и мышечных волокон могут быть: электрический ток, механические воздействия (щипок, удар, разрез), резкое охлаждение или согревание, различные кислоты, щелочи, концентрированные растворы солей и т. д.
Среди всех указанных раздражителей электрический ток занимает особое место, так как, во-первых, он может быть легко и точно дозирован по силе, длительности и крутизне нарастания, а во-вторых, он не повреждает живую ткань и его действие быстро и полностью обратимо при тех его силах, которые достаточны для вызова возбуждения.
Изучение действия электрического раздражения на возбудимые ткани представляет большой интерес для физиологии, потому что проведение возбуждения в нервах и мышцах осуществляется с помощью локальных электрических токов, возникающих между возбужденным и покоящимся участком ткани.
В лабораторных условиях и при проведении некоторых клинических исследований для раздражения нервов и мышц применяют электрические стимулы различной формы: прямоугольной, синусоидальной, линейно и экспоненциально нарастающей, индукционные удары, конденсаторные разряды и т. п.
Механизм раздражающего действия тока при всех видах стимулов в принципе одинаков, однако в наиболее отчетливой форме он выявляется при использовании постоянного тока прямоугольной формы.
Для того чтобы раздражитель вызвал возбуждение, он должен иметь достаточную силу, длительность и крутизну нарастания. Та наименьшая сила раздражителя, которая необходима для возникновения потенциала действия в возбудимой ткани, называется порогом раздражения. Стимулы, сила которых ниже пороговой величины, называются подпороговыми, а более сильные, чем пороговые,- сверхпороговыми.
При использовании в качестве раздражителя электрического тока порог выражается в единицах силы тока или напряжения. Абсолютная величина порога зависит от свойств и физиологического состояния ткани, а также от способа нанесения раздражения.
Существует два способа подведения электрического тока к ткани: внеклеточный и внутриклеточный. Первый состоит в том, что оба электрода располагают на поверхности раздражаемой ткани. Ток входит в ткань в области анода и выходит в области катода.
Недостаток этого метода заключается в значительном ветвлении тока: только часть его проходит через мембраны клеток, часть же ответвляется в межклеточные щели. Вследствие этого при раздражении приходится применять значительно большую силу тока, чем это в действительности необходимо для возникновения возбуждения.
Более точным является второй способ раздражения посредством внутриклеточного электрода. Микроэлектрод с диаметром кончика около 0,5 мК вводят в клетку, второй - обычный электрод - прикладывают к поверхности ткани. В этом случае весь приложенный ток проходит через мембрану клетки, что позволяет точно определить величину порога раздражения: у различных клеток он варьирует в пределах 10
9 а. Внутриклеточное раздражение обычно сочетают с регистрацией потенциалов через другой, внутриклеточный электрод.
Минимальное время, в течение которого электрический ток должен действовать на ткань, чтобы вызвать распространяющееся возбуждение, находится в обратной зависимости от напряжения и силы тока.
Если по оси абсцисс отложить минимально необходимое время действия электрического стимула (например, толчка постоянного тока) в миллисекундах, а по оси ординат -- напряжение или силу тока, то мы получим кривую силы -- времени. Эта кривая была подробно изучена в опытах на различных нервах и мышцах Л. Гоорвегом, Г. Вейссом, Л. Лапиком, а в недавнее время Д. Н. Насоновым с сотрудниками.
Из рассмотрения этой кривой прежде всего следует, что ток ниже некоторой минимальной силы или напряжения не вызывает возбуждения, как бы длительно он не действовал. Минимальная сила тока (или напряжение), способная вызвать возбуждение, названа Л. Лапиком реобазой.
Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно.
При очень коротких стимулах кривая силы - времени становится параллельной оси ординат. Это означает, что при таких кратковременных раздражениях возбуждение возникает, как бы ни была велика сила раздражения. Кривая силы - времени имеет форму равносторонней гиперболы.
Определение полезного времени практически трудно, так как величина реобазы претерпевает непрерывно небольшие колебания, отражающие колебания функционального состояния мембраны в покое. По этой причине Л. Лапик (1909) предложил измерять другую, условную, величину, названную им хронаксией. Хронаксия - это наименьшее время, в течение которого электрический ток, равный удвоенной реобазе (ОО), должен действовать на ткань, чтобы вызвать возбуждение. Полезное время и хронаксия характеризуют скорость возникновения возбуждения при действии раздражителя.
Опыты показали, что кривые силы - времени у самых разнообразных тканей, например нервов и мышц человека и теплокровных животных, желудка лягушки, ноги улитки и др., имеют одну и ту же форму.
Различия между ними лишь количественные: в нервных и мышечных волокнах позвоночных животных хронаксия измеряется тысячными и десятитысячными долями секунды, а в так называемых медленных тканях, например в мышечных волокнах ноги улитки или желудка лягушки,- в сотых долях секунды.
Эти факты привели исследователей к выводу, что возбудимые ткани отличаются друг от друга временной константой.
Определение хронаксии -- хронаксиметрия -- получило распространение не только в эксперименте, но и в клинической практике (А. Бургиньон, Ю. М. Уфлянд и др.). В частности, путем измерения хронаксии мышцы невропатолог может установить наличие повреждения волокон двигательного нерва. Дело в том, что при приложении электрического стимула к мышце ток проходит и через находящиеся в ней нервные волокна и их окончания.
Порог раздражения и хронаксия нервных волокон ниже, чем мышечных волокон. Поэтому при раздражении мышцы возбуждение прежде возникает в нервных волокнах и от них уже передается мышечным волокнам. Из этого следует, что при определении хронаксии нормальной мышцы человека фактически измеряется хронаксия иннервирующих ее нервных волокон. Если же нерв поврежден или произошла гибель иннервирующих мышцу клеток в спинном мозгу, то нервные волокна перерождаются, и тогда приложенный к мышце стимул выявляет хронаксию мышечных волокон, которая имеет большую продолжительность.
Величина порога раздражения нерва или мышцы зависит не только от длительности действия стимула, но и от крутизны нарастания его силы. Порог раздражения имеет наименьшую величину при толчках тока прямоугольной формы, характеризующихся максимально быстрым нарастанием силы.
Если же вместо толчков прямоугольной формы применять линейно или экспоненциально нарастающие стимулы, то пороги оказываю увеличенными и тем в большей мере, чем медленнее нарастает сила тока.
При уменьшении крутизны нарастания тока ниже некоторой минимальной величины потенциал действия вообще не возникает, до какой бы конечной силы не увеличивался ток. Обусловлено это тем, что за время нарастания силы раздражителя в ткани успевают развиться активные изменения, повышающие порог и препятствующие возникновению возбуждения.
Такое явление приспособления возбудимой ткани к медленно нарастающему раздражителю получило название аккомодации. Чем выше скорость аккомодации, тем более круто должен нарастать стимул, чтобы не утратить своего раздражающего действия.
Аккомодация развивается не только при раздражении возбудимых тканей электрическим током, но также и при применении механических, термических и прочих раздражителей.
Показателем скорости аккомодации является та наименьшая крутизна нарастания тока, при которой раздражающий стимул еще сохраняет способность вызывать потенциал действия. Эту минимальную крутизну нарастания тока называют минимальным градиентом, или критическим наклоном. Его выражают или в абсолютных величинах -- мА /сек, или в относительных единицах - реобаза/сек. При этом реобазу измеряют прямоугольным током, а затем рассчитывают, на сколько реобаз в секунду должен нарастать ток, чтобы он не утратил раздражающего действия.
Скорость аккомодации различных возбудимых образований широко варьирует. Наиболее велика скорость аккомодации двигательных нервных волокон теплокровных животных. Чувствительные волокна характеризуются меньшей скоростью аккомодации. Очень мала скорость аккомодации волокон сердечной мышцы, гладких мышц кишечника, мочеточников, желудка, т. е. всех образований, которые склонны к автоматической активности.
Постоянный электрический ток обладает полярным действием на возбудимую ткань. Оно выражается в том, что в момент замыкания цепи постоянного тока возбуждение в нерве или мышце всегда возникает только под катодом, а в момент размыкания только под анодом.
Э. Пфлюгер, открывший эти явления, доказал их путемследующего опыта: он умерщвлял участок нерва под одним из электродов, а второй электрод устанавливал на неповрежденный участок. Если с неповрежденным участком соприкасался катод, то возбуждение возникало в момент замыкания тока; если же катод устанавливали на поврежденном участке, а анод на неповрежденном, то возбуждение возникало только при размыкании тока. Порог раздражения при размыкании, когда возбуждение возникает под анодом, значительно выше, чем при замыкании, когда возбуждение возникает под катодом.
О возникновении возбуждения Пфлюгер судил косвенно по сокращению мышцы, иннервируемой раздражаемым нервом. В дальнейшем эти явления, обобщенные в форме полярного закона раздражения, были подтверждены и прямым способом - путем регистрации потенциалов действия непосредственно в участке приложения к ткани полюсов постоянного тока.
Для изучения механизма полярного действия электрического тока в настоящее время производят раздражение нервных и мышечных волокон и отведение от них электрических потенциалов с помощью внутриклеточных микроэлектродов. Установлено, что потенциал действия возникает только в том случае, если катод соприкасается с наружной поверхностью мембраны, а анод находится внутри клетки. При обратном расположении полюсов, т. о. наружном аноде и внутреннем катоде, возбуждение при замыкании тока не возникает, как бы он силен ни был.
Читайте также:
