
Биохимия возникновения и проведения нервного импульса
Механизм действия медиаторов.
Свойства сигнальных веществ нейромедиаторов, нейромодуляторов
· хранятся в везикулах;
· при поступлении нервного импульса выделяются в синаптическую щель,
· избирательно связываются со специфическим рецептором на постсинаптической мембране другого нейрона или мышечной клетки,
· индуцируют или ингибируют потенциал действия, контролируя возбуждение соседних нейронов и стимулируя клетки к выполнению специфических функций.
Синапсы образованы мембранами двух контактирующих клеток (пресинаптической и постсинаптической), которые разделены узкой синаптической щелью. Медиатор (или модулятор) выделяется в синаптическую щель за счет экзоцитоза, диффундирует к рецепторам постсинаптической мембраны, связывается с ними и передаёт сигнал соседней клетке.
Белки-рецепторы:
1) лиганд-активируемые ионные каналы – никотиновые холинэргические рецепторы (мышечные и нейрональные), ГАМК-рецепторы и глициновый рецептор. Вещество, действующее на ионотропные рецепторы, выступает медиатором. Большинство нейромедиаторов стимулирует открывание ионных каналов, и лишь немногие — закрывание.
2) чаще встречаются мембранные белки, которые управляют ионными каналами опосредованно с участием G-белков. Вещество, действущее на метаботропные рецепторы, выступает модулятором.
Клетки сообщаются друг с другом путём передачи сигналов, которые регистрируются в виде электрических процессов. Характер изменения мембранного потенциала постсинаптической клетки зависит от типа канала.
1)Изменение мембранного потенциала от -60 до +30 мВ за счёт открывания Nа + -каналов ведет к возникновению постсинаптического потенциала действия.
2)Изменение мембранного потенциала с -60 мВ до -90 мВ за счёт открывания Cl – -каналов ингибирует потенциал действия (гиперполяризация), в результате возбуждение не передается (тормозной синапс).
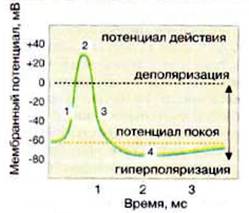
Рис. 6.Мембранный потенциал постсинаптической клетки
В состоянии покоя внутренняя сторона мембраны нейрона заряжена отрицательно. Это потенциал покоя, величина его достигает 60-75 мВ. Он обеспечивается в основном работой натриевого насоса (Na + ,К + -АТФазы). За счет энергии гидролиза каждой молекулы АТФ три иона Na + выкачиваются из клетки во внеклеточное пространство, а два иона К + перемещаются в противоположном направлении.
Разность потенциалов возникает в основном за счет следующих явлений. Несмотря на то что мембрана достаточно проницаема для ионов К + , их концентрация внутри аксона в 20-50 раз выше, чем во внешней среде. Главные внутриклеточные анионы (белки и нуклеиновые кислоты) не могут выходить наружу, а ионы С1 – , которых много во внешней среде, проходят через мембрану очень медленно. Проницаемость для ионов Na + составляет лишь 1/20 по сравнению с проницаемостью для ионов К + . При таких условиях трансмембранный потенциал определяется распределением ионов К + . Этот потенциал существует благодаря тому, что ионы К + стремятся покинуть клетку, чтобы уравнять внешнюю и внутреннюю концентрации. Однако в клетке при этом остается избыток анионов, что создает отрицательный электрический заряд, ограничивающий дальнейшее выравнивание концентраций ионов К + . Ионы С1 – ведут себя наоборот: они должны остаться снаружи, чтобы сбалансировать электрический заряд плохо проникающего Na + , но в то же время стремятся проникнуть в клетку по градиенту концентрации.
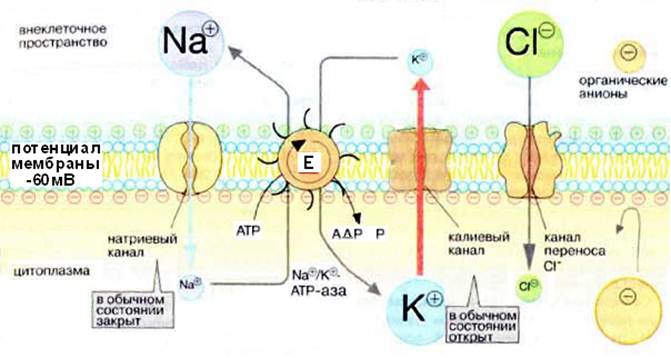
Рис. 6.Механизмы поддержания мембранного потенциала покоя
в постсинаптической клетке
За один импульс через мембрану проходит небольшое количество ионов Na + и К + , и концентрационные градиенты обоих ионов сохраняются: в клетке выше уровень К + , а вне клетки выше уровень Na + . Поэтому по мере получения клеткой новых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно.
Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращение мембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку.
Синапс - это функциональный контакт специализированных участков плазматических мембран двух возбудимых клеток. Синапс состоит из пресинаптической мембраны, синаптической щели и постинаптической мембраны. Мембраны клеток в месте контакта имеют утолщения в виде бляшек - нервных окончаний. Нервный импульс, достигший нервного окончания, не в состоянии преодолеть возникшее перед ним препятствие - синаптическую щель. После этого электрический сигнал преобразуется в химический. Пресинаптическая мембрана содержит специальные канальные белки, подобные белкам, формирующим натриевый канал в мембране аксона. Они тоже реагируют на мембранный потенциал, изменяя свою конформацию и формируют канал. В результате ионы Са2+ проходят через пресинаптическую мембрану по градиенту концентраций в нервное окончание. Градиент концентраций Са2+ создается работой Са2+-зависимой.
АТФазы - кальциевым насосом. Повышение концентрации Са2+ внутри нервного окончания вызывает слияние 200-300 имеющихся там везикул, заполненных ацетилхолином, с плазматической мембраной. Далее ацетилхолин секретируется в синаптическую щель путем экзоцитоза, и присоединяется к рецепторным белкам, расположенным на поверхности постсинаптической мембраны.
Ацетилхолиновый рецептор представляет собой трансмембранный олигомерный гликопротеиновый комплекс, состоящий из 6 субъединиц: 2-, 2-бета, 1-гамма и 1-дельта. Плотность расположения белков-рецепторов в постсинаптической мембране очень велика - около 20000 молекул на 1 мкм2. Пространственная структура рецептора строго сооответствует конформации медиатора. При взаимодействии с ацетилхолином белок-рецептор так изменяет свою конформацию, что внутри него формируется натриевый канал. Катионная селективость канала обеспечивается тем, что ворота канала сформированы отрицательно заряженными аминокислотами. Таким образом повышается проницаемость постсинаптической мембраны для натрия и возникает новый импульс (или сокращение мышечного волокна). Деполяризация постсинаптической мембраны выеывает диссоциацию комплекса "ацетилхолин-белок-рецептор" и ацетилхолин освобождается в синаптическую щель. Как только ацетилхолин оказывается в синаптической щели, он за 40 мкс подвергается быстрому гидролизу под действием фермента ацетилхолинэстеразы.
Во время гидролиза ацетилхолина образуется промежуточный фермент-субстратный комплекс, в котором ацетилхолин связан с активным центром фермента через серин.
Необратимое ингибирование холинэстеразы вызывает смерть. Ингибиторами холинэстеразы являются фосфорорганические соединения (хлорофос, дихлофос, табун, зарин, зоман, бинарные яды). Эти вещества связываются ковалентно с серином в активном центре фермента. Некоторые из них синтезированы в качестве инсектицидов, а некоторые - в качестве боевых отравляющих веществ (нервно-паралитические яды). Смерть наступает в результате остановки дыхания. Обратимые ингибиторы холинэстеразы используются как лечебные препараты. Например, при лечении глаукомы и атонии кишечника.
Катехоламины: норадреналин и дофамин. Адренэргические синапсы встречаются в постганглионарных волокнах, в волокнах симпатической нервной системы, в различных отделах головного мозга. Катехоламины в нервной ткани синтезируются по общему механизму из тирозина. Ключевой фермент синтеза - тирозингидроксилаза, ингибируемая конечными продуктами.
Катехоламины, как и ацетилхолин, накапливаются в синаптических пузырьках и тоже выделяется в синаптическую щель при поступлении нервного импульса. Но регуляция в адренэргическом рецепторе происходит иначе. В пресинаптической мембране здесь имеется специальный регуляторный белок - -ахромогранин (Мм = 77 кДа), который в ответ на повышение концентрации медиатора в синаптической щели связывает уже выделившийся медиатор и прекращает его дальнейший экзоцитоз. Фермента, разрушающего медиатор, в адренэргических синапсах нет. После передачи импульса молекулы медиатора перекачивается специальной транспортной системой путем активного транспорта с участием АТФ обратно через пресинаптическую мембрану и включается вновь в везикулы. В пресинаптическом нервном окончании излишек медиатора может быть инактивирован моноаминоксидазой, а также катехоламин-О-метилтрансферазой путем метилирования по оксигруппе. Кокаин тормозит активный транспорт катехоламинов.
Связывание медиатора с постсинаптическим рецептором почти мгновенно вызывает повышение концентрации ц-АМФ, что приводит к быстрому фосфорилированию белков постсинаптической мембраны. В результате изменяется генерация нервных импульсов постсинаптической мембраной (тормозится). В некторых случаях непосредственной причиной этого является повышение проницаемости постсинаптической мембраны для калия, либо снижением проводимости для натрия (эти события приводят к гиперполяризации).
ГАМК - тормозной медиатор. Повышает проницаемость постсинаптических мембран для ионов калия. Это ведет к изменению мембранного потенциала.
Глицин - тормозной медиатор, по вызываемым эффектам подобен гамк.
Образование нейромедиаторов – ацетилхолина, адреналина, дофамина, серотонина.
Информация, благодаря которой нейроны устанавливают только определенные связи с определенными нейронами, кодируется в структуре полисахаридных веточек мембранных гликопротеинов. Образование таких связей, не заложенных в период эмбрионального развития, является результатом опыта индивидуального организма и составляет материальную основу для хранения информации, определяющей особенности поведения данного организма.
Нейромедиаторы - это вещества, которые характеризуются следующими признаками. Накапливаются в пресинаптической структуре в достаточной концентрации. Освобождаются при передаче импульса. Вызывают после связывания с постсинаптической мембраной изменение скорости метаболических процессов и возникновение электрического импульса. Имеют систему для инактивации или транпортную систему для удаления из синапса, обладающие к ним высоким сродством. Таким образом, нейромедиаторы играют важную роль в функционировании нервной ткани, обеспечивая синаптическую передачу нервного импульса. Их синтез происходит в теле нейронов, а накопление - в особых везикулах, которые постепенно перемещаются с участием систем нейрофиламентов и нейротрубочек к кончикам аксонов.
Химическая классификация нейромедиаторов: Аминокислоты (и их производные) - К ним относят таурин, норадреналин, ДОФАминГАМК, глицин, ацетилхолин, гомоцистеин и некоторые другие (адреналин, серотонин, гистамин, серотонин).
Таурин - образуется из аминокислоты цистеина. Сначала происходит окисление серы в SH-группе до остатка серной кислоты (процесс идет в несколько стадий), а затем происходит декарбоксилирование. Таурин - это необычная кислота, в которой нет карбоксильной группы, а имеется остаток серной кислоты. Таурин принимает участие в проведении нервного импульса в процессе зрительного восприятия.
Ацетилхолин - Для синтеза холина требуются аминокислоты серин, метионин. Этаноламин может быть использован и в готовом виде. Но, как правило, из крови в нервную ткань пступает уже готовый холин. Второй же предшественник этого нейромедиатора - Ацетил-КоА, синтезируется в нервных окончаниях. Продукт этой реакции ацетилхолин участвует в синаптической передаче нервного импульса. Он накапливается в синаптических пузырьках, образуя комплексы с отрицательно заряженным белком везикулином. Передача возбуждения с одной клетки на другую осуществляется с помощью специального синаптического механизма.
Норадреналин - медиатор в постганглионарных волокнах симпатической и в различных отделах ЦНС.
Дофамин- медиатор проводящих путей, тела нейронов которого расположены в отделе мозга, который отвечает за контроль произвольных движений. Поэтому при нарушении дофаминэргической передачи возникает заболевание паркинсонизм.
Последнее изменение этой страницы: 2016-08-16; Нарушение авторского права страницы
Химическими передатчиками являются нейромедиаторы:
В передаче нервного возбуждения участвуют:
1. Натриевый насос (АТФ -аза)
2. Натриевые каналы
3. Калиевые каналы.
Этапы передачи нервного возбуждения.
1. Нервный импульс
2. Освобождение медиатора
3. Диффузия медиатора через синаптическую щель
4. Присоединение медиатора к рецептору
5. Изменение конфигурации рецептора
6. Открытие калий -натриевых каналов
7. Инактивация медиатора
8. Удаление натрия из клетки с помощью АТФ-азы.
Механизм, ну, чистая физиология.
Передача возбуждения в химическом синапсе - это сложный физиологический процесс, протекающий в несколько стадий. Он включает синтез и секрецию медиатора; взаимодействие медиатора с рецепторами постсинаптической мембраны; инактивирование медиатора. Каждый из этих этапов детально охарактеризован, и найдены препараты, которые избирательно усиливают или блокируют конкретный этап. В целом синапс осуществляет последовательную трансформацию электрического сигнала, поступающего по нервному волокну, в энергию химических превращений на уровне синаптической щели и постсинаптической мембраны, которая затем снова трансформируется в энергию распространяющегося возбуждения в эффекторной клетке. Исследования позволили проникнуть в механизм действия психотропных лекарственных средств, а также выявить связь некоторых нервных и психических болезней со специфическими нарушениями синаптических механизмов.
168. Медиаторы: ацетилхолин, катехоламины, серотонин, гамма-аминомаслянная кислота, глутаминовая кислота, глицин, гистамин.
Ацетилхолин. Для синтеза холина требуются аминокислоты серин, метионин. Этаноламин может быть использован и в готовом виде. Но, как правило, из крови в нервную ткань пступает уже готовый холин. Второй же предшественник этого нейромедиатора - Ацетил-КоА, синтезируется в нервных окончаниях. Продукт этой реакции ацетилхолин участвует в синаптической передаче нервного импульса. Он накапливается в синаптических пузырьках, образуя комплексы с отрицательно заряженным белком везикулином. Передача возбуждения с одной клетки на другую осуществляется с помощью специального синаптического механизма.
Дофамин- медиатор проводящих путей, тела нейронов которого расположены в отделе мозга, который отвечает за контроль произвольных движений. Поэтому при нарушении дофаминэргической передачи возникает заболевание паркинсонизм.
Катехоламины, как и ацетилхолин, накапливаются в синаптических пузырьках и тоже выделяется в синаптическую щель при поступлении нервного импульса. Но регуляция в адренэргическом рецепторе происходит иначе. В пресинаптической мембране здесь имеется специальный регуляторный белок - -ахромогранин (Мм = 77 кДа), который в ответ на повышение концентрации медиатора в синаптической щели связывает уже выделившийся медиатор и прекращает его дальнейший экзоцитоз. Фермента, разрушающего медиатор, в адренэргических синапсах нет. После передачи импульса молекулы медиатора перекачивается специальной транспортной системой путем активного транспорта с участием АТФ обратно через пресинаптическую мембрану и включается вновь в везикулы.
ГАМК - тормозной медиатор. Повышает проницаемость постсинаптических мембран для ионов калия. Это ведет к изменению мембранного потенциала. Глицин - тормозной медиатор, по вызываемым эффектам подобен гамк
Глутамат — наиболее распространенный возбуждающий нейротрансмиттер в нервной системе позвоночных. В химических синапсах глутамат запасается в пресинаптических пузырьках (везикулах). Нервный импульс запускает высвобождение глутамата из пресинаптического нейрона. На постсинаптическом нейроне глутамат связывается с постсинаптическими рецепторами, такими, как, например, NMDA-рецепторы, и активирует их. Благодаря участию последних в синаптической пластичности глутамат вовлечен в такие когнитивные функции, как обучение и память.
Гистамин. Некоторые количества гистамина содержатся в ЦНС, где, как предполагают, он играет роль нейромедиатора (или нейромодулятора). Не исключено, что седативное действие некоторых липофильных антагонистов гистамина (проникающих через гемато-энцефалический барьер противогистаминных препаратов, например, димедрола) связано с их блокирующим влиянием на центральные гистаминовые рецепторы. Подавление высвобождения нейромедиаторов (ГАМК, ацетилхолина, серотонина, норадреналина).
Серотонин играет роль нейромедиатора в ЦНС. Серотонинергические нейроны группируются в стволе мозга: в варолиевом мосту и ядрах шва. От моста идут нисходящие проекции в спинной мозг, нейроны ядер шва дают восходящие проекции к мозжечку, лимбической системе, базальным ганглиям, коре. При этом нейроны дорсального и медиального ядер шва дают аксоны, различающиеся морфологически, электрофизиологически, мишенями иннервации и чувствительностью к некоторым нейротоксичным агентам, например, метамфетамину.
Рассмотрим химические основы возникновения и поддержания биоэлектрических потенциалов (потенциала покоя и потенциала действия). Большинство исследователей придерживаются мнения, что явления электрической поляризации клетки обусловлены неравномерным распределением ионов К + и Na + по обе стороны клеточной мембраны. Мембрана обладает избирательной проницаемостью: большей для ионов К + и значительно меньшей для ионов Na + . Кроме того, в нервных клетках существует механизм, который поддерживает внутриклеточное содержание натрия на низком уровне вопреки градиенту концентрации. Этот механизм получил название натриевого насоса.
При определенных условиях резко повышается проницаемость мембраны для ионов Na + .
В состоянии покоя внутренняя сторона клеточной мембраны заряжена электроотрицательно по отношению к наружной поверхности. Объясняется это тем, что количество ионов Na + , выкачиваемых из клетки с помощью натриевого насоса, не вполне точно уравновешивается поступлением в клетку ионов К + . В связи с этим часть катионов натрия удерживается внутренним слоем противоионов (анионов) на наружной поверхности клеточной мембраны. Таким образом, на мембранах, ограничивающих нервные клетки, поддерживается разность электрических потенциалов (трансмембранная разность электрических потенциалов); эти мембраны электрически возбудимы.
При возбуждении, вызванном тем или иным агентом, селективно изменяется проницаемость мембраны нервной клетки (аксона): увеличивается избирательно для ионов Na + (примерно в 500 раз) и остается без изменения для ионов К + . В результате ионы Na + устремляются внутрь клетки. Компенсирующий поток ионов К + , направляющийся из клетки, несколько запаздывает. Это приводит к возникновению отрицательного заряда на наружной поверхности клеточной мембраны. Внутренняя поверхность мембраны приобретает положительный заряд; происходит перезарядка клеточной мембраны (в частности, мембраны аксона, т.е. нервного волокна), и возникает потенциал действия, или спайк. Продолжительность спайка не превышает 1 мс. Он имеет восходящую фазу, пик и нисходящую фазу. Нисходящая фаза (падение потенциала) связана с нарастающим преобладанием выхода ионов К + над поступлением ионов Na + – мембранный потенциал возвращается к норме. После проведения импульса в клетке восстанавливается состояние покоя. В этот период ионы Na + , вошедшие в нейрон при возбуждении, заменяются на ионы К + . Этот переход происходит против градиента концентрации, так как ионов Na + во внешней среде, окружающей нейроны, намного больше, чем в клетке после момента ее возбуждения. Переход ионов Na + против градиента концентрации, как отмечалось, осуществляется с помощью натриевого насоса, для работы которого необходима энергия АТФ. В конце концов все это приводит к восстановлению исходной концентрации катионов калия и натрия внутри клетки (аксона), и нерв готов для получения следующего импульса возбуждения. Заметим, что миелиновые мембраны, образуемые шванновскими клетками, окутывают нервные волокна и служат электрическим изолятором. Этот изоляционный слой покрывает большинство нервных волокон и сильно ускоряет распространение электрической волны (сигнала); при этом ионы входят в клетку и выходят из нее только в тех местах, где изолятор отсутствует. Как уже отмечалось, миелиновая мембрана состоит из фос-фолипидов, в частности из сфингомиелина, холестерина, а также белков и гликосфинголипидов. Некоторые заболевания, например рассеянный склероз, характеризуются демиелинизацией и нарушением проведения нервного импульса. Другим не менее важным процессом для нервной ткани является передача нервного импульса от одной нервной клетки к другой или воздействие на клетки эффекторного органа.
Возникновение нервного импульса связано с состоянием мембраны нейрона. Биологические мембраны, будучи липидно-белковым комплексом, непроницаемы для заряженных ионов. Перемещение ионов через мембрану осуществляется с помощью специальных транспортных систем: натриевого насоса, работающего с затратой энергии (Na + , К + -АТФ-аза) и ионпроводящих каналов – натриевых и калиевых.
В состоянии физиологического покоя по разные стороны мембраны имеет место разность концентраций ионов, главным образом Na + и К + . Концентрация ионов К + выше в клетке, ионов Na + – в межклеточной жидкости. Это влияет и на распределение других ионов. Разность концентраций ионов поддерживается с помощью натриевого насоса, перекачивающего ионы против градиента концентрации (в противном случае диффузия ионов по градиенту концентрации привела бы к выравниванию концентраций ионов по обе стороны мембраны и гибели клетки) Таким образом, внутри аксона образуется избыток (–) зарядов, снаружи – (+) зарядов, то есть возникает разность электрических потенциалов – потенциал покоя (ПП). Его значение составляет – 60-70 мВ. ПП одинаков по всей длине нервного волокна (рис. 4).
Раздражение нервного волокна приводит сначала к открытию Na + - и К + - каналов. Раньше открываются Na + -каналы, и в клетку устремляется поток ионов натрия. Это изменяет трансмембранный потенциал: сначала он становится равным 0 (деполяризация мембраны), затем происходит перезарядка мембраны, внутренняя сторона мембраны приобретает (+) заряд, наружная (-). Разность потенциалов при этом достигает 40 мВ. Эта величина и есть потенциал действия (ПД)
Натриевые каналы закрываются, открываются калиевые, начинается выход калия из клетки, потенциал изменяется от – 40 мВ до – 70 мВ, то есть идет реполяризация мембраны. После закрытия ионных каналов ионный насос восстанавливает исходное распределение ионов по сторонам мембраны.
Механизмы памяти.
Концепции биохимического кодирования индивидуального опыта в памяти опираются на две группы фактов:
· возможность передачи приобретенной информации необученному мозгу с помощью этих факторов.
Первые гипотезы, связывающие запечатление информации с биохимическими изменениями в нервной ткани родились на основе широко известных в 60-е гг. опытов Г. Хидена, когда было показано, что образование следов памяти сопровождается изменениями свойств РНК и белка в нейронах.
Выяснилось, что раздражение нервной клетки увеличивает в ней содержание РНК и оставляет длительные биохимические следы, сообщающие клетке способность резонировать в ответ на повторные действия одних и тех же раздражителей.
Таким образом, было установлено, что РНК играет важную роль в механизмах формирования и сохранения следов памяти.
Однако в более поздних работах было показано, что в консолидации энграмм памяти ведущую роль играет ДНК, которая может служить хранилищем не только генетической, но и приобретенной информации, а РНК обеспечивает передачу специфического информационного кода.
В настоящее время идея существования биохимических факторов, способных к сохранению и переносу информации, большинством исследователей воспринимается критически. Считается, что гипотеза молекулярного кодирования индивидуального опыта не имеет прямых фактических доказательств.
Медиаторам - химическим посредникам в синаптической передаче информации - придается большое значение в обеспечении механизмов долговременной памяти. Основные медиаторные системы головного мозга принимают самое непосредственное участие в обучении и формировании энграмм памяти. Так, экспериментально установлено, что уменьшение количества норадреналина замедляет обучение, вызывает амнезию и нарушает извлечение следов из памяти.
Р.И. Кругликов (1986) разработал концепцию, в соответствии с которой в основе долговременной памяти лежат сложные структурно-химические преобразования на системном и клеточном уровнях головного мозга. Показано, что под влиянием обучения увеличивается количество холинорецепторов, то есть рецепторов, расположенных на теле нейрона и отвечающих за обнаружение медиатора ацетилхолина. В процессе образования условного рефлекса повышается чувствительность соответствующих нейронов к ацетилхолину, что облегчает обучение, ускоряет запоминание и способствует более быстрому извлечению следа из памяти. В то же время вещества, препятствующие действию ацетилхолина, нарушают обучение и воспроизведение, вызывая амнезию (потерю памяти).
Биохимические методы, которые позволяют проникнуть в последовательность процессов, происходящих в синаптических мембранах с последующим синтезом новых белков, привлекают многих исследователей памяти. На этом пути ожидаются новые яркие открытия. Предполагается, например, что для различных видов памяти в ближайшем будущем будут выявлены различия в биохимических процессах.
Тем не менее следует подчеркнуть, что интенсивные биохимические исследования привели к явной переоценке и автономизации клеточно-молекулярного уровня изучения механизмов памяти. Как указывает С. Роуз, эксперименты, проводимые только на клеточном уровне, слишком ограничены и, по-видимому, не способны ответить на вопрос, как мозг человека запоминает, например, сложные симфонические партитуры или извлекает из памяти данные, необходимые для разгадывания простого кроссворда.
Для более полного знания специфики функционирования процессов памяти необходим переход на уровень сложных мозговых систем, где многие нейроны соединены между собой морфологическими и функциональными связями. При этом психофизиологические исследования на здоровых людях позволяют изучать процессы переработки и хранения информации, а изучение больных с различного рода амнезиями, возникающими после повреждения мозга, позволяет глубже проникать в тайны памяти.
Березов стр. 641-642
Дата добавления: 2019-07-15 ; просмотров: 727 ;
Большинство исследователей придерживаются мнения, что явления электрической поляризации клетки обусловлены неравномерным распределением ионов К+ и Na+по обе стороны клеточной мембраны. Мембрана обладает избирательной проницаемостью: большей для ионов К+ и значительно меньшей для ионов Na+. Кроме того, в нервных клетках существует механизм, который поддерживает внутриклеточное содержание натрия на низком уровне вопреки градиенту концентрации. Этот механизм получил название натриевого насоса. При определенных условиях резко повышается проницаемость мембраны для ионов Na+. В состоянии покоя внутренняя сторона клеточной мембраны заряжена электроотрицательно по отношению к наружной поверхности. Объясняется это тем, что количество ионов Na+, выкачиваемых из клетки с помощью натриевого насоса, не вполне точно уравновешивается поступлением в клетку ионов К+. В связи с этим часть катионов натрия удерживается внутренним слоем противоионов (анионов) на наружной поверхности клеточной мембраны. Таким образом, на мембранах, ограничивающих нервные клетки, поддерживается разность электрических потенциалов (трансмембранная разность электрических потенциалов); эти мембраныэлектрически возбудимы. При возбуждении, вызванном тем или иным агентом, селективно изменяется проницаемость мембраны нервной клетки (аксона): увеличивается избирательно для ионов Na+(примерно в 500 раз) и остается без изменения для ионов К+. В результате ионыNa+устремляются внутрь клетки. Компенсирующий поток ионов К+, направляющийся из клетки, несколько запаздывает. Это приводит к возникновению отрицательного заряда на наружной поверхности клеточной мембраны. Внутренняя поверхность мембраныприобретает положительный заряд; происходит перезарядка клеточной мембраны (в частности, мембраны аксона, т.е. нервного волокна), и возникает потенциал действия, или спайк. Продолжительность спайка не превышает 1 мс. Он имеет восходящую фазу, пик и нисходящую фазу. Нисходящая фаза (падение потенциала) связана с нарастающим преобладанием выхода ионов К+ над поступлениемионов Na+ мембранный потенциал возвращается к норме. После проведения импульса в клетке восстанавливается состояние покоя. В этот период ионы Na+, вошедшие в нейрон при возбуждении, заменяются на ионы К+. Этот переход происходит против градиентаконцентрации, так как ионов Na+во внешней среде, окружающей нейроны, намного больше, чем в клетке после момента ее возбуждения. Переход ионов Na+против градиента концентрации, как отмечалось, осуществляется с помощью натриевого насоса, для работы которого необходима энергия АТФ. В конце концов все это приводит к восстановлению исходной концентрации катионов калияи натрия внутри клетки (аксона), и нерв готов для получения следующего импульса возбуждения. Заметим, что миелиновые мембраны, образуемые шванновскими клетками, окутывают нервные волокна и служат электрическим изолятором. Этот изоляционный слой покрывает большинство нервных волокон и сильно ускоряет распространение электрической волны (сигнала); при этом ионы входят вклетку и выходят из нее только в тех местах, где изолятор отсутствует. Как уже отмечалось, миелиновая мембрана состоит из фос-фолипидов, в частности из сфингомиелина, холестерина, а также белков и гликосфинголипидов. Некоторые заболевания, например рассеянный склероз, характеризуются демиелинизацией и нарушением проведения нервного импульса. Другим не менее важным процессом для нервной ткани является передача нервного импульса от одной нервной клетки к другой или воздействие на клетки эффекторного органа.
Обмен веществ в организме человека. Белки, аминокислоты, жиры. Катаболизм и анаболизм. Биохимические процессы. Предмет биологическая химия. Вопросы на экзамен и Ответы.
У нас самая большая информационная база в рунете, поэтому Вы всегда можете найти походите запросы
Эта тема принадлежит разделу:
Биологическая химия
Обмен веществ в организме человека. Белки, аминокислоты, жиры. Катаболизм и анаболизм. Биохимические процессы. Предмет биологическая химия. Вопросы на экзамен и Ответы.
К данному материалу относятся разделы:
Похожие материалы:
Ответы на вопросы на экзамен по возрастной психологии, периоды развития человека, школьный, дошкольный возраст, юношество, взросление, старость. Готовая шпаргалка. Предмет возрастная психологя
Основы безопасности жизнедеятельности (ОБЖ) - область знаний, в которой изучаются опасности, угрожающие человеку, закономерности их проявлений и способы защиты от них. Материал с уклоном к Республики Казахстан
Слово - основная структурно-семантическая единица языка, которая служит для именования предметов и их свойств, явлений, отношений действительности. высказывании
Ознакомление с предприятием. Выполнение обязанностей дублёров инженерно-технических работников. Выполнение работ, связанных с выполнением выпускной квалификационной работы. Технологическая подготовка производства
Менеджмент и маркетинг. Учебное пособие. Менеджмент воплощает философию предпринимательства. Методологические основы менеджмента. Внешняя и внутренняя среда организации. Виды менеджмента. Социальные аспекты менеджмента.
Читайте также:
