
Брюшная нервная цепочка у насекомых представлена

Брюшная нервная цепочка – это часть центральной нервной системы насекомых, структуры которой находятся позади головного мозга.
- Строение и положение брюшной нервной цепочки
- Основные модификации брюшной нервной цепочки
- Особенности функционирования брюшной нервной цепочки
- Ссылки
Строение и положение брюшной нервной цепочки


Она представлена несколькими парами нервных узлов (ганглиев), располагающимися в грудном и брюшном отделах тела. Ганглии располагаются попарно, каждая пара соответствует определенному сегменту. [1] Продольно узлы соединены между собой нервными волокнами, носящими название коннективы. Кроме того, на уровне каждого сегмента пары узлов соединяются посредством поперечных волокон, называемых комиссурами. Комиссуры обычно довольно короткие, и поэтому визуально пара узлов может выглядеть как единое образование. [3]
В типичных случаях (у тараканов, прямокрылых) нервная цепочка включает 3 пары грудных узлов и 8 пар брюшных. [1] (фото)
Основные модификации брюшной нервной цепочки
Не у всех насекомых строение брюшного мозга соответствует типичной схеме; можно даже сказать, что те или иные его изменения имеются у большинства. Чаще всего видоизменения этого отдела нервной системы состоят в том, что коннективы между ганглиями укорачиваются, благодаря чему соседние ганглии могут сливаться между собой. Кроме того, нередко наблюдается миграция ганглиев в сегменты, расположенные впереди. [4]
Типичное строение, при котором пара ганглиев цепочки строго соответствует каждому сегменту тела, имеется у многих гусениц и личинок пчел. Однако уже у личинки Malacosoma(бабочки коконопряда) восьмой ганглий брюшка сместился в седьмой сегмент и слился с его нервным узлом. Аналогичным образом, у личинок стрекоз первый брюшной ганглий слит с последним грудным, а у взрослых пчел с заднегрудным ганглием соединены целых два первых брюшных узла.


Наконец, муха Sarcophaga (фото) отличается тем, что у нее все грудные и все брюшные ганглии слиты между собой в одну массу (синганглий), которая находится в груди. В брюшке же нервной цепочки нет, там располагаются только нервы периферической нервной системы, идущие к органам. То же самое произошло у хруща Lachnosterna. [4]
В некоторых случаях у насекомых одного вида, но разного пола брюшная нервная цепочка имеет различное число ганглиев. К примеру, у Человеческой блохи-самца имеется 8 брюшных ганглиев, а у самки их всего лишь 7. [4]
На разных стадиях развития строение нервной системы у одного и того же насекомого также может довольно сильно отличаться. Более того, существует установленная закономерность: у взрослых насекомых брюшная цепочка более концентрирована (т.е., большее число ее ганглиев слиты между собой), чем у личинок. Так, личинка пчелы имеет 10 ганглиев в цепочке, взрослая пчела – только 7. [4] Исключением из этого правила являются некоторые мухи. У личинки львинки цепочка представляет собой один синганглий, а у взрослой мухи в брюшке находится 5 отдельно лежащих нервных узлов. Примерно то же самое наблюдается у муравьиного льва. [4]


Особенности функционирования брюшной нервной цепочки
Каждая пара ганглиев отдает нервные волокна, связывающие ее с чувствительными окончаниями и органами-эффекторами. Эти волокна распространяются в тканях того сегмента, к которому эта пара изначально принадлежит, даже если по факту она в этом сегменте не находится. [2]
Несмотря на принадлежность к конкретному отделу тела, нервные узлы брюшной цепочки соединяются между собой, с элементами симпатической нервной системы и мозга, так что каждый ганглий работает в содружестве с другими. В то же время, нервные узлы в определенной степени автономны. Например, когда во время ужаливания пчела теряет жало (фото), вместе с ним из ее тела вырывается последний ганглий, который его иннервирует. На протяжении некоторого времени жало продолжает функционировать, находясь в ранке. [2]
Класс насекомые лидирует по числу видов среди всех животных. На настоящее время описано около 1,1 млн. видов насекомых, при том факте, что истинное число видов оценивается от 2 до 8 млн. разными исследователями. Можно смело заявить, что половина (скорее всего, гораздо больше) видов насекомых еще не изучены.
"Насекомые. Они - истинные хозяева земли" - сказал В.М. Песков. Это действительно так, люди - редкое исключение в мире насекомых. Именно они сейчас эволюционно достигли наивысшего расцвета, отлично приспособившись к жизни в среде людей. Так что с точки зрения эволюции мы с вами живем в эру господства насекомых, удивительных существ, сложные инстинкты и поведение многих из которых поражает.

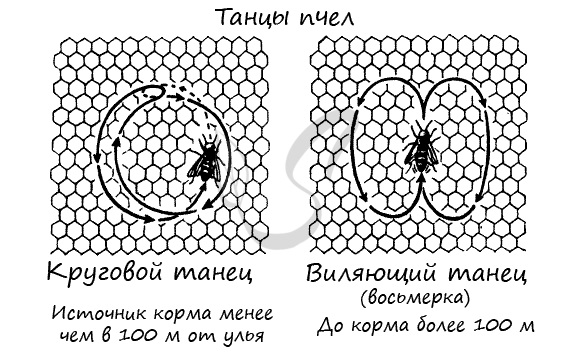
С помощью танца пчелы могут сообщать друг другу, в каком направлении и как далеко от их местоположения находится корм. Если расстояние менее 100 метров, пчела исполняет круговой танец, а если более 100 метров - виляющий танец, в виде восьмерки. Только насекомым свойственна общественная организация, разделение труда между особями.
Насекомых изучает интереснейшая наука - энтомология (от греч. entoma - насекомые и logos - слово, учение), в этой статье мы познакомимся с их общим строением.
- Покровы тела, опорно-двигательная система
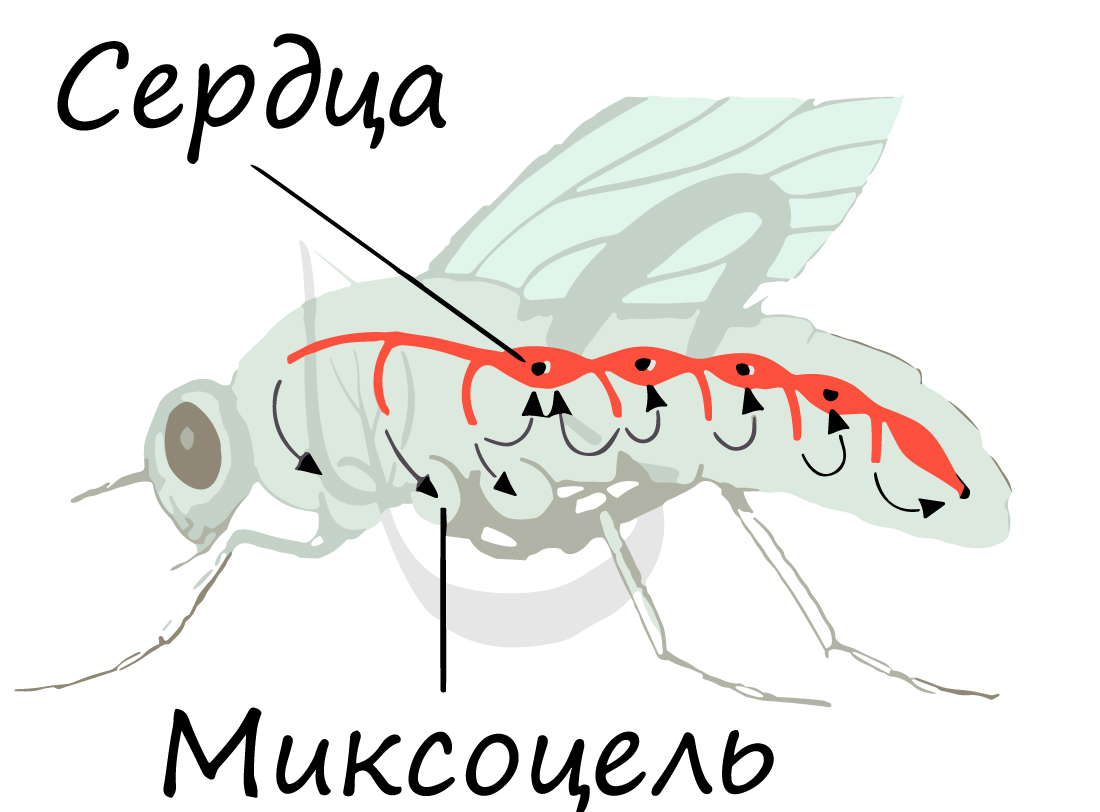
Тело дифференцировано на голову (5 слившихся сегментов), грудь (3 сегмента) и брюшко (8 сегментов). На голове находится одна пара усиков - антенны, являющиеся органами обоняния и осязания. Полость тела насекомых смешанная (миксоцель), она позволяет во время линьки значительно увеличивать объем тела за счет увеличения давления крови.
Многие насекомые способны к удивительному движению в воздухе - полету. Первая пара крыльев носит названия надкрылья: в полете они не участвуют, это плотные хитинизированные образования, прикрывающие часть груди и брюшка. Вторая пара крыльев принимает непосредственное участие в полете, имеет вид уплощенных перепончатых образований.
Три пары ходильных ног крепятся к груди. Членистая конечность насекомого оканчивается двумя коготками, между которыми иногда располагаются присоски. Конечности насекомых разнятся по выполняемой функции, в соответствии с ней получая свои названия: копательная, бегательная, прыгательная, плавательная, собирательная.

Тело насекомых, как и всех членистоногих, покрыто хитиновой кутикулой - наружным скелетом. Эта плотная оболочка насекомого сдерживает рост. Запомните, что насекомые активно растут только в личиночной стадии и в период линьки, когда хитиновый покров до конца не сформирован или сброшен.

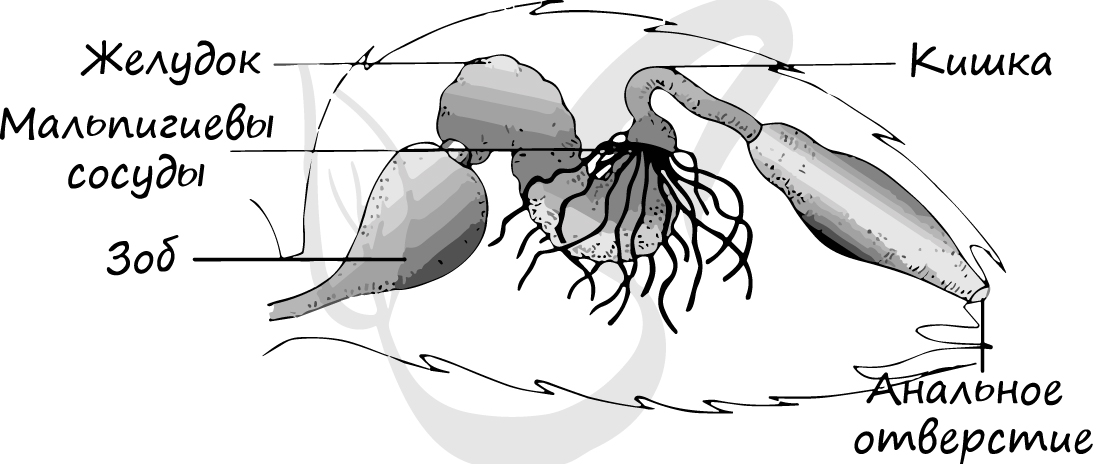
Состоит из переднего, среднего и заднего отделов. К переднему отделу относятся рот, глотка, пищевод, который часто имеет расширение - зоб, желудок. После желудка начинается средний отдел - кишечник, от которого отходят многочисленные слепо заканчивающиеся выросты, увеличивающие всасывательную поверхность. В заднем отделе кишечника происходит формирование экскрементов и всасывание воды, заканчивается задняя кишка анальным отверстием.
Особо необходимо отметить развитую мускулатуру желудка, который называется - мускульный. В нем происходит дополнительное перетирание пищи. После этого пищевые частицы расщепляются до мономеров, которые всасываются кишкой и попадают в гемолимфу. С ее током питательные вещества достигают внутренних органов и тканей.
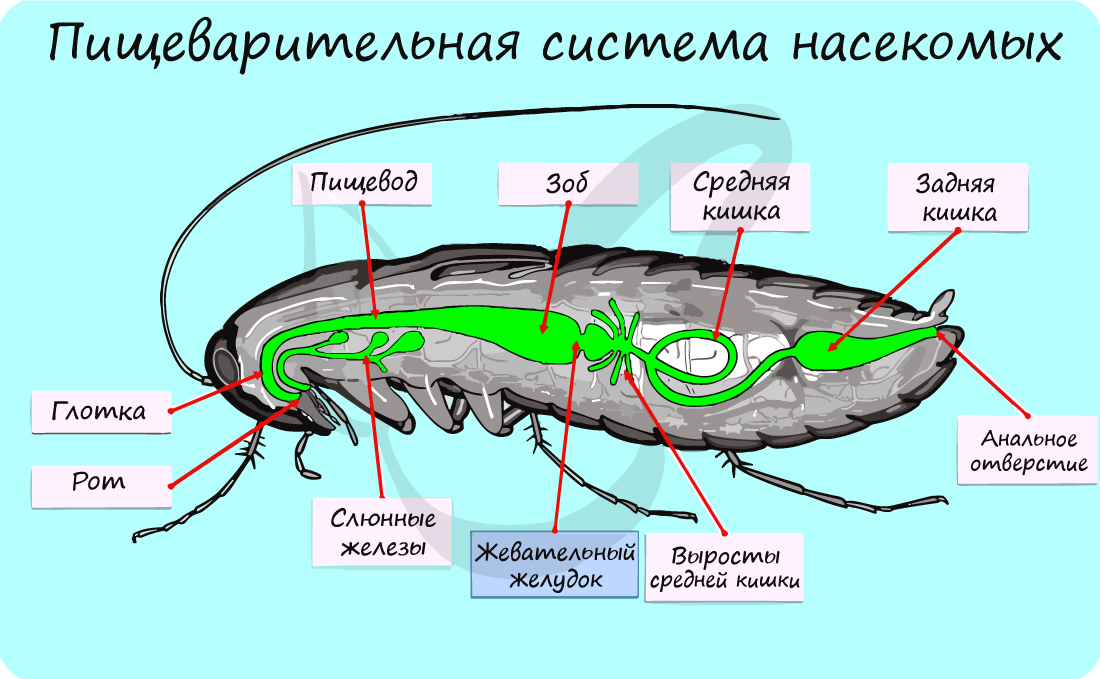
У большинства насекомых имеются слюнные железы. Насекомые обладают самыми разнообразными сложноустроенными ротовыми аппаратами. Строение ротового аппарата отражает способ питания. Ниже вы видите таблицу, отражающую многообразие ротовых аппаратов у насекомых.
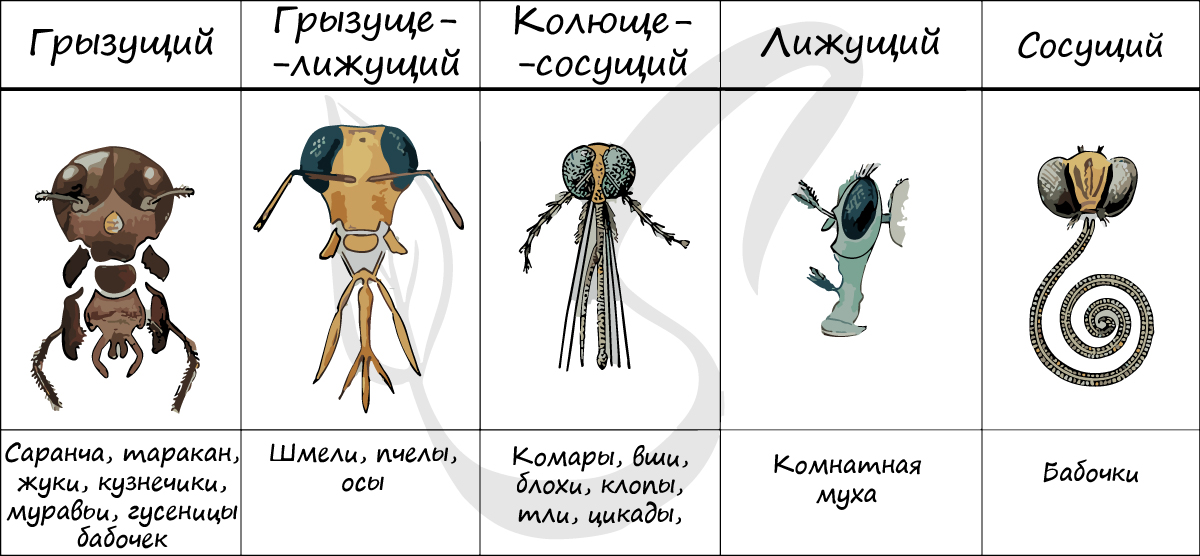
Дыхательная система представлена сильно разветвленной системой трахей, которые выполняют функцию наружного дыхания. На головогруди и брюшке у насекомых находятся дыхальца (стигмы) - дыхательные отверстия, которыми трахеи открываются во внешнюю среду.
Кровеносная система не переносит кислород, так что функция его доставки целиком принадлежит трахеям, которые ветвятся на тонкие трубочки (трахеолы) и подходят к небольшим группам клеток. У части быстролетающих насекомых (мухи, пчелы) трахеи образуют расширенные участки - воздушные мешки, которые улучшают вентиляцию трахейной системы и уменьшают удельный вес тела
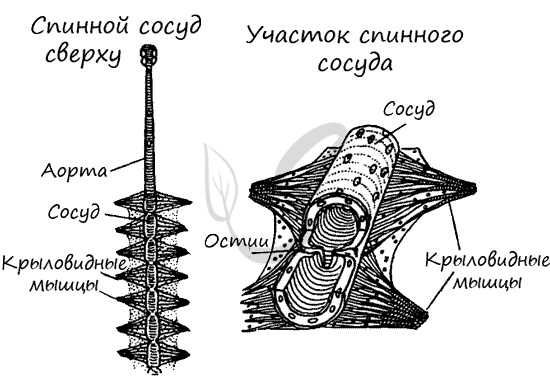
Для насекомых характерен незамкнутый (лакунарный) тип кровеносной системы. Кровь свободно движется по лакунам (синусам), непосредственно омывая внутренние органы и ткани. Функцию сердца выполняет спинной сосуд: благодаря его сокращениям кровь перекачивается из задней части тела в переднюю.
Функционирование сосуда-сердца схоже с таковым у ракообразных. В момент расслабления сосуда-сердца через отверстия (остии) кровь наполняет его, а в момент сокращения (систолы) кровь выталкивается в артерии, затем попадает в полость тела, омывает органы и ткани.
Внутреннюю среду насекомых составляет гемолимфа, представляющая собой бесцветную или желтоватую жидкость. В гемолимфу из кишечника всасываются питательные вещества, после чего доставляются к клеткам организма. В нее же удаляются побочные продукты обмена веществ.
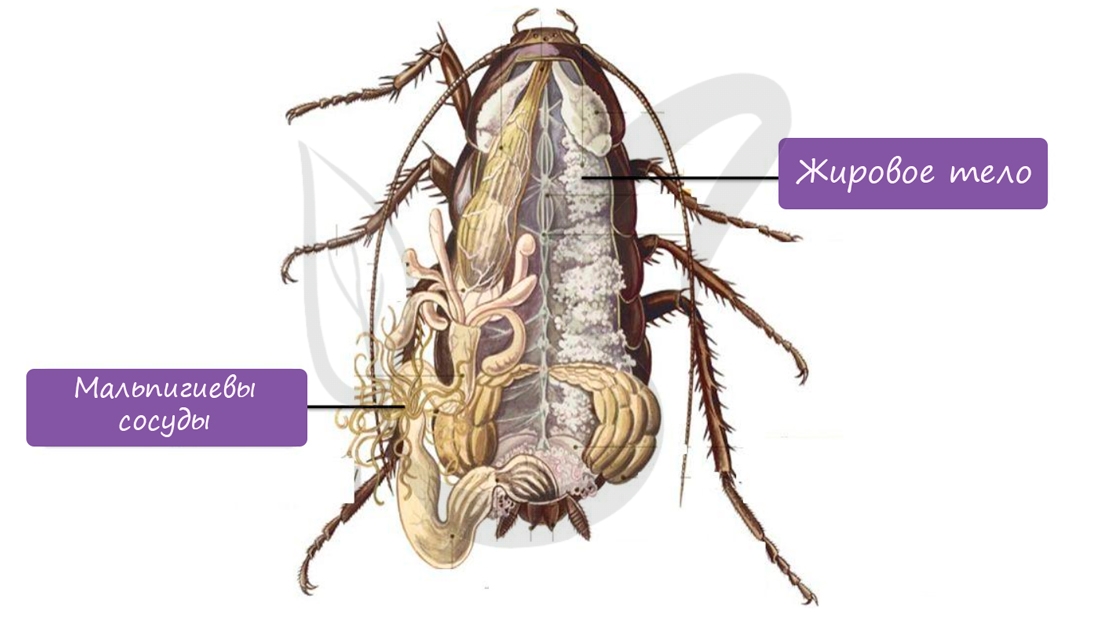
Органы выделения представлены мальпигиевыми сосудами (в честь итал. биолога и врача - Марчелло Мальпиги). Это длинные трубчатые выросты насекомых и паукообразных, которые расположены на границе средней и задней кишки.
Как вы помните, перед насекомыми стоит сложная задача: максимально сохранить воду в организме. Мальпигиевы сосуды этому способствуют: в них поступают продукты обмена веществ из гемолимфы в виде суспензии. По мере продвижения по мальпигиевым сосудам, из суспензии всасывается вся вода обратно в гемолимфу, а продукты обмена веществ (кристаллы мочевой кислоты) в сухом виде поступают в кишку и выводятся из организма с экскрементами.
К органам выделения также относится жировое тело. Жировое тело - образование мезодермального происхождения, содержащие запасы питательных веществ, которые постоянно расходуются организмом. В жировом теле могут накапливаться и продукты обмена веществ: продукты распада, что нейтрализует их токсическое действие.
Тип нервной системы насекомых - узловой. Состоит она из головного мозга (надглоточного ганглия), подглоточного ганглия и брюшной нервной цепочки.
Головной мозг имеет сложное строение, образован в результате слияния 3 ганглиев и состоит соответственно из 3 отделов: переднего, среднего и заднего. От мозга отходят нервные тяжи - коннективы, которые направляются к подглоточному ганглию, в совокупности образуя окологлоточное нервное кольцо.
Наиболее развитые ганглии в брюшной нервной цепочке находятся в груди, так как они иннервируют сложную работу конечностей и крыльев. Узлы распределены неравномерно: 3 ганглия находятся в груди, 8 - в брюшке.
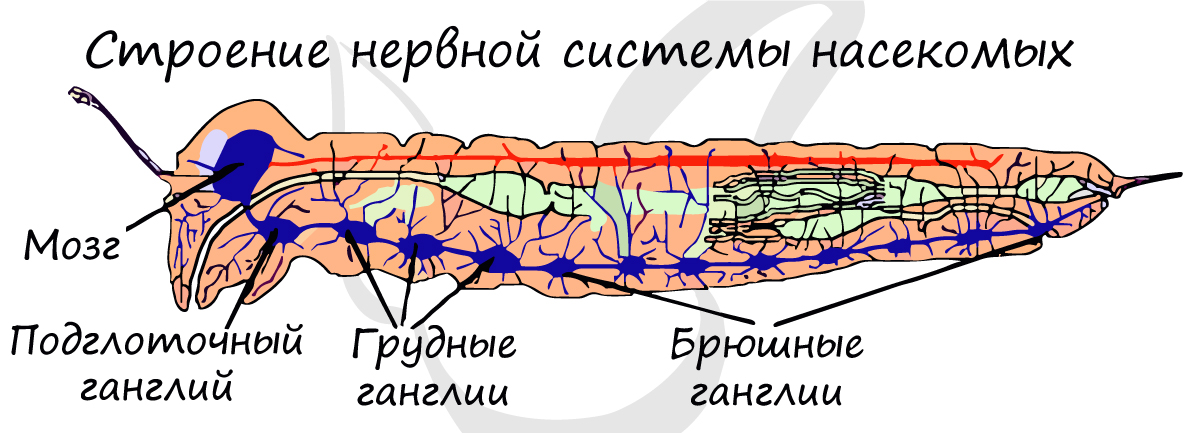
Органы чувств развиты хорошо. Глаза простые или сложные (фасеточные), одна пара усиков (антенн), на которых располагаются органы обоняния и осязания. Имеются органы вкуса, локализующиеся на щупиках нижней губы и нижней челюсти.

Такое прогрессивное развитие нервной системы заложило фундамент для появления у насекомых сложнейших и удивительных рефлексов. Среди всех беспозвоночных только насекомые отличаются общественным (социальным) образом жизни: они совместно строят гнездо, ухаживают за потомством, разделяют обязанности среди членов семьи. Общественными насекомыми являются пчелы, осы, муравьи, шмели.
Заметим, что в переднем отделе мозга расположены грибовидные тела, ассоциативные центры головного мозга. Особенно хорошо развиты грибовидные тела у насекомых, ведущих общественный образ жизни, что связано с их сложным поведением.

Насекомые раздельнополы, гермафродиты среди них встречаются очень редко. Часто встречается хорошо выраженный половой диморфизм - внешние различия между самцом и самкой.
Половые железы парные: у мужских особей - семенники, у женских - яичники. От семенников и яичников соответственно отходят семяпроводы и яйцеводы, впадающие в семяизвергательный канал и влагалище. Оплодотворение у насекомых внутреннее: с помощью совокупительных органов семя вводится самцом в половые пути самки.

Развитие может быть прямым или непрямым. Запомните, что у всех насекомых развитие непрямое.
Непрямое развитие может протекать с метаморфозом (от греч. metamorphosis - превращение) - полное превращение, или без него - неполное превращение.
Метаморфоз - глубокое преобразование строения организма, переход из одной формы в другую, сопровождающийся появлением новых элементов строения и функций.

Как заметно из схемы выше, неполное и полное превращение отличаются наличием стадии куколки, это именно та стадия, в которую и происходит метаморфоз. О том, для каких насекомых характерно развитие с метаморфозом, а для каких нет - вы узнаете из следующей статьи.
Классический пример метаморфоза - превращение гусеницы в бабочку. В коконе (стадия куколки) в организме гусеницы происходит растворение практически всех тканей, за исключением нервной и кровеносной систем. В результате такого метаморфоза образуется новый организм - бабочка, сильно отличающаяся от гусеницы.

Логично предположить, что у насекомых с неполным превращением личинка напоминает взрослую особь, но меньше ее в размерах. У насекомых с полным превращением, которое сопровождается метаморфозом (гусеница становится бабочкой), личинка совершенно не похожа на взрослую особь, разительно отличается от нее по строению и функциям.
Особо необходимо отметить партеногенез. Партеногенез (от греческого parthenos - дева, девственница и genesis - рождение) - одна из форм полового размножения, при котором новый организм развивается из яйцеклетки без ее оплодотворения. Поскольку мужская гамета не участвует в данном процессе, потомство генетически идентично матери.
Партеногенез встречается у следующих насекомых: тли, муравьи, пчелы, осы, шмели, тутовый шелкопряд. Партеногенез относится именно к половому (а не бесполому) типу размножения, поскольку новый организм развивается из неоплодотворенной яйцеклетки (женской гаметы). Данный процесс играет важную роль: он значительно увеличивает темпы роста популяции, регулирует соотношение женских и мужских особей, обеспечивает продолжение существования вида.

Искусственный партеногенез у тутового шелкопряда впервые был получен А.А. Тихомировым в 1886 году, а практика развита Б.Л. Астауровым, который придумал искусственный способ получения самцов тутового шелкопряда, дающих повышенный выход ценного материала - шелкового волокна высокого качества.

Значимость насекомых для человека трудно переоценить. Они выполняют множество полезных функций:
- Являются опылителями цветковых растений, в числе которых много культурных видов, употребляемых человеком в пищу
- Являются звеном в цепи питания (консументы)
- Регулируют численность других насекомых
- Участвуют в почвообразовании: способствуют разложению растительных останков, прокладывают в почве ходы, роют норки
- Производство шелка: как и 4000 лет назад, сегодня для получения шелка используют коконы тутового шелкопряда
- Способствуют разложению останков животных: мясные мухи питаются падалью (трупами животных)
- Производство меда: почти во всех странах мира разводят медоносных пчел (пчеловодство)
Некоторые насекомые определенно приносят человеку больше вреда, чем пользы:
- Часть является кровососущими эктопаразитами человека и животных - комары, вши, клопы, блохи
- Переносят инфекционные заболевания - малярия (комары), лейшманиозы (москиты), сыпной и возвратный тиф (вши), чума (блохи), дизентерия (мухи, тараканы)
- Личинки насекомых, а также взрослый особи (имаго) наносят значительный вред сельскохозяйственным культурам (саранча, жуки, тли)

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
1. Краткие сведения об анатомии и гистологии нервной системы
По особенностям организации и функции нервная система насекомых может быть подразделена на центральную, симпатическую и периферическую.
Центральная нервная система имеет метамерное строение: она состоит из серии нервных узлов, или ганглиев, соединенных в нервную цепочку межсегментными стволами - коннективами. Каждый ганглий парный, его левая и правая половины соединены более или менее выраженными комиссурами. Передний отдел нервной цепочки из надглоточного и подглоточного ганглиев называют головным мозгом, а остальные ганглии - брюшной нервной цепочкой. Число ганглиев в брюшной цепочке у разных филогенетических групп насекомых и даже разных стадий их индивидуального развития различно, однако степень интеграции ганглиев, как правило, не коррелирует с уровнем поведения насекомого.
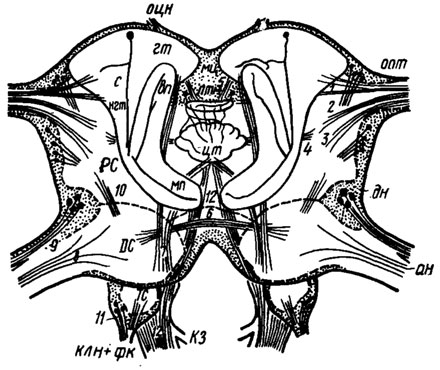
Рис. 1. Схематизированная картина фронтального среза через надглоточный ганглий сверчка Gryllus campestris(по Huber, 1960). Границы нейропиля прото-(PC), дейто-(ДС) и тритоцеребрума (ТС) обведены прерывистой линией. Ассоциативные ядра протоцеребрума обведены сплошными линиями. ан - антеннальный нерв; вп - возвратный придаток ножки грибовидного тела: гт - грибовидное тело; дн - двигательные нейроны дейтоцеребрума; клн+фк - клинео-лабральный нерв и фронтальный коннектив; кЗ - комиссура тритоцере-брума; мп - медиальный придаток ножки грибовидного тела; мц - межцеребральная часть; нгт - ножка грибовидного тела; опт - оптические доли; оцн - оцеллярный нерв; цт - центральное тело; птм - протоцеребральный мост. Нервные пути: 1-3 - оптический тракт; 4 - оптическая комиссура; 5 - оцеллярный тракт; 6 - комиссура дейтоцеребрума; 7- ольфакторно-глобулярный тракт; 8 - афферентные нервы антенн; 9 - эфферентные аксоны, идущие к мышцам антенн; 10 - связь между прото- и дейтоцеребрумом; 11 - моторные аксоны, идущие к верхней губе; 12-нервные пути, связывающие надглоточный ганглий с подглоточным
Надглоточный ганглий - самый крупный и наиболее сложный нервный узел, сходный по функциям с головным мозгом позвоночных. Его образуют, по крайней мере, три слившихся сегментальных ганглия: прото-, дейто- и тритоцеребрум. Более крупный протоцеребрум (рис. 1, PC) тесно связан с фасеточными глазами (или стеммами у личинок Holometabola) и дорсальными глазками, а его грибовидные тела (гт) и примыкающие к ним мозговые ядра служат высшим ассоциативным центром, где замыкаются условные связи. Грибовидные тела наиболее развиты у общественных насекомых, ведущих, в сравнении с остальными, более сложный образ жизни и обладающих высокой способностью к научению. Двустороннее разрушение грибовидных тел влечет выпадение приобретенных навыков, но безусловные реакции на те же стимулы у насекомого сохраняются. Дейтоцеребрум (DC) связан с антеннами, их сенсорным аппаратом и мышцами, поэтому считается обонятельным центром. Миниатюрный тритоцеребрум (ТС) иннервирует верхнюю губу, а также связывает надглоточный ганглий с симпатической нервной системой.
Подглоточный ганглий, также состоящий из трех сегментальных нервных масс, обслуживает сенсиллы и мышцы ротовых частей - соответственно мандибул, максилл и лабиума, а также рецепторы и эффекторы шейной области.
Брюшная нервная цепочка начинается тремя грудными ганглиями, которые у одних насекомых лежат раздельно, у других - сливаются вместе (иногда только мезо- и метаторакальный ганглии) или даже объединяются с несколькими (и изредка со всеми) брюшными ганглиями. Грудные ганглии ответственны в первую очередь за управление движениями ног и крыльев. В них могут также замыкаться условные локомоторные рефлексы.
Менее сложно устроены брюшные ганглии, число которых не превышает 8, иннервируют мышцы брюшных сегментов и их придатков и получают от них сенсорные нервы. Представление о том, в каких комбинациях могут сливаться ганглии брюшной нервной цепочки, дает рис. 2.
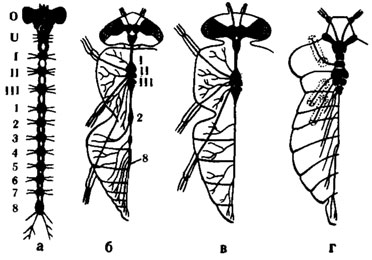
Рис. 2. Различные ступени концентрации центральной нервной системы насекомых(по Eidmann, Kuhlhorn, 1970): а - гипотетическое исходное состояние; б - слепень Tabanus bovinus; в - муха Sarcophaga; г - клоп семейства Pentatomidae. О - надглоточный ганглий; U - подглоточный ганглий; I-III - грудные ганглии; 1-8 - брюшные ганглии
Симпатической, или вегетативной, нервной системы, регулирующей работу внутренних органов, и связанного с ней нейросекреторного аппарата насекомых мы практически не будем касаться в дальнейшем изложении.
Периферическая нервная система состоит из афферентных и эфферентных нервов, чувствующих клеток со вспомогательным аппаратом и эфферентных окончаний. Наше последующее описание будет посвящено в основном именно этой части нервной системы.
Основная структурная единица нервной системы насекомых, как и всех высокоорганизованных животных, - нервная клетка, или нейрон. От тела нейрона, в котором заключено ядро, отходят обычно один длинный неразветвленный аксон и несколько коротких ветвящихся дендритов. В зависимости от числа и взаимного расположения этих отростков говорят о моно-, би- и мультиполярных нейронах. Дендрит служит входом нейрона, а аксон - выходом. Морфологически и функционально различают три главных типа нейронов: чувствующие, двигательные и вставочные. Нас в первую очередь будут интересовать чувствующие и вставочные.
Чувствующие нейроны. Тела чувствующих, или сенсорных, клеток обычно биполярной или мультиполярной формы * лежат всегда вблизи чувствующего органа или иннервируемой ткани. Дендриты одних нейронов, чаще всего биполярных, связаны с кутикулярными образованиями, других, всегда мультиполярных, - с тканями полости тела или же они образуют субэпидермальную сеть, как у личинок с мягкой кожей.
* ( Некоторые авторы относят чувствующие клетки к особой категории нервных клеток и не называют их нейронами.)
Соответственно различают две большие категории чувствующих клеток. Клетки первого типа отличаются тем, что практически всегда связаны с кутикулой или ее впячиваниями: аподемами, трахеями, выстилкой предротовой и ротовой полостей и т. п. К ним принадлежат разнообразные экстерорецепторные клетки, в том числе и зрительные, хотя их дендриты выражены неясно. Клетки второго типа никогда не связаны с кутикулой и лежат только на внутренней поверхности тела, стенках пищеварительного тракта, в мышечной и соединительной тканях. Электрофизиологически показано, что они принадлежат интеро или проприоцепторам.
Аксоны чувствующих клеток идут непосредственно в соответствующие ганглии ЦНС, иногда находящиеся непосредственно в головном мозгу, например оптические или обонятельные центры. Вопрос о каналах связи рецепторных клеток с нервным центром чрезвычайно важен для правильной интерпретации работы анализатора и механизма управления поведением насекомого. Теперь, по-видимому, все признают несостоятельным прежнее мнение о том, что в некоторых рецепторных системах, например в антеннах клопа Rhodnius, имеет место срастание аксонов нескольких чувствующих клеток в единое волокно. Но замыкание группы рецепторов на один периферический нейрон второго порядка, т. е. потеря "адреса" входного сигнала, характерно для первого оптического ганглия насекомых. Смысл такого способа связи с центром, ведущего к частичной потере информации от совокупности датчиков, пока не всегда ясен (см. ниже).
Нервная ткань, в том числе и сенсорные клетки, происходят из эктодермы. Их принадлежность покрову тела выражается и в том, что связь чувствующего органа с ЦНС устанавливается центростремительно. Так, В. Вигглесворс показал на клопе Rhodnius, что перерезанный афферентный нерв регенерирует в направлении к ЦНС. Точно так же во время каждой линьки, когда образуются дополнительные рецепторы, чтобы обслуживать увеличивающуюся поверхность тела, их чувствующие клетки посылают аксоны центростремительно.
Выявленный на гистологических препаратах факт центростремительного развития аксона может стать одним из оснований для важного заключения о том, что путь от чувствующей клетки до ЦНС прямой, без синаптического переключения. Вблизи рецепторных клеток и афферентных нервов встречаются другие, например, нейроглиальные (питающие) клетки, но они не имеют отношения к передаче рецепторного сигнала.
Двигательные (моторные) нейроны. Тела этих обычно униполярных нейронов лежат всегда в сегментальном ганглии, чаще с его вентральной стороны и с боков. Их дендриты густо ветвятся в нейропиле ганглия, вступая в синаптические контакты с вставочными нейронами или даже непосредственно с разветвлениями аксонов чувствующих клеток. Аксоны двигательных нейронов уходят за пределы ганглия к мышцам, где заканчиваются множеством моторных концевых пластинок, или бляшек. Одну и ту же мышцу иннервируют так называемые быстрые и медленные двигательные волокна: по первым идут импульсы, вызывающие быстрые сокращения мышцы, по вторым - вызывающие тоническое сокращение. При этом тело иннервирующего двигательного нейрона может лежать в ганглии как того же сегмента, которому принадлежит мышца, так и соседнего. Часто встречается контралатеральное положение тела двигательного нейрона по отношению к эффектору.
Вставочные нейроны, или интернейроны, выполняют функцию передачи возбуждения с одного нейрона на другой. Они связывают чувствующие клетки с моторными, причем в последовательную цепь может быть включено несколько клеток, и тогда говорят об интернейронах 2-го, 3-го и более высокого порядка. Их обычно униполярные небольшие (от 2-3 мкм и меньше) тела лежат всегда по периферии ганглия, а отростки, в числе которых аксон, нередко т-образный, вступают внутрь ганглия, в его нейропиль. Интернейроны могут быть сегментальными или мультисегментальными, с восходящими и нисходящими волокнами. Последние участвуют, например, в управлении локомоторными реакциями. Благодаря множеству связей интернейроны образуют коммутационные сети для выработки нервных команд, направляемых по специальным трактам в составе комиссур и коннектив.
Среди восходящих волокон у насекомых обращают на себя внимание аксоны так называемых гигантских интернейронов, в частности, берущие начало в последнем брюшном ганглии и идущие по цепочке до грудных и даже до головных центров. Они отличаются большой длиной и толщиной (до 30 мкм и более), а главное, высокой скоростью проведения сигналов. Так, если в обычных афферентных двигательных нервах импульсы распространяются со скоростью 1,5-3,0 м/с, то в гигантском волокне, например таракана Periplanefa, скорость проведения может достигать 5-7 м/с. Считалось, что с помощью таких "быстрых" интернейронов осуществляется реакция убегания, когда опасность грозит насекомому сзади, но теперь этот взгляд признан несостоятельным.
Очень сложна область ганглиозного нейропиля, в котором генерируются нервные команды, управляющие поведением насекомого. Ее образует переплетение отростков дендритов моторных нейронов с разветвлениями ассоциативных и афферентных волокон, связанных друг с другом множеством синаптических контактов. Волокна здесь особенно тонкие: от 0,1 мкм по сравнению 1-3 мкм на выходе из дорсальных (двигательных) и вентральных (чувствующих) корешков. Они содержат протонейрофибриллы диаметром 100-200 А, митохондрии, гранулярный эндоплазматический ретикулум, нейросекреторные включения и прочие элементы. Синаптическая передача основана на поперечных и продольных контактах волокон или на контакте типа терминальной пуговки. Ультраструктура синаптических образований напоминает таковую синапсов у позвоночных: пресинаптическое волокно в районе контакта содержит синаптические пузырьки, наполненные химическим медиатором. Между трехслойными пре- и постсинаптическими мембранами находится синаптическая щель шириной 100-300 А. Как полагают, у насекомых имеется не только холинэнергический, но и адренэнергический механизм синаптической передачи. Не исключается также возможность электрической передачи возбуждения с помощью соответствующих синапсов, в которых импульсы передаются в результате электрического воздействия пресинаптического тока на мембрану постсинаптического волокна. Такие электрические сикапсы недавно обнаружены между зрительными интeрнейронами - детекторами движения, где они обеспечивают максимально высокую скорость проведения сигналов.
Читайте также:
