
Что представляет собой нервный центр
Определение понятия нервного центра
Нервный центр — это совокупность связанных между собой нейронов, совместно выполняющих определённую функцию путём преобразования входящего возбуждения в выходящее с изменёнными характеристиками.
Данное определение содержит 7 критериев нервного центра, найдите и назовите их.
Суперкраткое определение: Нервный центр - это "полисинаптический трансформатор возбуждения".
Нервный центр - это та нервная структура, которая связывает сенсорную систему с эффекторной и преобразует сенсорное возбуждение в эффекторное или модулирующее.
Нервный центр - это многозначное понятие.
Анатомический подход: нервный центр - это совокупность сходных нервных клеток, выполняющих общую функцию и компактно расположенных в определенном участке ЦНС.
Здесь используется морфологический подход, т.е. нервные центры определяются по строению. Нервные клетки, образующие такой нервный центр, соединены в локальные компактные структуры: нервные узлы (ганглии) в периферической нервной системе или мозговые ядра в центральной нервной системе.
Физиологический подход (по деятельности) : нерный центр - это система взаимосвязанных клеток, которые объединяются для выполнения определённой функции, а физически могут находится в различных местах нервной системы.
Такое определение созвучно понятию "функциональной системы", которое было предложено П.К. Анохиным. Но если функциональная система подразумевает временное объединение нейронов, то нервный центр - это обычно устойчивое образование. По П.К. Анохину различные анатомические нервные центры при необходимости могут на время объединяться в функциональную систему для получения определенного полезного результата.
Нервные центры биорегуляции. В функциональном отношении нервный центр также может представлять собой сложное объединение нескольких анатомических нервных центров, расположенных в разных отделах ЦНС и обусловливающих сложные рефлекторные акты. В этом смысле говорят о "пищевом нервной центре", "болевом нервном центре" и т.п. Это центры регуляции функций организма.
Понятие нерный центр может также пересекаться с понятием доминанты. Доминанта - это устойчивый очаг возбуждения, подчиняющий себе другие очаги . Кроме единичного очага возбуждения в состав доминанты также могут входить и другие связанные с ним очаги. Создатель учения о доминанте А.А. Ухтомский называл такие объединения возбуждённых очагов "созвездиями" ("констелляциями").
Эволюция выбирает путь концентрации нервных элементов и увеличение количества связей между ними. Поэтому из рассеянных диффузно нервных клеток образуются компактные нервные центры.
Линия эволюционного развития структуры нервной системы
1) Отдельные нервные клетки соединены примерно равными связями.
2) Нервные цепи - нервные клетки соединены последовательно более сильными связями, образуя предопределённый путь для движения возбуждения от одного пункта к другому.
3) Нервные сети - нервные клетки соединены в виде решеток с неравноценными связями.
4) Нервные узлы (ганглии) - нервные клетки собраны в компактные структуры, соединёнными между собой продолными связями (коннективами) и поперечными (комиссурами) в виде лестницы.
5) Нервная трубка - нервные клетки расположены в виде сплошных слоёв, закрученных в виде трубы.
6) Нервные ядра - обособленные нейронные структуры из тесно связанных сходных между собой нейронов в составе нервной трубки, специализирующиеся на определённых функциях.
7) Нейронные поля — зоны коры головного мозга. Они состоят из вертикальных колонок, в которые сгруппированы нейроны.
Свойства нервных центров
Наличие входов и выходов для возбуждения . В нервном центре можно различить приносящие (афферентные) входы и выносящие (эфферентные) выходы.
Одностороннее проведение возбуждения. Это свойство отдельного синапса и нервной цепи. В нервном центре может быть множество путей между входами и выходами. За счёт обратных связей возможно возвратное движение возбуждения. Но это происходит внутри нервного центра. А если рассматривать нервный центр целиком, то возбуждение приходит внего по приходящим путям, а выходит по эфферентным выходящим. Таким образом, можно говорить об одностороннем проведении возбуждения нервным центром.
З адержка ( замедление ) проведения возбуждения . В нервных центрах возникает задержка в проведении возбуждения, так называемый латентный (скрытый) период. Задержка обусловлена синаптической передачей возбуждения. Чем больше синапсов участвует в проведении возбуждения, тем более длительной получается задержка.
Суммация возбуждения . Если одновременно подавать возбуждение на несколько входов нервного центра, то на выходе можно получить более сильное возбуждение. Свойством суммации обладает и отдельный нейрон за счёт суммации локальных потенциалов.
Трансформация (преобразование) входящего возбуждения в иное - выходящее . Нервный центр осуществляет изменение, перекодирование поступающих в него потоков импульсов. Трансформация возбуждения - это, пожалуй, самое главное свойство нервного центра. Наиболее известное свойство из этого ряда – трансформация ритма. Нервный центр получает на входе один ритм импульсации, а на выходе дает другой (более медленный или более частый).
Последействие (облегчение) . Это означает, что после возбуждения нервного центра он некоторое время ещё сохраняет повышенную возбудимость. Поэтому последующее возбуждение даёт более сильный эффект и получение эффекта от работы нервного центра облегчено.
Утомляемость и низкая лабильность. Лабильность - это предельная частота импульсации, доступная данной нервной структуре. Нервные центры могут пропускать через себя потоки возбуждения с ограниченной частотой импульсации вследствие задержки передачи возбуждения, которая происходит в многочисленных синапсах. Повышенная утомляемость нервных центров объясняется высокой утомляемостью синапсов и ухудшением метаболизма (обменнных процессов) в нейронах после нагрузки.
Тонус . Это означает, что даже без внешнего воздействия нервный центр сохраняет определённый уровень возбудимости и самостоятельно поддерживает у себя определённый уровень возбуждения.
Чувствительность к кислороду и к действию биологически активных веществ (нейротропных). Это создаёт предпосылки к хеморегуляции - химическому управлению деятельностью нервного центра. Например, усиление или ослабление кровоснабжения изменяет работу нервных центров.
В озбудимость ( возбуждение ) . Это способность нервных центров переходить в более возбуждённое состояние, например, при внешнем воздействии на них (стимуляции) или под влиянием других нервных центров.
Торможение ("тормозимость") . Это способность нервных центров переходить в менее возбуждённое состояние, например, при внешнем воздействии на них или под влиянием других нервных центров.
Иррадиация возбуждения . Это "растекание возбуждения" по нервному центру, распространение возбуждения на новые участки от места его первоначального появления.
Конвергенция (схождение) . Это объединение двух или нескольких входящих потоков возбуждения в один выходящий поток. Т.е. в нервный центр входит больше потоков возбуждения, че выходит из него.
Дивергенция (расхождение) . Это разделение входящего потока возбуждения на несколько выходящих потоков. За счёт дивергенции получается, что в нервный центр входит меньше потоков возбуждения, чем выходит из него.
Окклюзия (запирание) . Это блокирование одним из входящих потоков возбуждения другого входящего потока. В результате выходящий поток возбуждения получается слабее, чем сумма этих входящих потоков.
Индукция (отдача) . Это наведение противоволожного (возбуждённого или тормозного) состояния на другие нервные центры или на себя самого. Для понятия индукции очень важно, что данной структурой наводится именно противоположное состояние, а не то, в котором находится она сама. Так, возбуждённая структура индуцирует торможение, а заторможенная - возбуждение.
Автоматия (спонтанная активность, автономность) нервных центров. Это означает, что даже без внешнего воздействия нервный центр может самостоятельно порождать возбуждение на выходе или поддерживать свой тонус (как бы развлекать сам себя). Объясняется это свойство нервного центра существованием в нём специальных нейронов-пейсмекеров (водителей ритма). В них самопроизвольно возникает возбуждение, независимо от работы их афферентных входов. Таким образом, в нервных центрах может происходить периодическая или постоянная генерация (порождение) нервных импульсов, которые возникают даже при отсутствии входящего возбуждения. Самопроизвольная импульсация пейсмекеров обусловлена колебаниями процессов метаболизма в нейронах и действием на них гуморальных факторов.
Реципрокные (взаимоисключающие) отношения. Это означает, что возбуждение одного нейрона (или центра) подавляет работу другого, связанного с ним, нейрона (или центра).
П ластичность . Это способность перестраивать свою структуру и\или деятельность под влиянием предыдущей деятельности. Пластичность - это одно из важнейших свойств биологических систем, которое отличает их от технических систем.
Адаптация. Нервный центр способен приспосабливаться к новой нагрузке и новым условиям работы.
Компенсаторные возможности. При частичном повреждении нервный центр продолжает свою деятельность за счёт сохранившихся нейронов. Для этого он использует свои способности к пластичности и адаптации.
Основные принципы в работе нервных центров
Принцип общего конечного пути ("воронка Шеррингтона"). Как правило, центры имеют больше афферентных входов, чем эфферентных выходов. Поэтому входящие потоки возбуждения конкурируют за выход, имея общий конечный путь. В итоге количество афферентных входов превышает количество эфферентных выходов.
Принцип обратной связи. Это означает, что последующий элемент (нейрон или центр) в последовательной цепи взаимосвязанных элементов влияет на состояние предыдущего элемента. Обратная связь позволяет произвести отладку взаимодействия между элементами и добиться их оптимального взаимодействия для достижения предельно возможного положительного результата в работе системы, состоящей из этих элементов.
Принцип доминанты. Это означает, что нервный путь или нервный центр наиболее активный получает преимущество по отношению к другим путям или центрам и начинает доминировать, господствовать над ними. Он тормозит их деятельность и перехватывает их возбуждение, чтобы усилить своё.
Принцип иерархии (соподчинения). Это означает, что одни элементы (нейроны и/или центры) подчиняются влиянию других элементов. Как правило, вышерасположенные центры подчиняют себе нижерасположенные центры.
Принцип пластичности. Это означает, что нервный центр перестраивает свою деятельность, приспосабливаясь к наилучшему выполнению своей функции для достижения общего конечного системного результата. Пластичность является важнейшей отличительной особенностью биосистем по сравнению с техническими системами.
Низшие нервные центры
Низшие нервные центры играют важную роль в работе любой сенсорной системы. Они являются одним из необходимых элементов сенсорной системы, по которому это понятие отличается от понятия "анализатор". Нервные центры не просто переключают возбуждение с одних нейронов на другие с помощью вставочных нейронов, т.е. выполняют "релейную" функцию, как это считалось ранее. Важно понять, что нервные центры занимаются трансформацией поступающего в них возбуждения, т.е. его преобразованием, или перекодированием. В результате этой трансформации входящее афферентное возбуждение превращается в выходящее эфферентное , отличающееся от входящего.
Работа (функции) низших нервных центров
1. Трансформация возбуждения , т.е.преобразование входящего потока сенсорного возбуждения в новый поток - выходящий. Выходящий поток может сильно отличаться от входящего, например, в том случае, если он должен управлять мышцами, а не строить нервную модель раздражения в виде сенсорного образа.
Виды трансформации возбуждения в нервном центре
1. Усиление.
2. Ослабление.
3. Блокировка.
4. Изменение паттерна (узора, характера).
5. Контрастирование границ в пространстве. Обычно оно достигается с помощью латерального (бокового) торможения. Латеральное торможение усиливает возбуждение по контуру раздражителя и рецептивного поля и ослабляет возбуждение в центральной области рецептивного поля.
6. Контрастирование границ во времени. Происходит за счёт преобразования тонического (постоянного) возбуждения в кратковременное фазическое. Таким способом отмечаются начало и конец действия раздражителя.
2. Распределение входящих потоков сенсорного возбуждения по выходящим потокам, которые направляются в различные нервные структуры. Эта функция нервного центра наглядно показана с помощью нашей схемы "Пути сенсорного возбуждения".
Виды распределения возбуждения в нервном центре
1. Конвергенция (схождение).
2. Дивергенция (расхождение).
3. Окклюзия (запирание).
4. Иррадиация (распространение).
3. Детекция . С помощью детекции выделяются раздражители с определёнными характеристиками за счёт срабатывания специальных нейронов-детекторов с соответствующими рецептивными полями. На любые другие раздражители, неадекватные для них, такие нейроны-детекторы не срабатывают, т.к. просто не возбуждаются такими стимулами.
Под нервным центром понимают сложный комплекс (ансамбль) нейронов, совокупно участвующих в регуляции какой-то физиологической функции или осуществлении определенного рефлекторного акта. Существует "понимание" нервных центров в узком и широком смысле этого понятия. Под нервными центрами в узком смысле понимают лишь те структуры центральной нервной системы, которые являются жизненно необходимыми для осуществления или регуляции определенного физиологического процесса, тогда как нервный центр в широком смысле этого понятия представляет собой совокупность всех компонентов центральной нервной системы, имеющих отношение к осуществлению и регуляции определенного физиологического процесса. Так, например, жизненно важный дыхательный центр, обеспечивающий ритмическую смену вдоха выдохом, находится в продолговатом мозге, но в то же время сами мотонейроны, иннервирующие дыхательную мускулатуру, заложены в спинном мозге. Кроме того, нервные клетки, имеющие отношение к регуляции внешнего дыхания, находятся в мосте, гипоталамусе (части промежуточного мозга) и коре больших полушарий. В связи с отмеченным под дыхательным центром в узком смысле этого слова понимают лишь структуры продолговатого мозга, имеющие отношение к периодической смене вдоха выдохом, тогда как в широком смысле этого понятия – совокупность всех нервных элементов центральной нервной системы, имеющих отношение к регуляции дыхания.
Для всех нервных центров характерны следующие свойства:
1. Одностороннее проведение возбуждения (возбуждение проводится всегда в одном направлении от афферентного нейрона к эфферентному, а затем к структурам исполнительных органов). Достигается благодаря существованию синапсов химического типа между чувствительными, вставочными и эфферентными нейронами.
2. Замедленное проведение возбуждения через нервные центры. Обусловлено передачей информации от одних нейронов центральной нервной системы к другим, имеющим отношение к регуляции какой-то физиологической функции, преимущественно через химические синапсы, для которых характерна синаптическая задержка (0,3-0,5 мс при проведении информации через один синапс). Соответственно, чем больше синаптических контактов встречается на пути нервного импульса от чувствительного нейрона к эфферентному, тем длительнее будет общая задержка проведения через соответствующий нервный центр.
3. Зависимость рефлекторного ответа от силы и длительности раздражения рецептивного поля рефлекса. Существование такой связи обусловлено тем, что, чем сильнее действующий на рецептивное поле какого-то рефлекторного акта раздражитель и чем больше длительность его действия, тем соответственно больше будет количество элементов нервного центра, вовлекаемых в реакцию, и большим окажется количество структур иннервируемых рабочих органов, участвующих в рефлекторном ответе, а, следовательно, и выше будет амплитуда рефлекторного ответа.
4. Суммация возбуждений в нервном центре. Это свойство проявляется следующим образом: раздражение каких-то отдельных рецепторов рецептивного поля нервного центра слабыми раздражителями не вызывает возникновения рефлекторного ответа, тогда как последовательное с определенной частотой раздражение этого же рецептора или одновременное воздействие на несколько соседних рецепторных элементов теми же слабыми стимулами, приводит к возникновению рефлекторного ответа. В основе описанного феномена лежат явления суммации возбуждений, поступающих к одному и тому же центральному нейрону либо через какой-то один из его синаптических входов с определенной частотой (временная суммация), либо одновременно через несколько его синаптических входов (пространственная суммация).
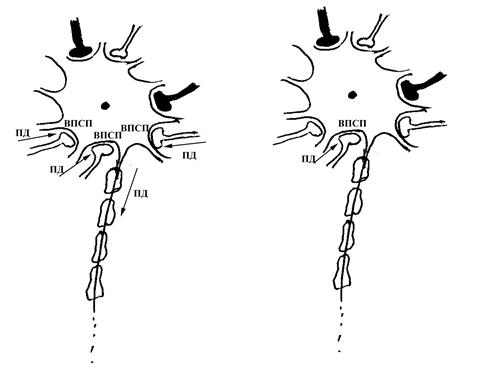
Рис. 15. Схема пространственной суммации информации, поступающей к мембране тела нейрона по различным его синаптическим входам. Обратите внимание на то, что на рисунке схематически изображен нейрон, на теле которого расположено большое количество синапсов (незатушеванные – возбуждающие, затушеванные – тормозные). На рисунке А показан такой вариант, когда к пресинаптическим полюсам сразу нескольких синапсов (в данном случае трех возбуждающих синапсов) одновременно приходит возбуждение, которое инициирует возникновение в постсинаптической мембране этих синапсов (являющейся частью мембраны тела рассматриваемого нейрона) возбуждающих постсинаптических потенциалов (ВПСП), суммирующихся в пространстве. Суммарный возбуждающий постсинаптический потенциал, возникающий в результате такой суммации, является достаточным для инициации потенциала действия (ПД) в области аксонного холмика данного нейрона.
На рисунке Б показан вариант, когда возбуждение приходит только к одному из пресинаптических полюсов возбуждающих синапсов, поэтому ВПСП возникает только в области постсинаптической мембраны данного синапса, и сам по себе оказывается недостаточным для инициации ПД в области аксонного холмика нейрона.
5. Трансформация ритма возбуждений. Заключается в том, что количество нервных импульсов, поступающих к какому-либо центральному нейрону (например, по чувствительным нервным волокнам), как правило, всегда отлично от такового, возникающего в ответ в этом нейроне. Так, частота импульсации, проводимой по чувствительным нервным волокнам, как правило, отлична от той, которая возникает во вставочных и эфферентных нервных волокнах. Отмеченное обстоятельство обусловлено разной лабильностью и электрофизиологическими свойствами (например, неодинаковой продолжительностью следовых потенциалов, сопровождающих пик потенциала действия) различных нервных элементов нервного центра
6. Рефлекторное последействие. Проявляется в том, что собственно рефлекторный ответ на действие раздражителя прекращается не мгновенно после прекращения раздражения соответствующего рецептивного поля рефлекса, а спустя какое-то время, что свидетельствует в пользу некоторой инерции возбуждения в нервном центре. Такая инерция возбуждения в нервном центре может быть следствием, как минимум, двух обстоятельств. Во-первых, возбуждение вставочного или эфферентного нейрона, возникающее в результате передачи информации от чувствительного нейрона, не исчезает сразу же после прекращения возбуждения чувствительного нейрона, а может сохраняться некоторое время благодаря выраженной следовой деполяризации в нейронах нервного центра после каждого потенциала действия, вызывающей возникновение новых импульсных ответов. Во-вторых, инерция возбуждения в нервных центрах может быть обусловлена способностью возбуждения циркулировать по замкнутым нейронным цепям нервного центра.
 |
Рис. 16. Схема циркуляции возбуждения по замкнутым нервным цепям в нервных центрах (по Бабскому Е.Б, 1972).
7. Утомление нервных центров. Состоит в снижении работоспособности нервных центров после длительного периода постоянной их активности и, как следствие, временном ослаблении или полном прекращении регуляции какой-то физиологической функции. Более высокая утомляемость нервных центров, по сравнению с нервными клетками, обусловлена наличием синаптических контактов химического типа между различными нейронными элементами в пределах нервного центра.
8. Тонус нервных центров, проявляющийся в постоянном их возбуждении. Возникает и поддерживается в большинстве нервных центров по причине постоянного поступления информации по афферентным нейронам от исполнительных органов рефлекторных актов (т.е. наличию постоянной афферентации от исполнительных органов в нервный центр).
9. Избирательная повышенная чувствительность нервных центров к определенным веществам, нарушающим их нормальную работу(т.н. нервным ядам). Предполагают, что нервные яды нарушают нормальное протекание обмена веществ в нервных центрах, действуя на определенные ферментативные системы их нейронов. Так, например, рвотный центр обладает высокой чувствительностью к апоморфину: это вещество вызывает стойкое возбуждение рвотного центра и связанную с этим длительную рвоту.
10. Существование постоянной обратной связи между нервным центром и исполнительными органами.Достигается благодаря постоянному поступлению по чувствительным нервным волокнам к нервному центру информации о функциональном состоянии рабочих (исполнительных) их органов, что, с одной стороны, обеспечивает поддержание постоянного тонуса (степени возбуждения нервного центра), а с другой – очень тонкую регуляцию деятельности исполнительных органов, поскольку эфферентные нервные посылки к органам постоянно модифицируются в зависимости от изменения функционального состояния исполнительных органов в момент осуществления определенного рефлекторного акта и соответственно поступающей к нервным центрам афферентной информации о состоянии регулируемых органов.
11. Пластичность нервных центров, проявляющаяся в компенсаторной их приспособляемости к повреждению и возможности функциональной перестройки. Так, частичное повреждение нервного центра вначале сопровождается возникновением определенных расстройств в регулируемой физиологической функции, которые со временем полностью или частично проходят по причине компенсаторных изменений в оставшихся нормальными структурах нервного центра, восполняющих функцию поврежденных структур. Функциональная перестройка нервных центров возникает также в случае искусственного, вызванного хирургическим вмешательством, изменения связи нервных центров с исполнительными органами. Так, например, перерезка каких-то нервов, иннервирующих различные органы, и последующее перекрестное их сшивание, в результате чего нервы начинают иннервировать другие внутренние органы, со временем сопровождается тем, что нервные центры начинают нормально регулировать деятельность других органов, с которыми оказались связанными (т.е. нервные центры приобретают новые, ранее им не свойственные, функции). Так, например, при параличе лицевого нерва иногда производят операцию по сшиванию периферического конца этого нерва (собственно вступающего в контакт с мимической мускулатурой лица) с какими-либо другими нормальными черепно-мозговыми нервами, и через некоторое время функция неработающих по причине паралича лицевого нерва мимических мышц лица восстанавливается.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
- Нервный импульс
- Нервы
НЕРВНЫЙ ЦЕНТР — совокупность нервных клеток (нейронов), необходимая для регуляции деятельности других нервных центров или исполнительных органов. Простейший нервный центр состоит из нескольких нейронов, образующих узел (ганглий). У высших животных и человека… … Большой Энциклопедический словарь
НЕРВНЫЙ ЦЕНТР — совокупность нейронов, б. или м. строго локализованная в нервной системе и участвующая в осуществлении рефлекса, в регуляции той или иной функции организма или одной из сторон этой функции. В простейших случаях Н. ц. состоит из неск. нейронов,… … Биологический энциклопедический словарь
нервный центр — совокупность нервных клеток (нейронов), необходимая для регуляции деятельности других нервных центров или исполнительных органов. Простейший нервный центр состоит из нескольких нейронов, образующих узел (ганглий). У высших животных и человека… … Энциклопедический словарь
нервный центр — nervinis centras statusas T sritis Kūno kultūra ir sportas apibrėžtis Grupė nervų ląstelių, reguliuojančių arba dalyvaujančių vykdant kurią nors organizmo funkciją (pvz., kvėpavimo, regėjimo). atitikmenys: angl. nerve centre vok. Nervenzentrum, n … Sporto terminų žodynas
НЕРВНЫЙ ЦЕНТР — совокупность нерв. клеток (нейронов), необходимая для регуляции деятельности др. Н. ц. или исполнит. органов. Простейший Н. ц. состоит из неск. нейронов, образующих узел (ганглий). У высших животных и человека Н.ц. включает тысячи и даже миллионы … Естествознание. Энциклопедический словарь
НЕРВНЫЙ ЦЕНТР — совокупность нейронов, более или менее локализованная в нервной системе и участвующая в осуществлении рефлекса, в регуляции той или функции организма или одной из ее сторон. Представления о Н. ц. лежат в основе представления о локализации функций … Психомоторика: cловарь-справочник
Нервный центр — более или менее локализованная совокупность нервных клеток, осуществляющая регуляцию какой либо функции организма. Нервные образования, связанные с регуляцией одной функции, могут находиться в различных отделах ЦНС. Н. ц. состоит из афферентных,… … Словарь дрессировщика
Нервный центр — – 1. вообще – любая область (локальная зона) центральной нервной системы, которая выполняет функции интегрирования и координирования нервной информации; 2. специальное значение – местоположение нервной ткани, где афферентная (входящая в мозг)… … Энциклопедический словарь по психологии и педагогике
НЕРВНЫЙ ЦЕНТР — 1. Вообще любая точка нервной системы, которая выполняет функции интегрирования и координирования нервной информации. 2. Специальное значение – местоположение нервной ткани, где афферентная информация совершает переход к эфферентной информации … Толковый словарь по психологии
Нервный центр — – совокупность нервных образований в ЦНС разных отделов, осуществляющих регуляцию специализированной функции организма или выполнение рефлекса; нервных центров в организме столько же, сколько и рефлекторных актов; основные свойства:… … Словарь терминов по физиологии сельскохозяйственных животных
Рефлексом (от лат. reflecto - отражение) называется ответная реакция организма, возникающая на раздражение рецепторов и осуществляемая с участием ЦНС.
В основе рефлекторной теории Сеченова лежат четыре принципа:
1. Структурности (структурной основой рефлекса является рефлекторная дуга)
2. Детерминизма (принцип причинно-следственных отношений). Ни одна ответная реакция организма не бывает без причины.
3. Анализа и синтеза (любое воздействие на организм сначала анализируется, затем обобщается).
4. Обратной связи (отображающий точность реакций и адаптацию), особенно развитый в трудах академика П.К. Анохина.
Итак, структурной основой рефлекса является рефлекторная дуга.
Функционально она состоит из: афферентного, центрального и эфферентного звеньев, связанных между собой синаптическими соединениями.
Морфологически она состоит из:
- рецепторных образований, назначение которых заключается в трансформации энергии внешних раздражений (информации) в энергию нервного импульса; В слизистой оболочке рта сосредоточено множество рецепторов, воспринимающих раздражения внешней и внутренней среды. Раздражение рецепторов полости рта вызывает ответные реакции в сердце, органах пищеварения, эндокринных железах и др., изменяя их деятельность.
- афферентного (чувствительного) нейрона, проводящего нервный импульс в нервный центр;
- интернейрона (вставочного) нейрона или нервного центра, представляющего собой центральную часть рефлекторной дуги;
- эфферентного (двигательного) нейрона, проводящего нервный импульс до эффектора;
- эффектора (рабочего органа), осуществляющего соответствующую деятельность. Полость рта и лицевая область могут являются эффекторным полем рефлекторного влияния с внутренних органов.
На основании принципа обратной связи необходимо дополнить рефлекторную дугу еще одним звеном – петлей обратной связи, которая устанавливает связь между исполнительной структурой и центром рефлекторной дуги или рецепторными структурами (морфологически – коллатераль аксона двигательного нейрона или другой нейрон?). Такая структура рефлекторной дуги превращает открытую рефлекторную дугу в закрытую, т.е. в самонастраивающийся нервный контур регуляции физиологической функции, оптимизируя поведение организма.
В зависимости от структуры рефлекторной дуги (т.е. количества нейронов или синаптических связей) различают моносинаптические (двухнейронные) рефлексы, например, коленный рефлекс, и полисинаптические (три и более нейронов) рефлексы.
На основании рассмотренного морфологического строения рефлекторной дуги мы уже можем заключить, что структурно-функциональной единицей нервной системы является нейрон.Действительно, в основе представления о структуре и функции ЦНС лежит нейронная теория, которая получила признание лишь на рубеже 20 века. Нервная система состоит из двух типов клеток: нейронов и клеток глии. Однако именно нейроны обеспечивают все многообразие процессов, связанных с передачей и обработкой информации.
Структура и функция нейронов и синапсов.
В каждом нейроне можно выделить 2 основных элемента:
2. отростки – дендриты и аксон с пресинаптическими окончаниями.
Тело нейрона содержит различные внутриклеточные органеллы, необходимые для обеспечения жизнедеятельности всей клетки: ядро, рибосомы, эндоплазматический ретикулум, аппарат Гольджи, митохондрии. Мембрана покрыта синапсами. Функции – синтез макромолекул, восприятие, интеграция сигналов, поступающих от других нейронов.
Дендриты – самая вариабельная часть нейрона. Сильно ветвятся, образуют различные выросты – шипики, значительно увеличивая поверхность тела нейрона. Это создает условия для размещения на дендритах большого числа синапсов. Функция – восприятие информации, передача информации к телу клетки.
Аксон – менее вариабельная часть нейрона. Начальный участок аксона называется аксональным холмиком, обладает низким порогом возбуждения по сравнению с другими участками мембраны нейрона. Аксон может образовывать коллатерали. Заканчивается аксон пресинаптическими окончаниями. Функция – проведение нервного импульса от тела клетки к другим клеткам или исполнительным органам; транспортная – внутриклеточный антероградный (синтезированные в соме активные вещества, митохондрии) и ретроградный транспорт (вирус герпеса, полиемиелита).
1. По количеству отростков –уни -, би-, мультиполярные.
2. По функции: сенсорные (чувствительные, афферентные), интернейроны (вставочные), моторные (двигательные, эфферентные).
Функции нейронов: Функции глии: 1). Защитная, опорная,
1. Интегративная; изолирующая.
2. Координирующая Трофическая ф. не доказана, но
3. Трофическая выявлена роль в процессах памяти.
Итак, уникальными особенностями нейрона являются способность генерировать электрические разряды и передавать информацию с помощью специализированных межклеточных соединений – синапсов. Являясь главным механизмом связи между нейронами, синапсы во многом обеспечивают все многообразие функций мозга.
Понятие синапсбыло введено в физиологию англ. физиологом Ч. Шеррингтоном в 1897 году для обозначения функционального контакта между нейронами. Синапсы различаются по:
- механизму действия (электрический, химический, смешанный);
- локализации на поверхности клетки (аксосоматические, аксодендрические, реже - аксоаксонные, дендродендрические);
- функции (возбуждающие или тормозящие);
- способностью к модуляции?[K1]
Синапс представляет собой сложное функциональное образование и состоит из:
- постсинаптической мембраны (субсинаптическая мембрана).
Мы разберем строение и работу наиболее часто встречающихся у человека химических синапсов.
Пресинаптическая мембрана, как мы уже говорили, это окончание аксона.
Синаптическая щель широкая и составляет в среднем 10-20 нм, подавляющая часть пресинаптического тока шунтируется низким сопротивлением щели. Поэтому в химических синапсах существует особый механизм, способный изменить мембранный потенциал постсинаптической мембраны и таким образом передать информацию от клетки к клетке. Таким механизмом является выделение пресинаптическим окончанием аксона особых химических веществ – нейротрансмиттеров (или медиаторов), которые, воздействуя на специфические рецепторы постсинаптической мембраны, способны изменять состояние ионных каналов постсинаптической мембраны.
В настоящее время выявлено более 15 нейротрансмиттеров (ацетилхолин, норадреналин, серотонин, глицин, гамма-аминомаслянная кислота и др.). Существует правило Дейла – каждый нейрон во всех своих пресинаптических окончаниях выделяет один и тот же медиатор, поэтому нейроны или синапсы иногда обозначают по типу медиатора (холинергические, адренергические, серотонинергические и др.). Однако как из каждого правила существуют исключения, так и в данном случае правило Дейла не абсолютно. Экспериментально обнаружена возможность выделения одним и тем же пресинаптическим окончанием множества других нейротрансмиттеров.
Постсинаптическая мембрана (а именно субсинаптическая) имеет специализированные рецепторы и хемочувствительные ионные каналы.
Разберем механизм синаптической передачи.
В пресинаптических окончаниях аксона депонируется определенное количество нейротрансмиттеров (медиаторов) в синаптических пузырьках или везикулах, каждая из которых содержит один квант медиатора, состоящий из нескольких тысяч молекул.
Деполяризация пресинаптической мембраны под действием распространяющегося импульса увеличивает проницаемость для Са 2+ . При наличии Са 2+ везикула, подойдя к внутренней поверхности мембраны пресинаптического окончания, сливается с пресинаптической мембраной. В результате происходит выброс кванта медиатора в синаптическую щель. Таким образом, выход кванта нейротрансмиттера (медиатора) из везикул представляет собой процесс экзоцитоза. После спадения везикулы окружающая его мембрана включается в мембрану пресинаптического окончания. В дальнейшем в результате эндоцитоза небольшие участки пресинаптической мембраны впячиваются внутрь, вновь образуя везикулы, которые снова способны включать нейротрансмиттер (медиатор) и вступать в цикл его освобождения.
Участие Са 2+ определяет ряд важных особенностей работы химических синапсов. Прежде всего, время, необходимое для вхождения Са 2+ внутрь клетки, определяет синаптическую задержку проведения, составляющую 0,2-0,5 мс. Кроме того, накопление Са 2+ внутри пресинаптического окончания в результате предшествующего поступления улучшает эффективность работы химического синапса, т.е. вызывает высвобождение большего числа квантов медиатора, что проявляется увеличением амплитуды постсинаптических потенциалов.
В возбуждающем синапсе происходит увеличение проницаемости для Nа + или Са 2+ , в результате чего происходит деполяризация мембраны и возникает локальный ответ – ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ (ВПСП).
В тормозном синапсе происходит увеличение проницаемости для К + , в результате чего происходит гиперполяризация мембраны и возникает локальный ответ - ТОРМОЗНОЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ (ТПСП).
Характер действия медиатора определяется свойствами субсинаптической мембраны.
Следует еще раз подчеркнуть, что на субсинаптической мембране возникает лишь локальный ответ. Возникновение потенциала действия (ПД) за счет суммации локальных ответов происходит в специальной низкопороговой области (зона аксонного холмика), откуда ПД распространяется по аксону и на мембрану соседних участков клетки.
Неиспользованный медиатор или его фрагменты всасываются обратно в пресинаптическую часть синапса.
Нейрон имеет множественные синаптические контакты с другими нейронами. Два принципа: дивергенции и конвергенции.
Дивергенция – это способность нервной клетки устанавливать многочисленные синаптические связи с различными нервными клетками. В результате афферентная информация поступает одновременно к разным участкам ЦНС, один нейрон может участвовать в нескольких различных реакциях, передавать возбуждение значительному числу других нейронов, обеспечивая широкую иррадиацию возбудительного процесса в центральных нервных образованиях (кашлевой рефлекс).
Конвергенция – это схождение различных импульсных потоков от нескольких нервных клеток к одному нейрону. Интегративная функция. Мотонейрон – общий конечный путь двигательной системы (англ. физиол. Шеррингтон)
Понятие о нервном центре, его свойства.
Если нейрон можно рассматривать как структурную единицу нервной системы, то физиологической системной единицей является нервный центр.
Нервный центр – это динамическая совокупность нейронов, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт.
Функциональный нервный центр может быть локализован в разных анатомических структурах. Например, дыхательный центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, в коре головного мозга.
Нервные центры имеют ряд общих свойств, определяемых наличием синаптических образований и структурой нейронных сетей, образующих эти центры:
1. Низкая лабильность. Обусловлена скоростью развития синаптической передачи импульса в химическом синапсе. 50-100 имп/с.
2. Высокая утомляемость. Утомление – временное снижение работоспособности в результате проведенной работы, которое исчезает после отдыха. Причины: а). истощение и несвоевременный синтез медиатора; б). адаптация постсинаптического рецептора к медиатору; в). инактивация рецепторов в результате длительной деполяризации постсинаптической мембраны.
3. Высокая чувствительность к недостатку кислорода. Мозг в 22 раза больше потребляет кислорода, чем мышечная ткань. Необратимые изменения наступают в коре через 4-5 мин, в стволовых клетках – через 15-20 мин.
4. Высокая чувствительность к фармакологическим веществам (блокаторы нервно-мышечной передачи, психомиметические средства), ядам.
Токсин Cl. вotulinum – блокада высвобождения возбуждающего медиатора.
Столбнячный токсин – блокада высвобождения тормозного медиатора.
Удаление Са 2+ или замещение на Мg 2+ - блокада высвобождения медиатора
Гемихолиний – нарушение синтеза медиатора.
Бунгаротоксин – необратимое связывание с АХ-рецепторами
Яд кураре – конкурентное связывание с АХ-рецепторами.
Стрихнин – конкурентное связывание с глицин-рецепторами.
Бикулин, пикротоксин (судорожные яды), пенициллин – конкурентное связывание с ГАМК-рецепторами.
Фосфоорганические соединения – угнетение холинэстеразы и вследствие этого продолжительная субсинаптическая деполяризация и инактивация рецепторов.
Релаксанты (сукцинилхолин) – аналоги АХ, но не расщепляющиеся АХЭ, продолжительная деполяризация субсинаптической мембраны и инактивация рецепторов.
Алкоголизм, привыкание, наркомания.
5. Пластичность. Основа: изменение структуры и функции синапсов. Поэтому синапсы участвуют в таких функциях ЦНС как научение и память, компенсации функции при нарушении за счет формирования новых нейронных связей; синтеза специфических белков.
Особенности проведения возбуждения по нервному центру определяют закономерности проведения возбуждения по рефлекторной дуге:
1. Одностороннее проведение. Обусловлено особенностями проведения возбуждения по синапсу. Медиаторы выделяются в пресинаптическом окончании, рецепторы находятся в постсинаптической мембране.
2. Замедленное проведение, обусловленное синаптической задержкой в центральной части рефлекторной дуги. Составляет 0.2-0.5 мс и определяет время рефлекса (от начала раздражения до начала ответной реакции).
3. Суммация подпороговых раздражений. Например, чихательный рефлекс. И.М. Сеченов открыл, изучил. Два типа:
1. Суммация последовательная (временная);
2. Суммация пространственная (одновременная)
П/п раздражения наносятся одновременно на несколько точек рецептивного поля, в результате конвергенции происходит суммация локальных ответов.
4. Трансформация ритма возбуждения.
При ритмическом возбуждении нервный центр перестраивает ритм как понижая, так и повышая частоту следования импульсов. Понижение связано с низкой лабильностью синапса (максимально – 100 имп/с). Повышение обусловлено: - возникновением повторных разрядов на фоне длительной следовой деполяризации; - наличием полисинаптических нервных цепей; - циркуляцией импульсов в замкнутых нейронных цепях.
5. Рефлекторное последействие – продолжение рефлекторной реакции после окончания действия раздражителя. Механизмы те же, что и механизмы повышающей трансформации.
6. Ритмическая активация синапса часто сопровождается значительным увеличением амплитуды синаптических потенциалов. Посттетаническая потенциация – это усиление рефлекторного ответа после тетанических раздражений. Длительность посттетанической потенциации может составлять от нескольких минут до нескольких часов. С функциональной точки зрения посттетаническая потенциация представляет собой процесс облегчения в ЦНС, связанный с приобретением опыта, т.е. процесс научения, памяти.
1). Ритмическая активация мембраны пресинаптического окончания аксона ведет к увеличению амплитуды ПД, что вызовет высвобождение большего количества медиатора в синаптическую щель.
2). Ритмическая активация сопровождается увеличением запаса доступного медиатора, готового к выделению.
3). Ритмическая активация приводит к вхождению и накоплению Са 2+ в пресинаптическом окончании(мы уже с вами об этом упоминали при рассмотрении работы синапса), что увеличивает высвобождение медиатора в синаптическую щель. Это самый важный фактор для посттетанической потенциации.
Читайте также:
