
Что такое чувствительные нервные окончания
Все нервные волокна без исключения имеют свой концевой аппарат, который носит название - нервные окончания.
Классификация нервных окончаний
В зависимости от функционального назначения нервные окончания подразделяются на три основные группы:
Рецепторные нервные окончания.
Рецепторные нервные окончания локализованы по всему организму и воспринимают различные раздражения как из внешней среды, так и от внутренних органов.
Экстерорецепторы воспринимают раздражение из окружающий среды. К экстерорецепторам (внешним) относятся: слуховые, зрительные, обонятельные, вкусовые и осязательные рецепторы.
Интерорецепторы воспринимают раздражение от собственных органов или тканей организма. К интерорецепторам(внутренним) относятся: висцеро-рецепторы (сигнализирующие о состоянии внутренних органов) и проприорецепторы (или рецепторы опорно-двигательного аппарата).
Разновидностью интерорецепторов являются проприорецепторы - чувствительные нервные окончания в мышцах, сухожилиях, которые принимают участие в регуляции движений и положения тела в пространстве.
В зависимости от специфичности раздражителя, воспринимаемого данным видом рецептора все чувствительные окончания делятся на механорецепторы, барорецепторы, хеморецепторы, терморецепторы и др.
По особенностям строения чувствительные окончания подразделяют на свободные нервные окончания, т.е. состоящие только из конечных ветвлений осевого цилиндра, и несвободные, содержащие в своем составе все компоненты нервного волокна, а именно ветвления осевого цилиндра и клетки глии. Несвободные окончания, кроме того, могут быть покрыты соединительнотканной капсулой, и тогда они называются инкапсулированными. Несвободные нервные окончания, не имеющие соединительнотканной капсулы, называются неинкапсулированными.
Свободные нервные окончания обычно воспринимают холод, тепло и боль. Такие окончания характерны для эпителия. В этом случае миелиновые нервные волокна подходят к эпителиальному пласту, теряют миелин, а осевые цилиндры проникают в эпителий и распадаются там между клетками на тонкие терминальные ветви.
Несвободные нервные окончания могут быть покрыты соединительнотканной капсулой и тогда они носят название инкапсулированных.
Несвободные нервные окончания, которые не имеют соединительнотканной капсулы, называются неинкапсулированными.
Инкапсулированные рецепторы соединительной ткани при всем их разнообразии всегда состоят из разветвленного осевого цилиндра и глиальных клеток, которые снаружи покрыты соединительнотканной капсулой. К инкапсулированным нервным окончаниям относятся также рецепторы мышц и сухожилий: это нервно-мышечные веретена и нервно-сухожильные веретена.
Нервно-мышечные веретена являются сенсорными органами в скелетных мышцах, которые функционируют как рецептор на растяжение.
Нервно-сухожильные веретена обычно располагаются в месте соединения мышцы с сухожилием.
Эффекторные нервные окончания: разновидности, строение. Функции.
Все нервные волокна без исключения имеют свой концевой аппарат, который носит название - нервные окончания.
Классификация нервных окончаний
В зависимости от функционального назначения нервные окончания подразделяются на три основные группы:
Среди эффекторных нервных окончаний различают двигательные и секреторные.
Двигательные нервные окончания — это концевые аппараты аксонов двигательных клеток соматической или вегетативной нервной системы. При их участии нервный импульс передается на ткани рабочих органов.
Двигательные окончания в поперечнополосатых мышцах называются нервно-мышечными окончаниями. Они представляют собой окончания аксонов клеток двигательных ядер передних рогов спинного мозга или моторных ядер головного мозга. Нервно-мышечное окончание состоит из концевого ветвления осевого цилиндра нервного волокна и специализированного участка мышечного волокна. Миелиновое нервное волокно, подойдя к мышечному волокну, теряет миелиновый слой и погружается в него, вовлекая за собой его плазмолемму и базальную мембрану. Мембрана мышечного волокна образует многочисленные складки, формирующие вторичные синаптические щели эффекторного окончания. В области окончания мышечное волокно не имеет типичной поперечной исчерченности и характеризуется обилием митохондрий, скоплением круглых или слегка овальных ядер.
Терминальные ветви нервного волокна в нервно-мышечном соединении характеризуются обилием митохондрий и многочисленными пресинаптическими пузырьками, содержащими характерный для этого вида окончаний медиатор — ацетилхолин. При возбуждении ацетилхолин поступает через пресинаптическую мембрану в синаптическую щель на холинорецепторы постсинаптической (мышечной) мембраны, вызывая ее возбуждение (волну деполяризации).
Постсинаптическая мембрана моторного нервного окончания содержит фермент ацетилхолинэстеразу, разрушающий медиатор и ограничивающий этим срок его действия.
Сходное строение имеют секреторные нервные окончания (нейрожелезистые). Они представляют собой концевые утолщения терминали или утолщения по ходу нервного волокна, содержащие пресинаптические пузырьки, главным образом холинергические.
Дата добавления: 2018-06-01 ; просмотров: 1118 ;
Нервные окончания — это концевые аппараты нервных волокон, различные по функциональному значению. Различают три вида нервных окончаний: межнейронные синапсы (контакты), обеспечивающие взаимосвязь между различными типами неронов; чувствительные или афферентные (от греч. affero — приносить) окончания, воспринимающие раздражения из окружающей среды; двигательные или эфферентные (от греч. effero — выносить) окончания, передающие сигналы от нейрона к исполнительному органу (мышце, железе).
Межнейронный синапс — разновидность специализированного контакта двух нейронов, обеспечивающего одностороннее проведение возбуждения. Термин синапс (от греч. synapses — соединение, связь) введен в 1897 г. австралийским ученым Ч. Шеррингтоном, обозначает структурно-функциональный контакт нейронов, информация (сигналы) от одной клетки к другой передается с помощью химического посредника — медиатора. В синапсе различают преси- наптическую мембрану (полюс) — концевой отдел аксона, для которого характерно скопление митохондрий и медиатора (посредника), заключенного в синаптические пузырьки. Постсинаптическая мембрана (полюс) синапса образована отростком второго нейрона.
Между пресинаптической и постсинаптической мембранами имеется небольшой просвет — синаптическое пространство, или синаптическая щель, в которую поступает медиатор.
Чувствительные, или афферентные, нервные окончания рецепторы дендритов чувствительных нейронов обеспечивают восприятие раздражения и преобразование его в нервный импульс.
В соответствии с локализацией и функцией различают две группы рецепторов: экстерорецепторы и интерорецепторы. Экстерорецеп- торы воспринимают сигналы из внешней среды, например, нервные окончания органов слуха, равновесия, зрения, вкуса, осязания. Интерорецепторы воспринимают раздражения из внутренних органов непосредственно терминальными отделами отростков чувствительных нейронов. Интерорецепторы подразделяются на свободные, неинкапсулированные и инкапсулированные нервные окончания.
Свободные нервные окончания состоят из терминальных разветвлений дендрита чувствительного нейрона, проникающих между эпителиальными клетками, например локализуются непосредственно в эпидермисе. Кроме того, встречаются в соединительной ткани поверхностных и глубоких слоев дермы кожи. Свободные нервные окончания выполняют универсальные функции и способны воспринимать механические, химические, температурные изменения, болевые стимулы. Неинкапсулированные нервные окончания представлены ветвящимся осевым цилиндром нервного волокна и снабжены клетками нейроглии, встречаются в соединительнотканных прослойках различных органов, и выполняют функцию барорецепторов (от греч. baros — тяжесть; recipion — воспринимать). Бароцепторы — чувствительные нервные окончания в кровеносных сосудах, воспринимающие изменения кровяного давления и рефлекторно регулирующие его уровень. Инкапсулированные нервные окончания представлены ветвящимся осевым цилиндром нервного волокна, в составе окончания имеются элементы нейроглии, окруженные капсулой из рыхлой волокнистой соединительной ткани.
Нервный импульс генерируется в условиях смещения наружной капсулы по отношению к внутренней колбе, образованной оголенными ветвлениями осевых цилиндров терминального отдела нервного волокна чувствительного нейрона, вокруг которых располагаются ядра леммоцитов. Капсула, или наружная колба, состоит из большого числа пластинок, образованных фибробластами и спирально ориентированными пучками коллагеновых волокон (рис. 55).
Двигательные, или эфферентные нервные окончания выполняют функцию передачи нервного импульса на рабочий орган (мышцы, железы). Различают нервно-мышечные двигательные окончания поперечнополосатых мышц, дистантные двигательные нервные окончания гладких мышц и сердечной мышечной ткани, а также секреторные двигательные нервные окончания желез.
Двигательные окончания поперечнополосатых мышц, так называемые моторные бляшки, представляют собой комплекс взаимосвязанных структур нервной и мышечной тканей. Моторная бляшка — двигательный аппарат терминальной части аксонов нейронов спинного (головного) мозга — это пресинаптическая мембрана и специализированный участок мышечного волокна — постсинаптическая мембрана, между мембранами имеется синаптическая щель, в которую поступает медиатор (ацетилхолин).

Рис. 55 [2]. Чувствительное нервное окончание:
7 — клетки поджелудочной железы; 2 — нервное волокно; 3 — осевой цилиндр; 4 — наружная колба; 5 — внутренняя колба; б — оголенный осевой цилиндр
В заключение следует обратить внимание на интегрирующую роль нервной ткани в организме. Основной формой деятельности нервной ткани является рефлекс (от греч. reflexus — отражение) — реакция организма на раздражение. Действие рефлекторного механизма вегетативного отдела нервной системы начинается с реакции чувствительных нейронов спинномозговых или черепно-мозговых нервов. В отличие от рефлекторных дуг соматического отдела двигательный нейрон рефлекторной дуги вегетативного отдела идет не в составе серого вещества центральной нервной системы, а в одном из периферических ганглиев (рис. 57).
Впервые представления о закономерностях деятельности животных и человека были сформулированы И.М. Сеченовым (1853). Заслуга разработки экспериментальных методов изучения и подтверждения теоретических предпосылок И.М. Сеченова принадлежит И.П. Павлову. Высшая нервная деятельность по И.П. Павлову — это

Рис. 56 [2]. Двигательное нервное окончание:
7 — нервное волокно; 2 — разветвления осевого цилиндра; 3 — моторная бляшка; 4 — мышечные волокна

Рис. 57 [13]. Схема рефлекторной дуги:
- 7 — чувствительное нервное окончание в коже; 2 — чувствительный нейрон в спинномозговом ганглии; 3 — ассоциативный нейрон; 4 — двигательный нейрон;
- 5 — двигательное нервное окончание в мышцах
объединенная рефлекторная (условно- и безусловно-рефлекторная) функция коры больших полушарий и ближайшей подкорки головного мозга, обеспечивающая приспособление (поведение) животного к окружающей среде. Безусловные рефлексы являются врожденными и передаются по наследству, осуществляются готовой к моменту рождения рефлекторной дугой, постоянны и относительно устойчивы в течение всей жизни. Условные рефлексы приобретаются в процессе жизни. Был установлен принцип: кора головного мозга благодаря выработке условных рефлексов осуществляет связь организма с внешней средой. Безусловные рефлексы свойственны трем отделам ЦНС: подкорковым ядрам, мозговым стволам, спинному мозгу; для того чтобы вызвать безусловный рефлекс, достаточно раздражения определенного рецептивного поля (например, пищеварительный рефлекс возникает при раздражении вкусовых рецепторов). Условные рефлексы изменчивы, возникают, утрачиваются по мере надобности; вырабатываются на базе безусловных при целостности коры полушарий головного мозга, так как дуга условного рефлекса проходит через корковые центры. По И.П. Павлову деятельность нервной системы осуществляется посредством взаимосвязанных морфологически и функционально механизмов-анализаторов и механизмов-рефлексов. Сложная рефлекторная дуга в данном случае состоит из анализатора и замыкательного аппарата, который включается при участии вставочных нейронов и в результате деятельности анализатора.
Контрольные вопросы
- 1. Как классифицируются нейроны?
- 2. Дайте характеристику и опишите локализацию специализированных структур нейронов.
- 3. Каковы функциональные особенности нейрофибрилл?
- 4. Назовите основные признаки дендритов и аксона нейрона.
- 5. Каковы особенности структуры клеток нейроглии?
- 6. Как классифицируют клетки нейроглии?
- 7. Что такое нервное волокно и каков механизм его образования?
- 8. Чем различаются миелиновые и безмиелиновые волокна?
- 9. Какова классификация нервных окончаний?
- 10. Что такое синапсы?
Классификация
1. По месту восприятия раздражения:
1.1. Экстерорецепторы, воспринимающие раздражения из внешней среды;
1.2. Интерорецепторы – восприятие раздражений внутренних органов;
1.3. Проприорецепторы – восприятие информации от опорно-двигательного аппарата.
2. В зависимости от природы раздражения, воспринимаемого рецептором:
2.1. Механорецепторы – механические раздражители, перемещение частей тела;
2.2. Хеморецепторы – химические раздражители.
2.3. Терморецепторы – изменения температуры.
2.4. Ноцирецепторы – чувство боли.
3. По способу восприятия:
3.1. Контактные рецепторы.
3.2. Дистантные рецепторы (рецепторные клетки сетчатки глаза, органа слуха, обоняния).
4. Морфологическая классификация основана на особенностях их структурной организации.
4.1. Свободные нервные окончания состоят только из конечных ветвлений дендрита чувствительного нейрона. Они встречаются в эпителии (кожа и серозные оболочки) и в соединительной ткани, обеспечивая восприятие температурных, механических и болевых сигналов.
4.2. Несвободные нервные окончания – содержат все компоненты нервного волокна (осевой цилиндр и клетки глии). Они могут быть инкапсулированными (имеющими особую соединительнотканную капсулу) и неинкапсулированные.
Неинкапсулировпнные нервные окончания представлены обязательными дисками Меркеля и встречаются в дерме кожи и собственной пластинки слизистых оболочек.
Инкапсулированные нервные окончания имеют общий принцип строения: их основу составляют ветвления дендрита, которые окружены леммоцитами и покрыты соединительнотканной капсулой. К этому виду окончаний относят: пластинчатые тельца (Фатер-Пачини), обязательные тельца (Мейснера), тельца Руффини, колбы Краузе, генитальные тельца Догеля, нервно-мышечные и нервно-сухожильные веретена.
Пластинчатые тельца (Фатер-Пачини) – в соединительной ткани внутренних органов и кожи. Имеют вид округлых образований (1-5 мм), воспринимают давление и вибрацию.
Структурными компонентами являются:
1) внутренняя колба (луковица), образованная видозимененными леммоцитами, в которую проникают нервные волокна;
2) наружная колба – слоистая соединительнотканная капсула из фибропластов и коллагеновых волокон, образующих концентрические пластины, между которыми имеется жидкость.
При деформации пластин давление передается на нервные окончания и вызывает деполяризацию мембраны.
Осязательные тельца (Мейснера) – в сосочковом слое дермы, имеют эллипсоидную форму и небольшие размеры (50-140 мкм). Внутренняя колба состоит из глиальных клеток, лежащих перпендикулярно длинной оси тельца, между которыми располагаются веточки дендритов. Снаружи находится очень тонкая, переходящая в периневрит слоистая капсула – наружная колба. Незначительная деформация капсулы передается глиоцитам и далее на дендриты.
Тельца Руффини лежат в соединительнотканной части кожи и капсулах суставов: они воспринимают давление и имеют вид веретеновидных структур длиной 1-2 мм. Внутреннюю колбу образуют глиальные клетки между которыми располагаются терминали дендритов с расширениями на концах. Капсула хорошо выражена.
Колбы Краузе – мелкие (40-150 мкм) округлые тельца, являющиеся механорецепторами и Холодовыми рецепторами. Они лежат в дерме кожи, слизистой оболочке полости рта, подгортанника, в конъюнктиве глаза. Внутренняя колба образована плоскими глиоцитами между которыми тонкие ветви дендрита образуют сплетения в виде клубочка. Наружная капсула очень тонкая.
Генитальные тельца Догеля – находятся в особо чувствительных областях кожи (наружные половые органы, молочные железы). Они по строению похожи на колбы Краузе, но в отличие от них в тельце входит несколько отростков от нейроцитов, что вызывает сильную иррадиацию возбуждения.
Нервно-мышечные веретена – рецепторы растяжения поперечнополосатых мышц - нервные окончания, обладающие как чувствительной, так и двигательной иннерваций. Наружная капсула веретена окружает несколько тонких интрафузальных мышечных волокон. Волокна, лежащие за пределами капсулы, называются экстрафузальными. Интрафузальные мышечные волокна двух видов:
- волокна с ядерной сумкой – в расширенной центральной части которых содержатся скопления ядер (ядерная сумка);
- волокна с ядерной цепочкой – более тонкие, ядра лежат по всей длине волокна в виде цепочки в центральной части (ядерная цепь).
Чувствительные нервные волокна образуют кольцеспиральные окончания (отростки нервных клеток закручены вокруг центральной части волокна по спирали) на волокнах обоих типов и гроздьевидные окончания у краев волокон с ядерной цепочкой.
Двигательные нервные волокна образуют мелкие нервномышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус, регулируя длину волокон. Всё свободное пространство между мышечными волокнами заполнено жидкостью и ограничено тонкой капсулой. Изменение тонуса мышцы ведет к изменению давления жидкости и передается на дендриты. Кольцеспиральные окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, гроздьевидные окончания – только на изменение длины. Число веретен в мышце зависит от её функции и тем выше, чем более точными движениями она обладает.
Нервно-сухожильные веретена – рецепторы растяжения, располагаются в месте соединения мышцы с сухожилием, веретеновидные структуры длиной 0,5-1 мм. Каждое веретено имеет капсулу из фиброцитов, которая охватывает группу сухожильных пучков, оплетенных окончаниями нервных волокон. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.
Рефлекторная дуга – это цепь нейронов, связанных синапсами и обеспечивающих проведение нервного импульса к рабочему органу. Простая рефлекторная дуга состоит из двух нейронов – чувствительного и двигательного. В сложной рефлекторной дуге между ними включены вставочные, или ассоциативные нейроны. Возбуждение по рефлекторной дуге передается только в одном направлении.
| | | следующая лекция ==> | |
| Нервные окончания | | | Поверхность в пространстве |
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
− чувствительные нервные окончания,
− двигательные нервные окончания и
Чувствительные нервные окончания (рецепторы). Определения:
1. Рецептор – это специализированный нервный аппарат, воспринимающий действия раздражителей,
2. (по И.П. Павлову), рецепторы – это трансформаторы энергии раздражителей в нервные импульсы.
Физиологическая классификация рецепторов:
− механорецепторы – стимулируются механическими движениями органов или их частей,
− барорецепторы – воспринимают изменения давления,
− хеморецепторы – стимулируются различными химическими веществами,
− терморецепторы – воспринимают колебания температуры.
Морфологическая классификация чувствительных нервных окончаний. По строению их делят на: свободные и несвободные. Несвободные в свою очередь делят на: неинкапсулированные и инкапсулированные.
Свободные нервные окончания – это голые ветвления осевого цилиндра, т.е. они (ветвления) свободны от оболочек (миелиновой и шванновской) (Рис. 18). Отличаются большой чувствительностью. Их много в составе эпителия, особенно в составе разновидностей многослойного эпителия. Пример. Роговица глаза отличается высокой чувствительностью, из-за наличия между клетками эпителия большого количества свободных чувствительных нервных окончаний.
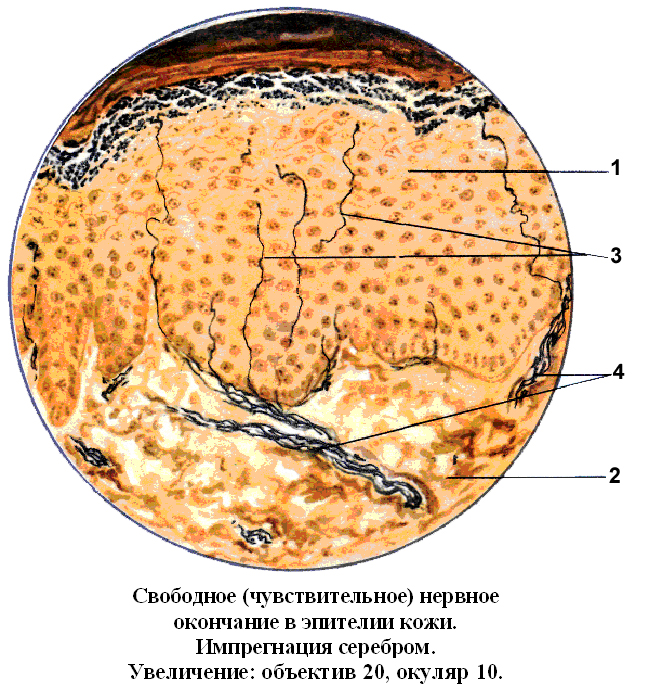
- Слой шиповатых клеток
- Рыхлая соединительная ткань
- Терминали
- Нервные волокна
Несвободные нервные окончания. В них концевые ветвления осевого цилиндра покрыты оболочками.
Несвободные неинкапсулированныенервные окончания – когда концевые ветвления осевого цилиндра покрыты оболочкой из клеток нейроглии. Таких рецепторов много в составе эпителия и соединительных тканей.
Несвободные инкапсулированные нервные окончания. Их разновидности: колбы Краузе, тельца Мейснера, тельца Фатер – Пачини. По строению они сходны. Рассмотрим их строение на примере телец Фатер – Пачини (Рис. 19). В средне части тельца располагается концевое ветвление осевого цилиндра. Оно окружено видоизмененными глиальными клетками, образующими внутреннюю капсулу тельце. Внутренняя капсула окружена наружной капсулой, состоящей из наслоенных друг на друга соединительнотканных пластинок. Тельца Фатер – Пачини воспринимают давление на орган, т.е. функционально механорецепторы.
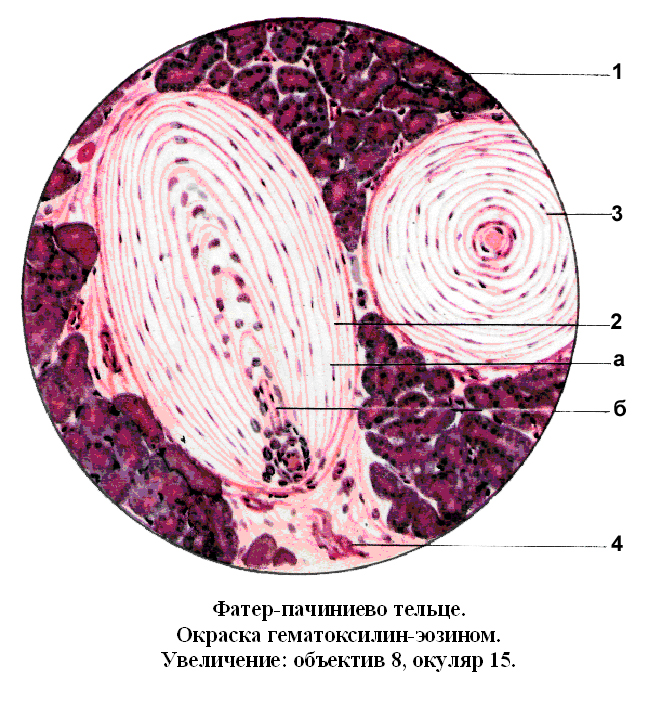
1.Секреторные отделы железы
2, 3. Пластинка соединительной ткани
б. Осевой цилиндр
4. Миелиновое нервное волокно.
Двигательные нервные окончания передают нервные импульсы на рабочие органы и ткани. В скелетных мышцах они называются нервно-мышечные окончания.
Синапс. Это место контакта двух нейронов. В зависимости от того, какие части двух нейронов контактируют, различают следующие виды синапсов:
− аксо-соматические, когда аксон первого нейрона контактирует с телом второго,
− аксо-дендритические, когда аксон первого нейрона контактирует с дендритов второго,
− аксо-аксональные, когда аксон первого нейрона контактирует с аксоном второго.
Синапсы имеют две части:
− пресинаптический, полюс и
Пресинаптический полюс – это концевое расширение аксона первого нейрона; содержит скопления митохондрий и пресинаптические пузырьки, имеющие в своем составе медиатор (ацетилхолин или норадреналин) и пресинаптическую мембрану (часть аксолеммы концевого расширения).
Постсинаптический полюс – это прилежащая часть второго нейрона. Имеет постсинаптическую мембрану.
Между пре- и постсинаптической мембраной располагается синаптическая щель. Основное свойство синапса – одностороннее проведение нервных импульсов от пресинаптического полюса в постсинаптический.
Область, раздражение которой приводит к возбуждению чувствительной единицы, называют рецепторным полем. Чем больше размер рецепторного поля, тем меньшей остротой сенсорной чувствительности обладает данная область: например, в верхней части руки рецепторные поля занимают площадь 2 см 2 , в области запястья — 1 см 2 , на подушечках пальцев — 5 мм 2 .
Чувствительные единицы переплетаются между собой, за счет чего становится возможным одновременное восприятие одним участком кожи разных видов чувствительности.
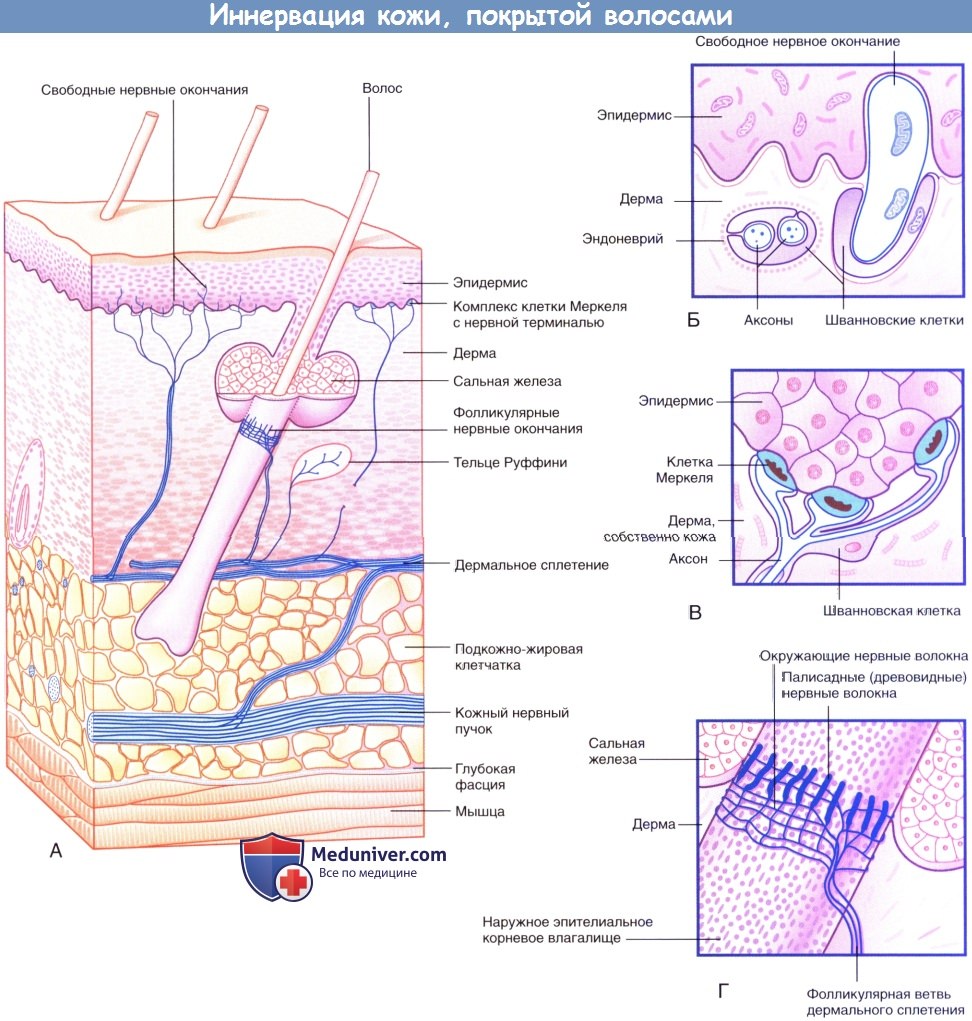
Иннервация кожи, покрытой волосами.
(А) Три морфологических типа чувствительных нервных окончаний в коже, покрытой волосами.
(Б) Свободные нервные окончания в базальном слое эпидермиса.
(В) Комплекс клетки Меркеля с нервной терминалью.
(Г) Палисадные и циркулярные нервные окончания на поверхности наружного корневого влагалища волоса.
б) Нервные окончания:
1. Свободные нервные окончания. По мере приближения к поверхности кожи многие чувствительные нервные волокна утрачивают периневральную, а затем и миелиновую оболочку (в случае ее наличия). Впоследствии нервные волокна разветвляются и формируют субэпидермальное нервное сплетение. Аксон освобождается от оболочек, сформированных шванновскими клетками, что позволяет ему, разветвляясь между коллагеновыми пучками дермы, образовывать дермальные нервные окончания, а внутри эпидермиса — эпидермальные нервные окончания.
2. Фолликулярные нервные окончания. Нервные окончания волосяного фолликула представлены палисадными нервными волокнами, образованными обнаженными терминалями миелинизированных нервных волокон, расположенными на поверхности наружного корневого влагалища волосяных фолликулов ниже уровня сальных желез, а также циркулярными нервными окончаниями. Каждая фолликулярная единица иннервирует несколько волосяных фолликулов и образует множественные перекресты. Фолликулярные единицы — быстро адаптирующиеся: они возбуждаются при изменении положения волос, однако при сохранении этого положения возбуждения не происходит. Человек, одеваясь, чувствует давление одежды, но затем за счет быстрой адаптации вскоре перестает ощущать ее прикосновение. Иннервация волос у других млекопитающих организована сложнее. Иннервация волосяных фолликулов осуществляется тремя типами механорецепторов, каждый из которых передает информацию определенным структурам мозга, что свидетельствует о важности выполняемой ими чувствительной функции.
3. Комплексы клетки Меркеля с нервной терминалью. Нервная терминаль, расширяясь в области базального слоя эпидермальных гребешков и бороздок, образует комплекс с осязательным тельцем овальной формы — клеткой Меркеля. Комплексы клетки Меркеля с нервной терминалью — медленно адаптирующиеся. В ответ на продолжительное давление (например, при удержании ручки или ношении очков) эти комплексы непрерывно генерируют нервные импульсы. Комплексы клетки Меркеля с нервной терминалью особенно хорошо распознают края удерживаемых в руке предметов.
4. Инкапсулированные нервные окончания. Капсулы описанных ниже свободных нервных окончаний состоят из трех слоев: наружный слой представлен соединительной тканью, средний — периневральным эпителием, а внутренний — видоизмененными шванновскими клетками (телоглией). Инкапсулированные нервные окончания являются механорецепторами, преобразующими механическое воздействие в нервный импульс.
• Тельца Мейснера в большом количестве находятся в подушечках пальцев и расположены вблизи бороздок эпидермиса. Тельца представляют собой клетки овальной формы, внутри которых аксоны располагаются зигзагообразно между уплощенными клетками телоглии. Тельца Мейснера— быстро адаптирующиеся, вместе с медленно адаптирующимися комплексами клетки Меркеля с нервной терминалью они обеспечивают точное восприятие текстур (например, текстуры ткани одежды или поверхности дерева), а также рельефных поверхностей (например, шрифта Брайля). Такие кожные рецепторы способны воспринимать изменение рельефа поверхности даже на высоту 5 нм.
• Тельца Руффини присутствуют как на гладкой коже, лишенной волос, так и на коже с волосами. Они воспринимают плавные скользящие касательные прикосновения и являются медленно адаптирующимися. Внутреннее строение телец сходно со строением сухожильных органов Гольджи: аксоны образуют разветвления в центральной части телец, представленной коллагеновыми волокнами.
• Тельца Пачини по величине соответствуют размерам рисового зерна. В области кисти имеется около 300 телец, которые преимущественно сконцентрированы на боковых участках пальцев и ладони. Тельца Пачини расположены подкожно, близко к надкостнице. Несколько слоев периневрального эпителия внутри соединительнотканной капсулы расположены овально и по форме напоминают луковицу в разрезе. В центральной части тельца Пачини несколько пластинок телоглии окружают единичный аксон, который, попадая в тельце, утрачивает миелиновую оболочку. Тельца Пачини — быстро адаптирующиеся рецепторы преимущественно вибрационной чувствительности. Эти структуры особенно восприимчивы к вибрации костной ткани: большое количество телец расположено в надкостнице длинных трубчатых костей.
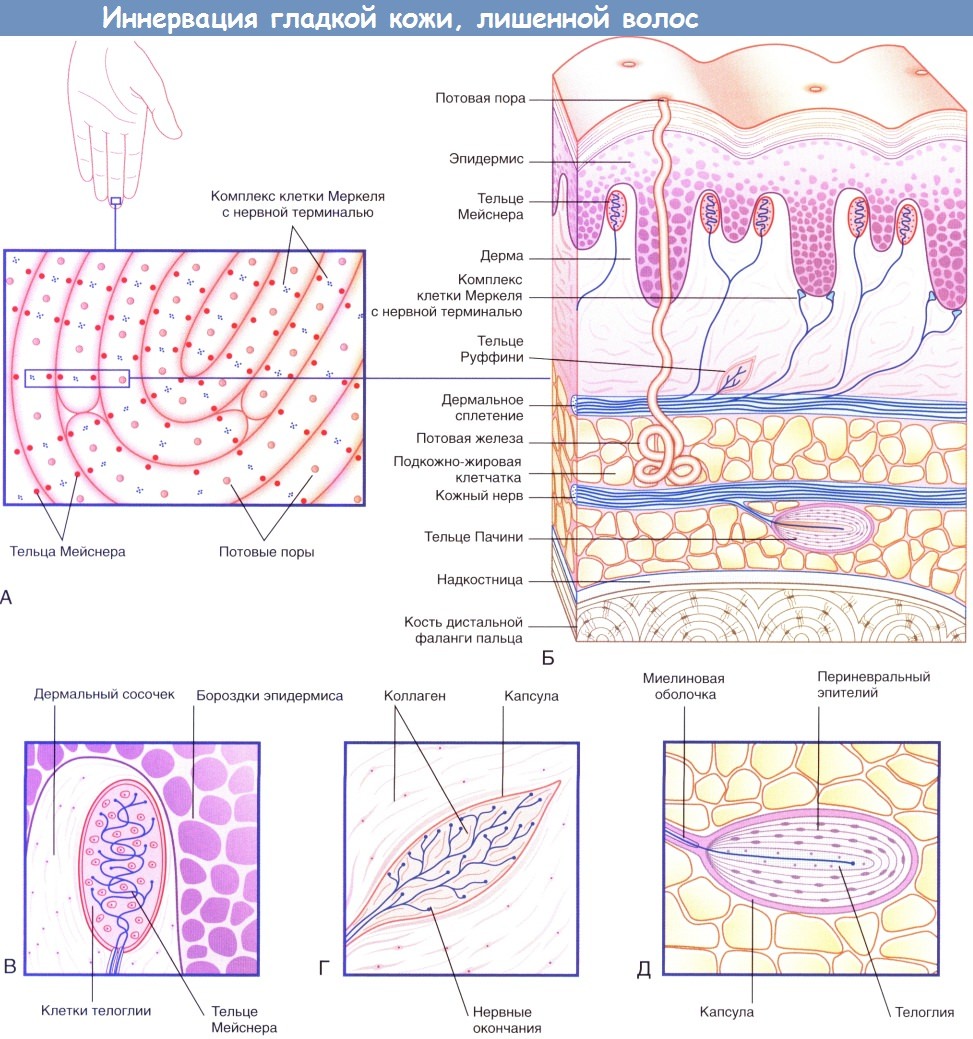
Иннервация гладкой кожи, лишенной волос.
(А) На подушечках пальцев располагаются нервные окончания двух видов.
(Б) На схеме строения участка кожи с изображения (А) представлены четыре типа чувствительных нервных окончаний.
(В) Тельца Мейснера.
(Г) Тельца Руффини.
(Д) Тельца Пачини.
Специалисты по физиологии чувствительности выделяют следующие виды рецепторов, локализованных в коже пальцев.
• Комплексы клетки Меркеля с нервной терминалью — медленно адаптирующиеся рецепторы I типа (MAP I).
• Тельца Мейснера — быстро адаптирующиеся рецепторы I типа (БАР I).
• Тельца Руффини — медленно адаптирующиеся рецепторы II типа (MAP II).
• Тельца Пачини — быстро адаптирующиеся рецепторы II типа (БАР II).
1. Полимодальные ноцицепторы преобразуют действие болевого раздражителя в нервные импульсы.
2. Аксоны посылают нервные импульсы в центральную нервную систему не только в обычном ортодромном направлении, но и в противоположном антидромном направлении от мест бифуркации к прилежащим участкам кожи. Ответная реакция ноцицептивных нервных окончаний на антидромную стимуляцию проявляется в высвобождении пептидных веществ, среди которых в большом количестве представлена субстанция Р.
3. Субстанция Р связывается с рецепторами на стенках артериол и вызывает их расширение, что приводит к появлению гиперемии.
4. Кроме того, субстанция Р связывается с рецепторами на поверхности тучных клеток, что приводит к высвобождению из них гистамина. Гистамин увеличивает проницаемость капилляров, за счет чего происходит местное накопление тканевой жидкости, обусловливающее возникновение бледного отечного валика.
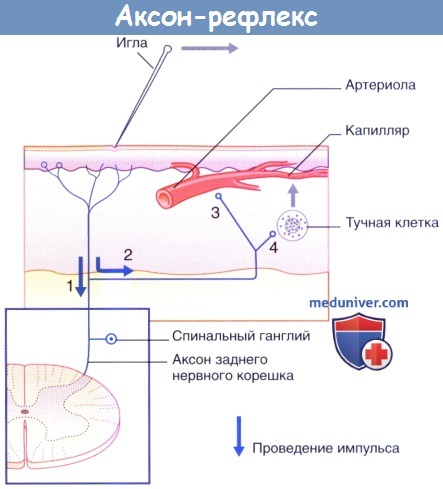
д) Резюме. Направляющиеся к коже нервы разветвляются и образуют дермальное нервное сплетение. Чувствительные нервные волокна дермального сплетения разветвляются и перекрывают друг друга. Каждое стволовое нервное волокно и его рецепторы формируют чувствительную единицу. Область, иннервируемую стволовым нервным волокном, называют его рецептивным полем.
К чувствительным единицам со свободными нервными окончаниями относят рецепторы температурной чувствительности, а также механические и температурные рецепторы болевой чувствительности. Рецепторы волосяных фолликулов—быстро адаптирующиеся осязательные механорецепторы, которые активируются только при движении волос. Комплексы клеток Меркеля с нервными терминалями обеспечивают восприятие края предметов, их относят к медленно адаптирующимся.
Редактор: Искандер Милевски. Дата публикации: 13.11.2018
Читайте также:
