
Что такое ганглия у червей
НЕРВНАЯ СИСТЕМА И ОРГАНЫ ЧУВСТВ. РЕФЛЕКСЫ
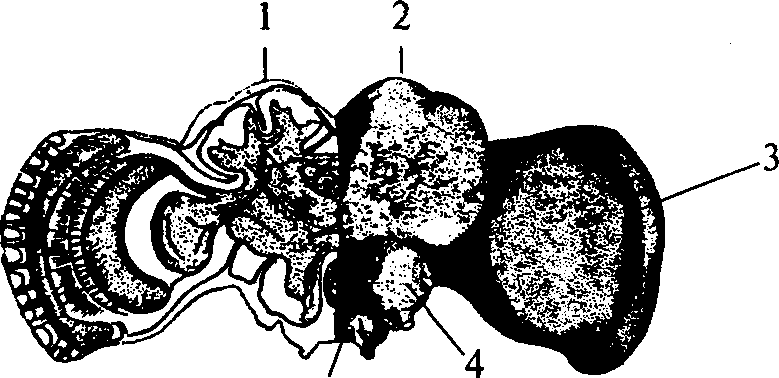
Вдоль средней линии брюшной стороны тела у дождевых червей под мускулатурой тянется нервный ствол, называемый брюшной нервной цепочкой. В каждом сегменте тела имеется нервный узел, или ганглий, представляющий собой скопление нервных клеток и отдающий от себя 3 пары нервов. Ганглии соединены друг с другом перемычками, коннективами, которые, кроме нервных волокон, также содержат нервные клетки. В переднем конце тела, в 3-м сегменте, брюшная нервная цепочка разделяется на правую и левую глоточные коннективы, образующие окологлоточное нервное кольцо, соединяющееся с надглоточным, или головным ганглием (рис. 17).
Этот ганглий — парный, состоит из правой и левой половин, плотно соединенных друг с другом. Но в отличие от всех остальных нервных узлов, лежащих на брюшной стороне тела под кишечником этот ганглий находится на спинной стороне тела и лежит над кишечником. Ганглий этот можно противопоставить всем прочим и вследствие того, что морфологически он сравним с головным мозгом вышестоящих форм (членистоногих). От него идут вперед многочисленные нервные стволы, обильно разветвляющиеся и образующие густые нервные сплетения в первых трех сегментах. Под глоткой, в месте расхождения глоточных коннектив, лежит подглоточный ганглий, представляющий собой результат слияния нескольких ганглиев брюшной нервной цепочки.
Как видно на поперечных разрезах, нервные клетки лежат по периферии ганглия, а средняя его часть занята сплетением отростков нервных клеток (рис. 18). В брюшной нервной цепочке обращают на себя внимание три очень толстых волокна, пробегающих но всей длине тела червя под соединительнотканной капсулой нервной цепочки на ее спинной стороне. Это так называемые нейрохорды, которые еще совсем недавно принимали за гигантские нервные волокна. Однако теперь окончательно выяснено, что они являются своеобразными опорными образованиями (Невмывака, 19476).
Эти образования и по строению, и по функции, и по положению между нервной системой и кишечником сходны с хордой позвоночных. Брюшная часть поперечного среза через дождевого червя Брюшная часть поперечного среза через дождевого червя Нервы, отходящие от ганглиев брюшной цепочки, содержат двигательные волокна, заканчивающиеся в мускулатуре, и чувствительные, по которым раздражения поступают в нервную систему с периферии. Тела чувствительных нервных клеток находятся на периферии, в том числе в наружном эпителии (рис. 18).
Нервные клетки здесь стоят в ряду эпителиальных клеток. Этот чрезвычайно древний тип соотношений элементов нервной системы сохранился у дождевых червей от их далеких предков, примитивных многоклеточных животных. Очень интересно, что чувствительными нервными клетками здесь становятся не только клетки наружного эпителия, но, как это недавно установлено, также и клетки кишечника, происходящие из внутреннего зародышевого листка (Невмывака, 1947а). Чувствительные нервные клетки и их окончания имеются и в других местах тела. Ими богато снабжены также нефридии, щетинковые мешки и другие органы. Таким образом, у дождевых червей, как и у высших животных, работа внутренних органов протекает под контролем регулирующей и централизующей роли нервной системы. Из рефлексов дождевых червей лучше всего известны наблюдаемые в акте ползания.
При движении червя по всей длине тела, от переднего конца до заднего, пробегают перистальтические волны комбинированных сокращений мускулатуры. Они следуют друг за другом, и каждая следующая волна может возникнуть еще задолго до того, как первая дойдет до заднего конца тела. Казалось бы очевидным, по аналогии с высшими животными, что причиной этих волн сокращений служит последовательная передача раздражений вдоль по брюшной нервной цепочке. Однако, к удивлению исследователей, выяснилось, что перерезание брюшного нервного ствола и даже вырезание из нее нескольких нервных узлов не останавливают бегущих волн сокращений мускулатуры: волна сокращения проходит через место повреждения так же, как это было и у нормального червя. С тем же результатом можно вдобавок к разрыву брюшной нервной цепочки удалить мускулатуру нескольких сегментов или повредить ее кислотой. Анализ этих и аналогичных опытов показал, что поступательное движение червя представляет собой длинную цепь рефлекторных актов, в которой каждый сегмент является в значительной мере автономной физиологической единицей.
Раздражения с периферии приводят к сокращению мускулатуры данного сегмента. Вследствие этого сокращения раздражаются периферические аппараты в соседнем сегменте, которые вызывают и в нем сокращения и т. д. Таким образом, комбинированные сокращения мышц в каждом сегменте могут представлять собой самостоятельный рефлекс, начинающийся с возбуждения чувствительных клеток на периферии и кончающийся эффектом сокращения мускулатуры этого сегмента. Это наиболее примитивный тип реакции на внешние воздействия. Некоторым его усложнением является передача полученного раздражения по нервной цепочке в соседний задний сегмент тела, в ответ на которое мускулатура этого сегмента сокращается. На рис. 19 изображена схема рефлекса при дугообразном сгибании червя, когда волна сокращений мускулатуры идет по одной стороне тела. Этот рефлекс и является основным при осуществлении поступательного движения червя вперед. Этот способ передачи раздражений по телу, как указано выше, говорит о слабой централизации его нервной системы.
О том же говорят и опыты с удалением надглоточного ганглия. Выше отмечено, что морфологически надглоточный ганглий можно сопоставить с головным мозгом вышестоящих форм (членистоногих). У многих морских кольчецов надглоточный ганглий имеет довольно сложное устройство. Однако у наземных червей надглоточный ганглий подвергся упрощению и физиологическая роль его очень невелика.
При слабом раздражении заднего конца тела (при легком прикосновении) червь ползет вперед, при раздражении спереди он быстро весь сокращается и ползет в другую сторону, при сильном раздражении в любом месте червь начинает сильно сокращаться дугообразно, в разных направлениях (так называемые гимнастические движения); черви обнаруживают быстрые реакции на свет, на запахи и т. д. Таким образом, указанные выше несовершенство нервной системы и ее слабая централизация обнаруживаются только при внимательном наблюдении и в специально поставленных экспериментах. Итак, нам известно, что червь располагает довольно богатым арсеналом возможностей для осуществления тех или иных реакций на изменения, происходящие в окружающей их среде. Рассмотрим теперь, каким путем он может распознавать эти изменения. Средствами для этого являются органы чувств. Как уже сказано, вся поверхность тела червя покрыта огромным количеством чувствительных нервных клеток. Эти клетки служат органами осязания, развитого у червей очень сильно. Известно, что достаточно, осторожно приблизившись, слабо дунуть на червя, чтобы он ответил резким сокращением продольной мускулатуры; с помощью такого движения он прячется в норку. Вдобавок к чувствительным нервным клеткам в наружном эпителии между клетками имеется очень большое количество свободных нервных окончаний, которые, скорее всего, также обеспечивают функцию осязания.
Светочувствительная клетка из кожного эпителия дождевого червя Светочувствительная клетка из кожного эпителия дождевого червя Как известно уже более ста лет назад, дождевые черви, несмотря на отсутствие глаз, хорошо воспринимают свет. Восприятие света производится особыми светочувствительными клетками, которые по большей части располагаются поодиночке между клетками наружного эпителия (рис. 20). Внутри этих клеток, кроме ядра и густой сети тончайших волоконец — нейрофибрилл, имеется прозрачное светопреломляющее тельце бобовидной или удлиненной формы; его называют линзой по аналогии с хрусталиком глаза более высоко организованных животных. От тела клетки отходит нервный отросток, вступающий в подкожное нервное сплетение и связывающий ее с центральной нервной системой. Такая клетка несомненно представляет собой простейший глаз, как бы изолированную и автономную клетку сетчатки глаза высших животных.
Светочувствительные клетки сосредоточены главным образом в передних сегментах тела; больше всего их в головной лопасти, где их может быть свыше 50 (рис. 21). В последующих сегментах число их быстро убывает, в середине тела их не находят, а в последних трех сегментах их снова становится больше. У некоторых видов дождевых червей, кроме изолированных светочувствительных клеток в наружном эпителии, имеются большие группы светочувствительных клеток, расположенные под кожей по ходу нервов, особенно в головной лопасти (рис. 22). Дождевой червь. Разрез через головную лопасть со светочувствительными клетками Дождевой червь. Разрез через головную лопасть со светочувствительными клетками Группа светочувствительных клеток, расположенных по ходу нерва Группа светочувствительных клеток, расположенных по ходу нерва Дарвин тщательно исследовал восприятие света дождевыми червями. Он нашел, что если осторожно освещать их бесшумным фонарем, имеющим только узкий пучок света, интенсивность которого уменьшена красным или синим стеклом (цвет стекла безразличен), то только очень немногие черви реагируют, а именно: уходят в свои норы. Дарвин вел наблюдения над теми видами, которые ночью выходят из нор в поисках пищи или для спаривания; это большой красный червь (Lumbricus terrestris), длинный червь (Allolobophora longa) и некоторые другие. Задний конец у них обычно остается в норке.
Но вряд ли можно сомневаться в том, что ими же осуществляются и функции обоняния и вкуса. В пользу такого заключения свидетельствует тот факт, что эти органы имеются в большом количестве в ротовой полости. Поперечный разрез через дождевого червя Поперечный разрез через дождевого червя Чувство обоняния, т. е. способность к распознаванию различных веществ в газообразном состоянии (что и является способностью воспринимать запахи), развито у червей сравнительно .слабо. В опытах Дарвина черви не реагировали на запах табачного сока, духов, уксусной кислоты, но куски лука (который они очень любят) и листья капусты они находили по запаху. На эфир, поднесенный на близком расстоянии к переднему концу тела, черви реагировали отрицательно и немедленно устранялись от него. Чувство вкуса, т. е. способность распознавать химические различия веществ при контакте с ними, развито у червей очень тонко и наряду с чувством осязания служит для них главным источником восприятия событий внешнего мира. Опытами Дарвина, развитыми в последнее время рядом исследователей, способность червей выбирать себе пищу доказана совершенно бесспорно и возражения, высказанные некоторыми авторами по этому поводу (например, Тарнани, 1928), несомненно основаны на ошибках.
Нужно сказать, что черви, собирая корм, попадающийся им на поверхности земли, никогда его глубоко не вносили в норки, а оставляли его недалеко от наружного отверстия или же только пододвигали к нему. Поэтому вышеуказанный подсчет сделать нетрудно. Если червь не различает концов испытателя вкуса, то при достаточно большом числе повторений опыта должно оказаться, что как за тот, так и за другой его конец червь берется одинаково часто. Если он предпочитает испытуемое вещество чистому желатину, то пропитанный им конец должен оказываться чаще впереди при затаскивании. Наоборот, если вещество оказывается на вкус хуже, чем чистый желатин, то за него червь должен ухватываться реже. Этот опыт видоизменяется подкидыванием червям испытателей вкуса, пропитанных разными веществами, с последующим определением числа втянутых в норки тех и других. Результаты обрабатывались статистически.
Проведенные опыты показали, что черви предпочитают гниющие листья только что опавшим осенним; еще менее нравятся им свежие зеленые листья и в еще меньшей степени — высушенные зеленые. Чистый желатин их привлекает более, чем высушенные листья. Гниющие листья разных растений могут быть выстроены в такой ряд в порядке убывающей склонности к ним червей: ива, сладкий лупин, грецкий орех, белая акация, тополь, дуб, горький лупин, липа, бук, клеи, конский каштан. Свежие листья располагаются совсем в другой последовательный ряд. От желатина с примесью хинина черви отказываются, причем чувствуют это вещество уже в концентрации 0.07%. От минеральных кислот они отказываются и любых концентрациях, но прибавка к желатину 1—2% лимонной и фосфорной кислот им нравится. К сахарам они относятся безразлично, но от очень крепких растворов Сахара отказываются вовсе. К сахарину обнаруживается отрицательная реакция уже начиная с ничтожных концентраций. Способность определять форму тел у червей, по-видимому, отсутствует.
Предпочтительное затаскивание ими в норки листьев за передний конец, а сосновой хвои за основание (факт, установленный Дарвином), дальнейшими исследованиями подтвержден. Однако опытами Мангольда установлено, что черви при этом руководствуются только чувством вкуса, позволяющим им отличать верхушку листа от черешка. Говоря о рефлекторной деятельности дождевых червей, необходимо отметить, что у них давно доказана способность к научению и изменению поведения в связи с пережитыми ранее ощущениями, т. е. условные рефлексы.
3. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ В ФИЛОГЕНЕЗЕ
Для беспозвоночных животных характерно наличие нескольких источников происхождения нервных клеток. У одного и того же типа животных нервные клетки могут одновременно и независимо происходить из трех разных зародышевых листков. Полигенез нервных клеток беспозвоночных является основой разнообразия медиаторных механизмов их нервной системы.
Нервная система впервые появляется у кишечнополостных животных. Кишечнополостные — это двухслойные животные. Их тело представляет собой полый мешок, внутренняя полость которого является пищеварительной полостью. Нервная система кишечнополостных принадлежит к диффузному типу. Каждая нервная клетка в ней длинными отростками соединена с несколькими соседними, образуя нервную сеть. Нервные клетки кишечнополостных не имеют специализированных поляризованных отростков. Их отростки проводят возбуждение в любую сторону и не образуют длинных проводящих путей. Контакты между нервными клетками диффузной нервной системы бывают нескольких типов. Существуют плазматические контакты, обеспечивающие непрерывность сети (анастомозы). Появляются и щелевидные контакты между отростками нервных клеток, подобные синапсам. Причем среди них существуют контакты, в которых синаптические пузырьки располагаются по обе стороны контакта — так называемые симметричные синапсы, а есть и несимметричные синапсы: в них везикулы располагаются только с одной стороны щели.
Нервные клетки типичного кишечнополостного животного гидры равномерно распределены по поверхности тела, образуя некоторые скопления в районе ротового отверстия и подошвы (рис. 8). Диффузная нервная сеть проводит возбуждение во всех направлениях. При этом волну распространяющегося возбуждения сопровождает волна мышечного сокращения.
Рис. 8. Схема строения диффузной нервной системы кишечнополостного животного:
1 — ротовое отверстие; 2 — щупальце; 3 — подошва
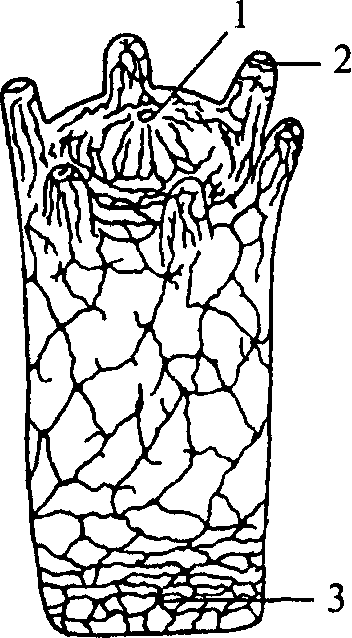
Рис. 9. Схема строения диффузностволовой нервной системы турбеллярии:
1 — нервный узел; 2 — глотка; 3 — брюшной продольный ствол; 4 — боковой нервный ствол
Следующим этапом развития беспозвоночных является появление трехслойных животных — плоских червей. Подобно кишечнополостным они имеют кишечную полость, сообщающуюся с внешней средой ротовым отверстием. Однако у них появляется третий зародышевый слой — мезодерма и двусторонний тип симметрии. Нервная система низших плоских червей принадлежит диффузному типу. Однако из диффузной сети уже обособляются несколько нервных стволов (рис. 9, 3, 4).
У свободно живущих плоских червей нервный аппарат приобретает черты централизации. Нервные элементы собираются в несколько продольных стволов (рис. 10, 4, 5) (для самых высокоорганизованных животных характерно наличие двух стволов), которые соединяются между собой поперечными волокнами (комиссурами) (рис. 10, 6). Упорядоченная таким образом нервная система называется ортогоном. Стволы ортогона представляют собой совокупность нервных клеток и их отростков (рис. 10).
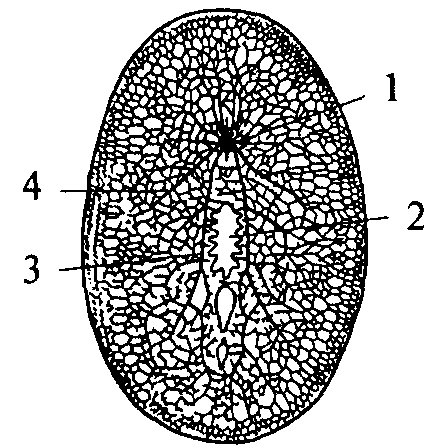
1 — щупальцевидный вырост; 2 — нерв, иннервирующий вырост; 3 — мозговой ганглий; 4 — боковой продольный нервный ствол; 5 — брюшной продольный нервный ствол; 6 — комиссура
ганглия появляются длинные отростки, идущие в продольные стволы ортогона (рис. 10, 4, 5).
Таким образом, ортогон представляет собой первый шаг к централизации нервного аппарата и его цефализации (появлению мозга). Централизация и цефализация являются результатом развития сенсорных (чувствительных) структур.
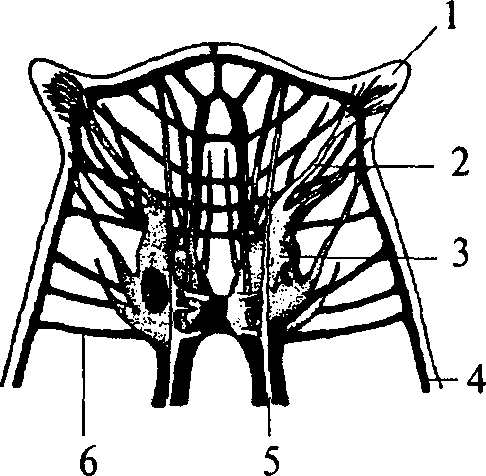
Следующим этапом развития беспозвоночных животных является появление сегментированных животных — кольчатых червей. Их тело метамерно, т.е. состоит из сегментов. Структурной основой нервной системы кольчатых червей является ганглий — парное скопление нервных клеток, расположенных по одному в каждом сегменте. Нервные клетки в ганглии размещаются по периферии. Центральную его часть занимает нейропиль — переплетение отростков нервных клеток и глиальные клетки. Ганглий расположен на брюшной стороне сегмента под кишечной трубкой. Он посылает свои чувствительные и двигательные волокна в свой сегмент и в два соседних. Таким образом, каждый ганглий имеет три пары боковых нервов, каждый из которых является смешанным и иннервирует свой сегмент. Приходящие с периферии чувствительные волокна попадают в ганглий через вентральные корешки нервов. Двигательные волокна выходят из ганглия по дорсальным корешкам нервов. Соответственно этому чувствительные нейроны расположены в вентральной части ганглия, а двигательные — в дорсальной. Кроме того, в ганглии есть мелкие клетки, иннервирующие внутренние органы (вегетативные элементы), они расположены латерально — между чувствительными и двигательными нейронами. Среди нейронов чувствительной, двигательной или ассоциативной зон ганглиев кольчатых червей не обнаружено группирования элементов, нейроны распределены диффузно, т.е. не образуют центров.
Ганглии кольчатых червей соединены между собой в цепочку. Каждый последующий ганглий связан с предыдущим при помощи
1 — надглоточный нервный ганглий;
2 — подглоточный нервный ганглий;
3 — сложный слившийся ганглий грудного сегмента; 4 — брюшной ганглий; 5 — периферический нерв; 6 — коннектива
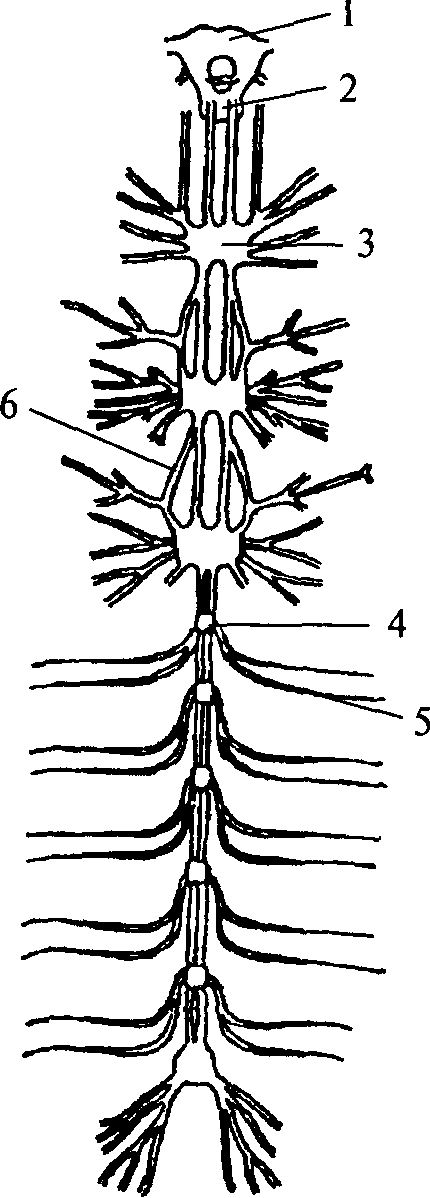
нервных стволов, которые называются коннективами. На переднем конце тела кольчатых червей два слившихся ганглия образуют крупный подглоточный нервный узел. Коннективы от подглоточного нервного узла, огибая глотку, вливаются в надглоточный нервный узел, который является самой ростральной (передней) частью нервной системы. В состав надглоточного нервного ганглия входят только чувствительные и ассоциативные нейроны. Двигательных элементов там не обнаружено. Таким образом, надглоточный ганглий кольчатых червей является высшим ассоциативным центром, он осуществляет контроль над подглоточным ганглием. Подглоточный ганглий контролирует нижележащие узлы, он имеет связи с двумя-тремя последующими ганглиями, тогда как остальные ганглии брюшной нервной цепочки не образуют связей длинней, чем до соседнего ганглия.
В филогенетическом ряду кольчатых червей есть группы с хорошо развитыми органами чувств (многощетинковые черви). У этих животных в надглоточном ганглии обособляются три отдела. Передний отдел иннервирует щупальца, средняя часть иннервирует глаза и антенны. И наконец, задняя часть развивается в связи с совершенствованием химических органов чувств.
Сходную структуру имеет нервная система членистоногих, т.е. построена по типу брюшной нервной цепочки, однако может достигать высокого уровня развития (рис. 11). Она включает в себя значительно развитый надглоточный ганглий, выполняющий функ-
1 — грибовидное тело; 2 — протоцеребрум; 3 — зрительная лопасть; 4 — дейтоцеребрум; 5 — тритоцеребрум
цию мозга, подглоточный ганглий, управляющий органами ротового аппарата, и сегментарные ганглии брюшной нервной цепочки. Ганглии брюшной нервной цепочки могут сливаться между собой, образуя сложные ганглиозные массы.
Головной мозг членистоногих состоит из трех отделов: переднего — протоцеребрума, среднего — дейтоцеребрума и заднего — тритоцеребрума. Сложным строением отличается мозг насекомых. Особенно важными ассоциативными центрами насекомых являются грибовидные тела, располагающиеся на поверхности протоцеребрума, причем чем более сложным поведением характеризуется вид, тем более развиты у него грибовидные тела. Поэтому наибольшего развития грибовидные тела достигают у общественных насекомых (рис. 12).
Практически во всех отделах нервной системы членистоногих существуют нейросекреторные клетки. Нейросекреты играют важную регулирующую роль в гормональных процессах членистоногих.
В процессе эволюции первоначально диффузно расположенные биполярные нейросекреторные клетки воспринимали сигналы либо отростками, либо всей поверхностью клетки, затем сформировались нейросекреторные центры, нейросекреторные тракты и нейросекреторные контактные области. В последующем произошла специализация нервных центров, увеличилась степень надежности во взаимоотношениях двух основных регуляторных систем (нервной и гуморальной) и сформировался принципиально новый этап регуляции — подчинение нейросекреторным центрам периферических эндокринных желез.
1 — церебральная комиссура; 2 — церебральные ганглии; 3 — педальные ганглии; 4 — коннектива; 5 — висцеральные ганглии
Нервная система моллюсков также имеет ганглионарное строение (рис. 13). У простейших представителей типа она состоит из нескольких пар ганглиев. Каждая пара ганглиев управляет определенной группой органов: ногой, висцеральными органами, легкими и т.д. — и расположена рядом с иннервируемыми органами или внутри их. Одноименные ганглии попарно соединены между собой комиссурами. Кроме того, каждый ганглий связан длинными коннективами с церебральным комплексом ганглиев.
У более высокоорганизованных моллюсков (головоногие) нервная система преобразуется (рис. 14). Ганглии ее сливаются и образуют общую окологлоточную массу — головной мозг. От заднего отдела головного мозга отходят два крупных мантийных нерва и образуют два больших звездчатых ганглия. Таким образом, у головоногих наблюдается высокая степень цефализации.
Мы об'единяем в этой группе несколько типов животных, имеющих между собой то общее, что их нервная система состоит из отдельных нервных узлов (ганглиев). В простейшем случае имеется всего два ганглия, расположенных вблизи переднего конца тела животного и дающих начало двум нервным тяжам — как это можно видеть у низших червей (например, у плоских червей — Plathelmintes). У более высокоорганизованных червей (у кольчатых червей — Annelides), имеющих расчлененное тело, состоящее из отдельных членов, в каждом сегменте находится одна пара ганглиев; самая передняя пара ганглиев, лежащая над глоткой, обозначается как головной мозг. Остальные ганглии расположены в отдельных отрезках, сегментах, всегда на брюшной стороне тела. В своей совокупности они образуют брюшную нервную цепь или брюшной мозг. Каждая пара ганглиев связана посредством поперечной комиссуры; ганглии каждого ряда (правого и левого) соединены продольным нервным тяжем.
Рисунок 5.1. Нервная система плоского, круглого червей, насекомого, моллюска
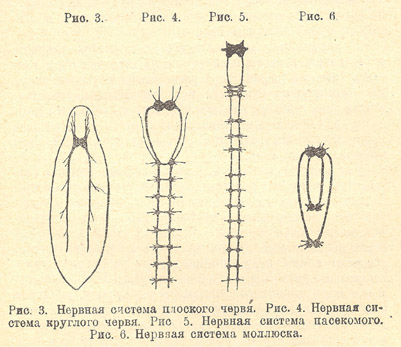
Рис. 3. Нервная система плоского червя
Рис. 4. Нервная система круглого червя
Рис. 5. Нервная система насекомого
Рис. 6. Нервная система моллюска
Подобную нервную систему мы находим у всех членистоногих: раков, многоножек, пауков и насекомых. Если мы рассмотрим нервную систему какого-нибудь насекомого, например, пчелы, то мы найдем в голове над глоткой два
ганглия, которые соединены в одно целое и образуют, таким образом, надглоточный ганглий, или мозг. Под глоткой расположены три ганглия брюшной нервной цепи; эти ганглии также слиты воедино и образуют подглоточный ганглий, от которого отходят нервы к ротовым частям. На остальном протяжении тела в каждом сегменте залегает пара ганглиев брюшного мозга. На Рис. 5.1, рис. 5 можно видеть три пары подглоточных ганглиев трех грудных сегментов, и, наконец, несколько пар ганглиев, залегающих в сегментах задней половины тела.
У мягкотелых, или моллюсков, т.-е. улиток, ракушек и каракатиц, в простейшем случае имеется три пары ганглиев (Рис. 5.1): первая пара расположена над ртом, это — так называемые головные, или церебральные ганглии; вторая пара ганглиев находится на ноге и называется ножными, или педальными ганглиями; третья пара ганглиев принадлежит внутренностям, это — внутренностные, или висцеральные ганглии.
Для всех тех животных, нервная система которых слагается из отдельных ганглиев, имеет силу следующее общее положение: каждая отдельная пара ганглиев может явиться рефлекторным центром, ибо сюда постоянно входят чувствующие нервы и отсюда выходят нервы двигательные; таким образом, все те части, которые содержат в себе пару ганглиев, могут производить самостоятельное движение в ответ на раздражение. Но деятельность всего животного связана с совокупным действием всех вместе взятых ганглиев, при чем, разумеется, на долю головной (мозговой) пары ганглиев, связанной с важнейшими органами чувств головы, выпадает в некотором роде руководящая роль.
Из всего многообразия животных, нервная система которых состоит из ганглиев, нам придется выбрать в качестве примеров всего лишь два-три типа животных; во-первых — ресничатых червей (Turbellaria), во-вторых — кольчатых червей (Annelides), и в-третьих — насекомых. У Turbellaria, или ресничатых червей, чувственные впечатления воспринимаются, главным образом, через посредство чувствующих клеток, расположенных в передней части тела, и затем передаются в мозг. Глаза ресничатого червя просты по строению и тоже связаны с мозгом. К мозгу примыкают два нервные ствола, идущие вдоль брюшной стороны тела животного (Рис. 5.1).
У более низко организованных ресничатых червей продольно идущие нервы еще содержат в себе ганглиозные клетки, тогда как у высших ресничатых червей эти продольные нервные тяжи обратились в простые проводящие пути, не содержащие нервных клеток. Взявши, напр., планарию (Planaria torva), мы можем, повторяя опыт Жака Лёба, разрезать животное поперек по середине, и тогда мы убедимся, что не только передняя половина животного, но и задняя его половина продолжает ползти, что, очевидно, стоит в связи с тем обстоятельством, что продольные волокна нервной системы, благодаря залегающим в них ганглиозным клеткам, обладают известной самостоятельностью. Если же мы таким же способом разрежем более высоко организованного червя (напр.,Thysanozoon), то лишь передняя часть червя будет продолжать ползать, задняя же его половина останется неподвижной.
Не приходится говорить о душе и применительно к ресничатым червям. Их жизнь регулируется рефлексами и инстинктами, при чем раздражения доходят к мозгу через посредство глаз и чувствующих клеток кожи.
Ресничатые черви обладают поразительной способностью регенерации. Судя по опытам, производившимся Рандольфом над одной американской планарией, животное можно подвергнуть поперечному сечению или разрезать по середине вдоль — и в обоих случаях каждая часть регенерирует и становится
целым животным. То же самое происходит даже и в том случае, если одним продольным и тремя поперечными разрезами расчленить животное на 8 частей, и даже если вырезать из тела червя небольшие доли. Поэтому мы так же мало можем приложить идею неделимой души по отношению к этим червям, как и к выше разобранным животным — книдариям.
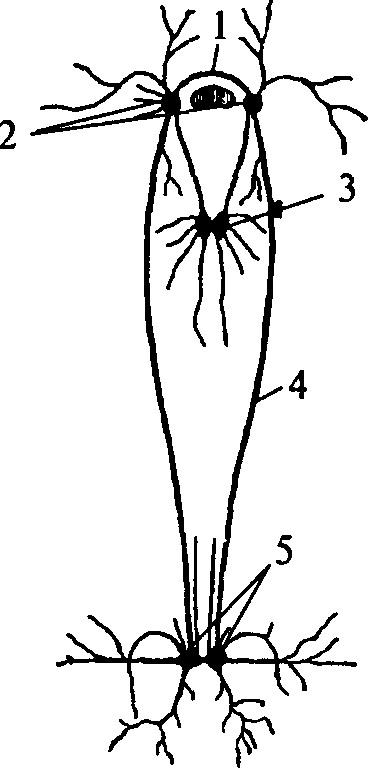
Кольчатые черви. Перейдем теперь к высшим червям, к кольчатым червям (Annelides), представителем которых может служить наш обыкновенный дождевой червь. Тело дождевого червя состоит из многочисленных колец, или члеников (сегментов). Первый сегмент расположен над ротовым отверстием и называется головной лопастью; на втором сегменте находится самое ротовое отверстие. Нервная система располагается таким образом, что спереди лежит мозг, а от него отходит брюшная нервная цепь.
В каждом сегменте имеется пара ганглиев (Рис. 5.1); ганглии соединены двумя продольными тяжами; образуется, таким образом, так называемая веревочно-лестничная нервная система.
Если разрезать дождевого червя на две или на три части, то все отрезки тела придут в оживленное движение. Отсюда можно заключить, что брюшные ганглии суть рефлекторные центры и обладают известной самостоятельностью. Правда, правильных движений ползания задняя . половина тела не производит, — эти движения обычно свойственны переднему концу; задние куски тела червя хотя и совершают оживленные бьющиеся движения, но ползут они лишь в том случае, если их подталкивать. Кусок передней части тела червя обнаруживает совершенно нормальный образ действий. Но можно сделать так, что и задний кусок разрезанного на части червя будет способен к нормальному движению ползания; для этого оказывается достаточным связать обе разрезанные части ниткой — тогда червь будет ползти как единое целое. Нитка тянет за собой задний кусок червя — и это ее действие играет роль возбудителя для произведения движений ползания задней части тела.
Если у дождевого червя вырезать мозг, то это повлечет серьезные повреждения. Животные тогда обычно не зарываются больше в землю, они живо ползают взад и вперед, при чем так быстро, что совсем не могут успокоиться. Они также теряют способность различения питательных веществ от посторонних предметов, так как их чувствующие клетки, расположенные на переднем конце тела, были связаны с мозгом.
Особняком стоит следующая проблема. Известно, что дождевые черви вносят в свои норы увядшие листья или маленькие листики салата и поедают их, при чем черви тащат листья таким образом, чтобы всего удобнее втянуть их в норку, т.-е. они берут их за черешок или за противоположный конец — за верхушку. И вот пробовали давать червям кусочки бумаги треугольной формы, и в этом случае черви тащили эти кусочки острым углом вперед.
Естественно было, в виду этого, склоняться к тому взгляду, чтобы приписать в данном случае червям некоторую долю размышления. Но недавние опыты Иордана доказали, что об'яснение этих действий червей может быть чисто механическое. Ночью черви хватают обрезки бумаги за самые различные места. Когда червь ощупью то там, то здесь касается края бумаги, то наибольшее противодействие бумаги он ощущает, приближаясь к углу ее, и он бессознательно тащит к себе именно угол. Совершенно то же, разумеется, имеет место и в случае втаскивания червем листа. Жизнь дождевого червя регулируется инстинктами и рефлексами. Нет решительно никаких оснований приписывать ему какие-либо высшие душевные способности. Мы не можем признать у червя с достоверностью ни одного чувства; правда, червь пораненный или полураздавленный производит оживленные движения, но эти движения могут быть чисто рефлекторные, с которыми не связывается никакого чувства. Мы впадаем в несомненный антропоморфизм, когда представляем себе, будто раздавленный червь извивается от боли. Мы не должны забывать, что нервная система червя настолько отлична от человеческой, что мы не в праве предполагать в нем душевной жизни или жизни чувств, аналогичной нашей. О том же свидетельствуют и опыты с регенерацией. Если разрезать дождевого червя на несколько частей, то каждая из них путем регенерации обратится в нового, целого, самостоятельного червя. То же имеет место у многих морских кольчатых червей — аннелид. Некоторые из них самостоятельно, сами делятся на несколько частей, из которых вырастают целые, отдельные животные. Учение о неделимой душе и здесь, таким образом, оказывается неприложимым.
Переходя от кольчатых червей к членистоногим , мы и у них видим сходно устроенную нервную систему. Остановимся только на насекомых . У них над глоткой находится мозг, или надглоточная пара ганглиев, от которых идут нервы к щупальцам и глазам; под глоткой находятся подглоточные ганглии, состоящие из трех, слившихся воедино, пар ганглиев, иннервирующих ротовые придатки; к подглоточным ганглиям примыкает брюшной мозг, с тремя парами ганглиев в трех грудных сегментах и с 5 — 10 парами ганглиев в сегментах задней половины тела (Рис. 5.1).
Сообразно этим анатомическим данным, является ожидаемой известная самостоятельность участков тела, не идущая, впрочем, так далеко, как у червей. Если, например, отрезать голову у мясной мухи, то в остальном теле мухи еще продолжают обнаруживаться различного рода инстинкты и рефлексы: животное может еще ходить, его задние лапки еще производят движения чистки, крылья трепещут (если схватить лапку или крыло); если положить животное на спину, оно пытается перевернуться — и, в ответ на более сильное раздражение, начинает летать вокруг. Здесь мы имеем дело не с выражением боли, — ибо ведь головы нет, — по с процессами возбуждения в ганглиях брюшной нервной цепи. С другой стороны, можно у пчелы или осы, сосущей мед, отрезать заднюю половину тела, и насекомое не прекращает сосания, чем доказывается, во-первых, отсутствие у животного болевых ощущений, во-вторых, то, что передние ганглии насекомого могут продолжать свою деятельность независимо от ганглиев задней половины тела.
Жизненные процессы у насекомых определяются рефлексами и инстинктами, анатомическим субстратом которых являются ганглии, и, главным образом — именно надглоточные ганглии. Многие насекомые обладают весьма сложными инстинктами; так, например, одиночные пчелы возводят постройки для своего выводка; некоторые роющие осы делают норы в земле, куда втаскивают гусениц для пищи своим личинкам; жук-навозник приготовляет катышек из навоза и катит его в свою земляную нору; черный жук-водолюб (Hydrophilus piceus) приготовляет тонко сработанный кокон для своих яиц. Все книги, трактующие о жизни насекомых, полны разнообразных рассказов об удивительных инстинктах насекомых.
У насекомых часто можно наблюдать, что инстинкт приводится в действие определенным чувственным раздражением, напр., каким-нибудь запахом, которой воспринимается через посредство щупалец (антенн). Проф. Форель вырезал у мясной мухи оба глаза и посадил ее затем на мертвого крота. Муха вскарабкалась на падаль и положила яйца на место поранения. Но когда Форель у такой мухи обрезал щупальца (антенны), то муха уже не обращала внимания на падаль.
У общественных насекомых [55] каждой отдельной форме животных отвечают определенные инстинкты; так, например, у пчел и у ос трутни, царицы и работницы имеют различные задачи, и, в соответствии с различием их инстинктов, различно устроенным оказывается и их мозг [56] . Совместная жизнь у социальных животных основана не на рассудочном мышлении, а исключительно на инстинктах. Хотя для целей взаимного общения насекомые не имеют языка, но у них есть выразительные движения, в которых выявляются определенные инстинкты. Так, напр., голодный муравей обыкновенно стучит щупальцами по голове другого муравья, в ответ на что и получает пищу. В муравейниках живут паразитирующие жуки, которые таким же способом выпрашивают у муравьев пищу, каковую вслед затем и получают. У пчел существуют различные звуки, действующие на всех сотоварищей по улью; например, звук, который означает призыв к роению, и звук, призывающий к тому, чтобы жалить, — звук аналогичный сигналу тревоги. Недавно К. Фриш показал, что те пчелы, которым удалось найти хороший корм, прилетев домой, исполняют на сотах род танца, что является для других пчел сигналом к немедленному отлету за добычей. Все эти выразительные движения и совершаются и воспринимаются инстинктивно.
И все же жизнь насекомых определяется не одними только рефлексами и инстинктами. У них является уже и память, разумеется, весьма различно развитая у различных форм. Топографической памятью (т.-е. памятью на места) должны обладать все те насекомые, которые возводят какие-либо сооружения, в противном случае эти животные не могли бы, отойдя в сторону, вновь отыскать оставленное место. У многих насекомых топографическая память распространяется также на те места, где можно найти корм. Проф. Форель наблюдал, что осы в часы его завтрака прилетали к столу, чтобы полакомиться засахаренными фруктами, и продолжали прилетать в то же самое время и тогда, когда на столе не имелось для них ничего привлекательного. Нечто аналогичное наблюдается и у пчел; и у них была установлена К. ф.-Фришем наличность памяти на запахи, цвета и формы предметов. Некоторые насекомые, как, напр., жук-водолюб (Hydrophilus piceus), могут становиться ручными, и ожидают пищи, которой их кормят из рук.
Таким образом, имеется достаточно доказательств того, что у многих насекомых наряду с унаследованными нервными путями, играют роль еще и пути, приобретенные ими в индивидуальной жизни, т.-е. что у насекомых наряду с рефлексами и инстинктами имеется также и память.
Читайте также:
