
Что такое мембрана нервной клетки
Внеклеточная и внутриклеточная жидкости представляют собой водно-солевые растворы, в которых множество растворимых молекул диссоциируют на положительно и отрицательно заряженные атомы,— ионы. Ионы и молекулы водных растворов находятся в состоянии постоянного возбуждения и осуществляют процесс диффузии—движения из областей высокой концентрации в область меньшей концентрации. Диффузия ионов происходит не только по градиенту концентрации, но и под влиянием электрических градиентов. Положительно заряженные ионы, например ионы натрия (Na + ) и калия (К + ), называют катионами, поскольку в электрическом поле они движутся к катоду.
Отрицательно заряженные ионы, например ионы хлора (Cl - ), называют анионами, так как они движутся к аноду. Одноименные заряды (например, Na + и К + ) отталкиваются, а разноименные (например, Na + и Cl - ) — притягиваются.
Клеточную мембрану можно рассматривать в качестве электрического конденсатора, поскольку она состоит из двух противоположно заряженных слоев, между которыми расположен изолирующий липидный слой. Электрический потенциал тканевой жидкости на плазматической мембране стремится к нулю (0 мВ) за счет нейтрализующих влияний анионов Cl - на катионы Na + (и другие катионы), а в цитозоле — за счет влияний белков-анионов на катионы К + .
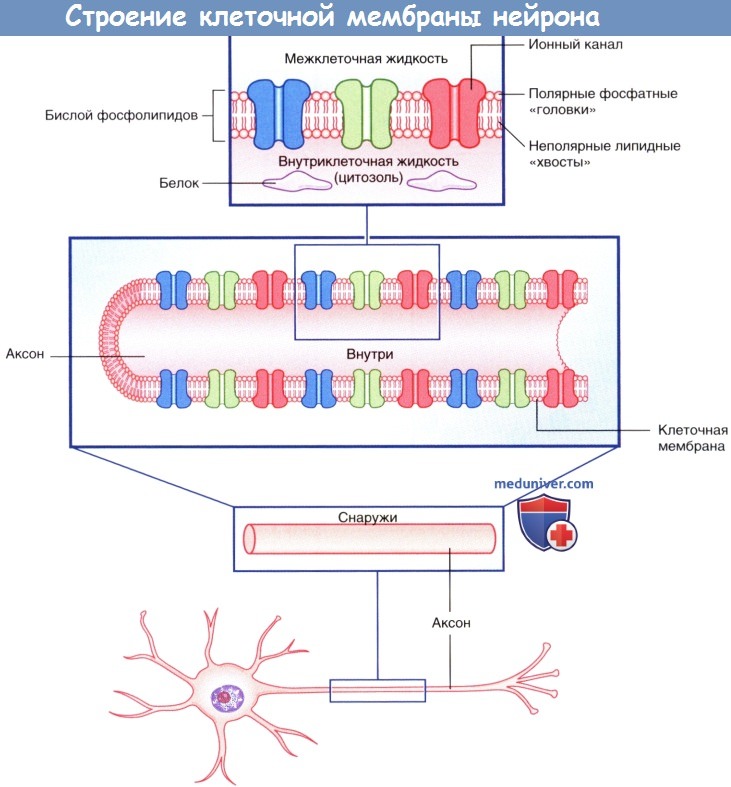
Строение клеточной мембраны нейрона.
Изображены мембранные белки, образующие ионные каналы.
а) Ионные каналы. Ионные каналы представляют собой трансмембранные белки, в центре которых находятся отверстия, по которым осуществляется транспорт ионов через клеточную мембрану. Большинство ионных каналов селективно пропускает определенные ионы, например, Na + , К + , Cl - . Выделяют несколько видов ионных каналов, из которых наиболее значимы первые три типа из следующих:
• Неуправляемые (независимые) ионные каналы постоянно находятся в открытом состоянии и осуществляют трансмембранный перенос ионов, создавая потенциал покоя на мембранах нейронов.
• Потенциалозависимые (потенциал-управляемые) ионные каналы содержат потенциалчувствительную цепь аминокислот, регулирующую открытие и закрытие отверстий ионных каналов в зависимости от изменений мембранного потенциала. Потенциал-управляемые ионные каналы играют важную роль в формировании потенциала действия.
• Энергозависимые транспортеры — ионные обменники (насосы) — обеспечивают постоянство концентраций ионов. Натрий-калиевый насос обеспечивает поддержание потенциала покоя.
• Хемоуправляемые (медиаторозависимые) ионные каналы нервной системы обеспечивают временное преобразование мембранного потенциала. В основном эти каналы представлены на постсинаптических мембранах. Активация хемоуправляемых ионных каналов может осуществляться молекулами медиаторов напрямую или опосредованно.
• Механочувствительные каналы активируются под действием физических раздражителей, вызывая деполяризацию мембраны и образование потенциалов действия, что обеспечивает восприятие определенных сигналов нервной системой. Каждый рецептор осуществляет преобразование специфических сигналов, например изменения длины или сократимости мышечных волокон, температурной и тактильной чувствительности кожи, хемочувствительности полости носа и рта или электромагнитных импульсов сетчатки.
На рисунке ниже изображены три неуправляемых ионных канала, обеспечивающих развитие потенциала покоя.
Если бы пассивная диффузия различных ионов через соответствующие им каналы осуществлялась свободно в одинаковом объеме, то не существовало бы разницы зарядов внутри и снаружи мембраны. В действительности проницаемость плазматической мембраны для ионов К + и Cl - намного выше, чем для ионов Na + , поскольку количество ионных каналов различается: калиевые каналы наиболее многочисленны, количество хлорных каналов вдвое меньше, а натриевые каналы встречают реже всего.
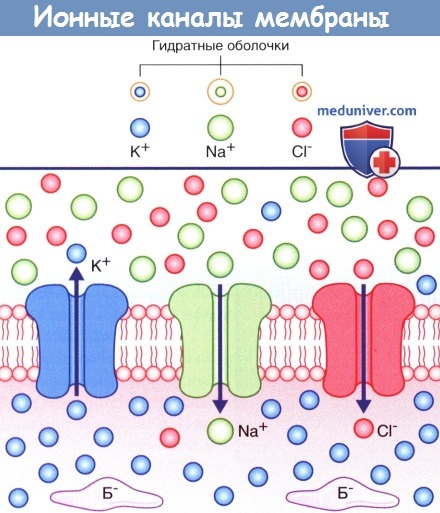
В состоянии покоя ионы Na + и Cl - сконцентрированы на внешней стороне плазматической мембраны за счет взаимного притяжения и образования толстой гидратной оболочки вокруг ионов Na + .
Ионы К + сконцентрированы на внутренней стороне плазматической мембраны за счет притяжения к белкам-анионам (Б - ).
Стрелками показано направление ионных градиентов концентрации.
б) Мембранный потенциал покоя. Мембранный потенциал невозбужденного нейрона (находящегося в состоянии покоя) формируется за счет разницы концентраций ионов Na + и К + во внеклеточной жидкости и цитозоле. Концентрация ионов К + в цитозоле выше, чем во внеклеточной жидкости в 20 раз, в то время как концентрация ионов Na + и Cl - во внеклеточной жидкости выше, чем в цитозоле в 10 и 3,8 раз, соответственно. Это обусловлено тем, что ионы К + под действием химических движущих сил переносятся из клетки в межклеточную жидкость, а ионы Na+ и Cl - — в обратном направлении.
На рисунке ниже показан вольтметр, электроды которого погружены в окружающую аксон внеклеточную жидкость. Один из электродов находится в стеклянной пипетке с тонким кончиком. В левой части рисунка, где оба электрода погружены во внеклеточную жидкость, разница электрического напряжения отсутствует, и вольтметр регистрирует нулевой показатель. В правой части рисунка пипетка проникает через плазматическую мембрану аксона в цитозоль, что позволяет оценить электрический заряд внутриклеточной жидкости. В данном случае разность электрических потенциалов составляет -70 мВ, однако у разных нейронов она может варьировать от -60 до -80 мВ.
Этот показатель отражает мембранный потенциал покоя, т.е. потенциал мембраны аксона, который не проводит электрические импульсы.
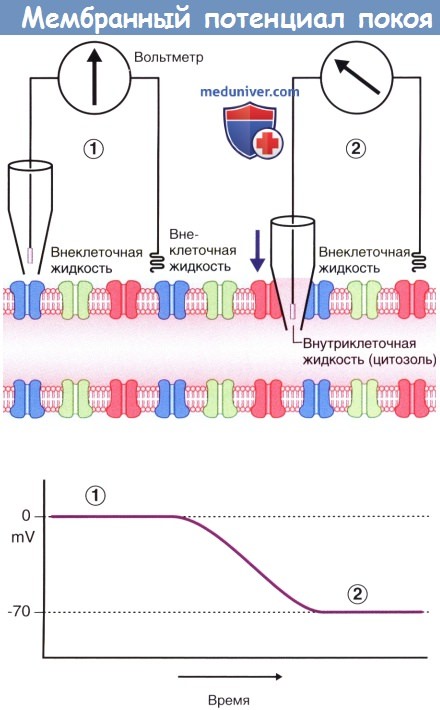
Мембранный потенциал покоя.
(1) Оба электрода вольтметра погружены во внеклеточную жидкость, окружающую аксон. Конец левого электрода находится в стеклянной пипетке.
Разница электрических потенциалов не регистрируется; на графике отмечены нулевые показатели.
(2) При опускании пипетки (направление указано стрелкой) производят пункцию плазматической мембраны для забора внутриклеточной жидкости.
Регистрируется разница электрических потенциалов величиной -70 мВ.
в) Проницаемость мембраны в состоянии покоя: 1. Ионы К + . На уровне ионного канала перемещение ионов К + осуществляется под электрическими влияниями притягивающего действия белков-анионов на внутренней стороне мембраны и отталкивающего действия ионов Na + на ее наружной стороне; при отсутствии этих факторов концентрация ионов К + на разных сторонах плазматической мембраны была бы одинаковой.
Концентрационный градиент на внешней стороне плазматической мембраны и электрический градиент потенциала на внутренней ее стороне уравновешиваются, когда мембранный потенциал достигает определенной величины, которую называют равновесным потенциалом для ионов К + (Ek). Его величина может быть рассчитана из уравнения Нернста, которое позволяет представить электрический градиент потенциала иона через его концентрационный градиент на основании законов термодинамики:
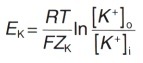
где Ek — равновесный потенциал для ионов К + (мВ)
R — газовая постоянная (8,31 Дж/(моль/°К)
Т — абсолютная температура в градусах Кельвина (310 К = 37 °С) F — число Фарадея (96500 Кл/моль)
Zk — валентность ионов К + (+1)
ln — натуральный логарифм
[К + ]0 — концентрация ионов K + с наружной стороны мембраны
[К + ]i — концентрация ионов К + с внутренней стороны мембраны.
Переведем натуральный логарифм в десятичный и рассчитаем значение показателя дроби:
Рассчитаем эти же значения для ионов Na + и Cl - :

Для расчета мембранного потенциала покоя применяют уравнение Гольдмана, учитывающее проницаемость мембраны для трех основных ионов и их приблизительное количество.
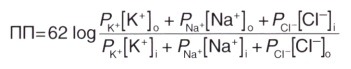
где ПП — потенциал покоя
62 — RT/Fx2,3 (постоянная для перевода натурального логарифма в десятичный)
Р — константа проницаемости мембраны для каждого из ионов (соответствует приблизительному количеству ионных каналов) о, i—концентрации ионов на внешней и внутренней сторонах мембраны, соответственно; концентрация отрицательно заряженных ионов Cl - внутри клетки расположена в числителе, а снаружи — в знаменателе дроби, поскольку log (X/Y) = log (Y/X).
Квадратными скобками обозначены концентрации ионов.
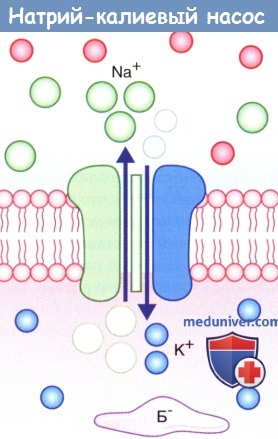
Натрий-калиевый насос.
На рисунке показан одновременный перенос трех ионов Na + из клетки в обмен на два иона К + в клетку.
Б - — белок-анион.
2. Натрий-калиевый насос. Под влиянием градиента концентрации происходит непрерывное перемещение ионов Na + внутрь клетки, а ионов К + — наружу, что нарушает постоянство потенциала покоя. Поддержание этого равновесия обеспечивает натрий-калиевый насос, корректирующий пассивное движение ионов. Натрий-калиевый насос представляет собой канал, способный перемещать ионы Na + наружу, а ионы К + — внутрь клетки. Во время работы насоса происходит перенос трех ионов Na + из клетки на каждые два иона К + в клетку.
Перенос обоих катионов осуществляется против градиента концентрации за счет энергии превращения АТФ в АДФ ферментом АТФазой. Активация этого фермента происходит при повышении концентрации ионов Na + в цитозоле.
Аксональная дегенерация при рассеянном склерозе возникает вследствие нарушения работы натрий-калиевого насоса в демиелинизированной аксолемме, что вызывает увеличение концентрации ионов Na + , которое в свою очередь приводит к повышению высвобождения ионов Са 2+ из внутриклеточного пула.
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
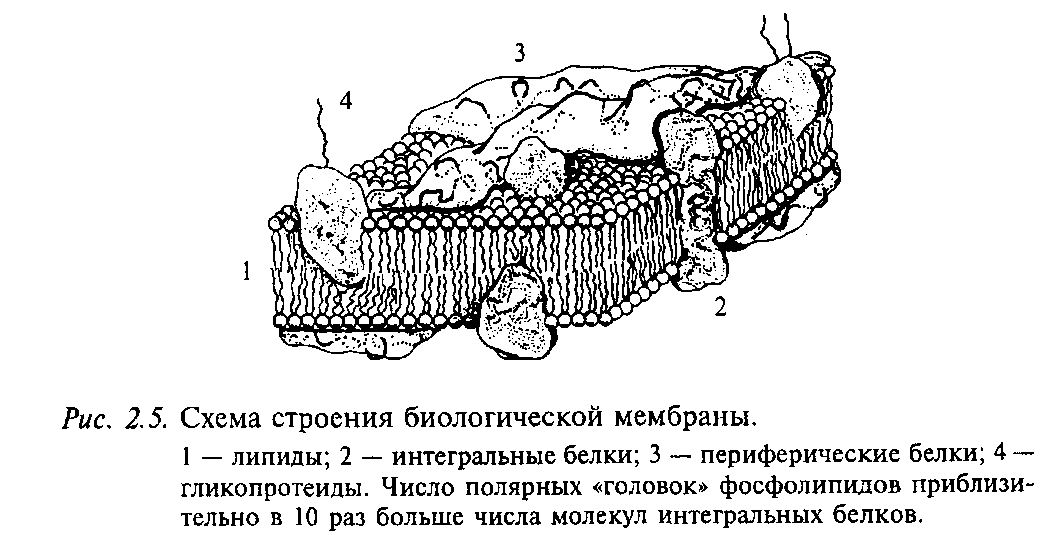
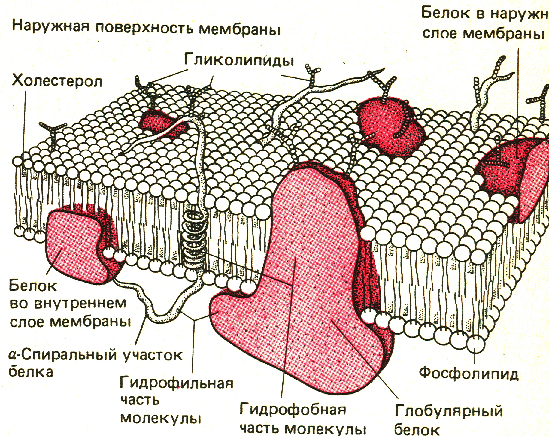
Мембрана нейрона, как и любой другой клетки, устроена очень сложно. В своей основе все известные биологические мембраны имеют однообразное строение (рис. 2.5): слой молекул белка, затем слой молекул липидов и еще один слой молекул белка. Вся эта конструкция напоминает два бутерброда, сложенных маслом друг к другу. Толщина такой мембраны составляет 7 - 11 нм. Чтобы представить эти размеры, вообразите, что толщина вашего волоса уменьшилась в 10 тыс. раз. В такую мембрану встроены разнообразные частицы. Одни из них являются частицами белка и пронизывают мембрану насквозь (интегральные белки), они образуют места прохождения для ряда ионов: натрия, калия, кальция, хлора. Это так называемые ионные каналы. Другие частицы прикреплены на внешней поверхности мембраны и состоят не только из молекул белка, но и из полисахаридов. Это рецепторы для молекул биологически активных веществ, например медиаторов, гормонов и др. Часто в состав рецептора, кроме места для связывания специфической молекулы, входит и ионный канал.
3.Механизмы и структуры пассивного транспорта.
Пассивный перенос веществ через клеточные мембраны не требует затраты энергии метаболизма. Активный транспорт осуществляется транспортнымиаденозинтрифосфатазами (АТФазами) и происходит за счет энергии гидролиза АТФ.
Схематически основные виды транспорта веществ через мембрану клеток представлены на рис.1.12
Рис.1.12 Виды пассивного и активного транспорта веществ через мембрану.
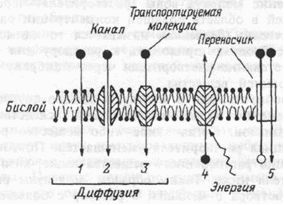
1,2 — простая диффузия через бислой и ионный канал, 3 — облегченная диффузия, 4 — первично-активный транспорт, 5 — вторично-активный транспорт.
2.1. Простая диффузия
Диффузия представляет собой процесс, при помощи которого газ или растворенные вещества распространяются и заполняют весь доступный объем.
Если концентрация вещества по обе стороны мембраны различна, возникает поток частиц, направленный из более концентрированного раствора в разбавленный. Диффузия происходит до тех пор, пока концентрация вещества по обе стороны мембраны не выравнивается. Через клеточную мембрану проходят как хорошо растворимые в воде <гидрофильные) вещества, так и гидрофобные, плохо или совсем в ней нерастворимые.
Гидрофобные, хорошо растворимые в жирах вещества, диффундируют благодаря растворению в липидах мембраны. Вода и вещества хорошо в ней растворимые проникают через временные дефекты углеводородной области мембраны, т.н. кинки, а также через поры, постоянно существующие гидрофильные участки мембраны.
В случае, когда клеточная мембрана непроницаема или плохо проницаема для растворенного вещества, но проницаема для воды, она подвергается действию осмотических сил. При более низкой концентрации вещества в клетке, чем в окружающей среде, клетка сжимается; если концентрация растворенного вещества в клетке выше, вода устремляется внутрь клетки. 2.2. Осмос
Осмос — движение молекул воды (растворителя) через мембрану из области меньшей в область большей концентрации растворенного вещества. Осмотическим давлением называется то наименьшее давление, которое необходимо приложить к раствору для того, чтобы предотвратить перетекание растворителя через мембрану в раствор с большей концентрацией вещества.
Объем клеток в значительной степени регулируется количеством содержащейся в них воды. Клетка никогда не находится в состоянии полного равновесия с окружающей средой. Непрерывное движение молекул и ионов через плазматическую мембрану изменяет концентрацию веществ в клетке и, соответственно, осмотическое
давление ее содержимого. Если клетка секретирует какое-либо вещество, то для поддержания неизменной величины осмотического давления она должна либо выделять соответствующее количество воды, либо поглощать эквивалентное количество иного вещества. Поскольку среда, окружающая большинство клеток гипотонична, для клеток важно предотвратить поступление в них больших количеств воды. Поддержание же постоянства объема даже в изотонической среде требует расхода энергии, поэтому в клетке концентрация веществ неспособных к диффузии (белков, нуклеиновых кислот и т.д.) выше, чем в околоклеточной среде. Кроме того, в клетке постоянно накапливаются метаболиты, что нарушает осмотическое равновесие. Необходимость расходования энергии для поддержания постоянства объема легко доказывается в экспериментах с охлаждением или ингибиторами метаболизма. В таких условиях клетки быстро набухают.
В целом объем клеток, не имеющих жестких стенок, определяется тремя факторами:
а) количеством содержащихся в них и неспособных к проникновению через мембрану веществ;
б) концентрацией в интерстиций соединений, способных проходить через мембрану;
в) соотношением скоростей проникновения и откачки веществ из клетки.
Большую роль в регуляции водного баланса между клеткой и окружающей средой играет эластичность плазматической мембраны, создающей гидростатическое давление, препятствующее поступлению воды в клетку. При наличии разности гидростатических давлений в двух областях среды вода может фильтроваться через поры барьера, разделяющего эти области.
Явления фильтрации лежат в основе многих физиологических процессов, таких, например, как образование первичной мочи в нефроне, обмен воды между кровью и тканевой жидкостью в капиллярах. 2.3. Диффузия ионов
Диффузия ионов происходит, в основном, через специализированные белковые структуры мембраны — ионные каналы, когда они находятся в открытом состоянии. В зависимости от вида ткани клетки могут иметь различный набор ионных каналов. Различают натриевые, калиевые, кальциевые, натрий-кальциевые и хлорные каналы. Перенос ионов по каналам имеет ряд особенностей, отличающих его от простой диффузии. В наибольшей степени это касается кальциевых каналов.
Ионные каналы могут находиться в открытом, закрытом и инак-тивированном состояниях. Переход канала из одного состояния в другое управляется или изменением электрической разности потенциалов на мембране, или взаимодействием физиологически активных веществ с рецепторами. Соответственно, ионные каналы подразделяют на потенциал-зависимые и рецептор-управляемые. Избирательная проницаемость ионного канала для конкретного иона определяется наличием специальных селективных фильтров в его устье. 2.4. Облегченная диффузия
Через биологические мембраны кроме воды и ионов путем простой диффузии проникают многие вещества (от этанола до сложных лекарственных препаратов). В то же время даже сранительно небольшие полярные молекулы, например, гликоли, моносахариды и аминокислоты практически не проникают через мембрану большинства клеток за счет простой диффузии. Их перенос осуществляется путем облегченной диффузии. Облегченной называется диффузия вещества по градиенту его концентрации, которая осуществляется при участии особых белковых молекул-переносчиков.
Транспорт Na + , K + , Сl - , Li + , Ca 2+ , НСО3 - и Н + могут также осуществлять специфические переносчики. Характерными чертами этого вида мембранного транспорта являются высокая по сравнению с простой диффузией скорость переноса вещества, зависимость от строения его молекул, насыщаемость, конкуренция и чувствительность к специфическим ингибиторам — соединениям, угнетающим облегченную диффузию.
Все перечисленные черты облегченной диффузии являются результатом специфичности белков-переносчиков и ограниченным их количеством в мембране. При достижении определенной концентрации переносимого вещества, когда все переносчики заняты транспортируемыми молекулами или ионами, дальнейшее ее увеличение не приведет к возрастанию числа переносимых частиц — явление насыщения. Вещества, сходные по строению молекул и транспортируемые одним и тем же переносчиком, будут конкурировать запереносчик — явление конкуренции.
Различают несколько видов транспорта веществ посредством облегченной диффузии (рис. 1.13):
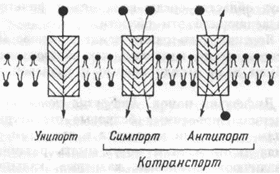
Рис. 1.13 Классификация способов переноса через мембрану.
Унипорт, когда молекулы или ионы переносятся через мебрану независимо от наличия или переноса других соединений (транспорт глюкозы, аминокислот через базальную мембрану эпителиоцитов);
Симпорт, при котором их перенос осуществляется одновременно и однонаправленно с другими соединениями (натрий- зависимый транспорт Сахаров и аминокислот Na + K + , 2Cl - и котран-спорт);
Антипорт — (транспорт вещества обусловлен одновременным и противоложно направленным транспортом другого соединения или иона (Na + /Ca 2+ , Na + /H + Сl - /НСО3 - — обмены).
Симпорт и антипорт — это виды котранспорта, при которых скорость переноса контролируется всеми участниками транспортного процесса.
Большинство клеток обладают системой облегченной диффузии. Однако перечень метаболитов, переносимых с помощью такого механизма, довольно ограничен. В основном, это сахара, аминокислоты и некоторые ионы. Соединения, являющиеся промежуточными продуктами обмена (фосфорилированные сахара, продукты метаболизма аминокислот, макроэрги), не транспортируются с помощью этой системы. Таким образом, облегченная диффузия служит для переноса тех молекул, которые клетка получает из окружающей среды. Исключением является транспорт органических молекул через эпителий, который будет рассмотрен отдельно.
§ 8. Заряды мембраны нервных клеток
Однако основные свойства нервной системы обусловлены способностью быстро реагировать на изменение ситуации внутри или вне организма. Скоростные процессы не могут осуществляться по медленным гуморальным законам, они происходят по законам электрохимическим. Нервные клетки способны получать, хранить, перерабатывать и передавать информацию при помощи специальной электрической активности. Они обладают зарядом мембраны — потенциалом покоя и могут его изменять в потенциал действия, который с высокой скоростью распространяется по телу клетки.
В основе потенциала покоя нервных клеток лежит баланс электрохимических и осмотических сил, которые действуют на клеточной границе — мембране. Мембрана клетки полупроницаема. Это означает, что через неё могут проникать далеко не все вещества. Мембрана всегда проницаема для воды, избирательно проницаема для определённых ионов и непроницаема для большинства органических соединений. Молекулы ДНК, РНК, белков и аминокислот находятся внутри клетки и не могут свободно диффундировать через мембрану. В соответствии с законами осмоса вода должна проникать в клетку. Поскольку мембрана клетки непроницаема для органических молекул, осмотическое равновесие достигнуто быть не может. Клетка должна была бы лопнуть. Этого не происходит, поскольку осмотическим силам оказывается постоянное противодействие со стороны сил совершенно другой природы.
Эти силы не осмотические, а электрохимические. Работа осмотических сил уравновешивается работой электрохимических. С одной стороны, это не позволяет клетке лопнуть, а с другой — является источником постоянного заряда мембраны нервной клетки. Внутри клетки находятся молекулы ДНК, РНК, белков, аминокислот и углеводов, которые имеют постоянный заряд. Как правило, этот заряд отрицателен и органические молекулы представляют собой набор внутриклеточных анионов (А). Их заряд уравновешивается внутри клетки положительно заряженными ионами калия (К + ). Снаружи клетки основным анионом является хлор (Cl - ), а катионом — натрий (Na + ). В абстрактной идеальной ситуации концентрации ионов должны были бы выровняться в результате диффузии через мембрану. Однако внутриклеточные анионы неподвижны, а специальные каналы для всех подвижных ионов обычно закрыты. Более того, специализированные ионные каналы постоянно откачивают избыток натрия и хлора из клетки и закачивают внутрь внеклеточный калий. Это процесс осуществляется с затратой энергии. Она тратится на то, чтобы создать такую величину заряда мембраны, чтобы её хватило для противодействия осмотическим силам, стремящимся разрушить клетку.
В реальной клетке основные проблемы с осмотическими (гидростатическими) силами обусловлены различиями в концентрации ионов калия и натрия по обе стороны мембраны. Некоторую роль в этом
процессе играют ионы натрия и подвижность воды, свободно движущейся через мембрану клетки. Тем не менее основное значение имеет калий, поскольку его концентрационные различия максимальны. Внутри клетки калия примерно в 40 раз больше, а натрия в 9 раз меньше, чем в межклеточном пространстве, поэтому калий стремится уравновесить ситуацию, двигаясь по концентрационному градиенту из клетки, а натрий — в клетку. Поскольку концентрации этих ионов внутри и снаружи клетки известны, можно выразить эти процессы в реальных физических величинах. Работа, которую надо выполнить для предотвращения движения ионов калия из клетки по концентрационному градиенту (А 0), будет равна:

где R — газовая постоянная; Т — абсолютная температура; [К + ] — молярная концентрация калия внутри (in) и снаружи клетки (out).
Противодействовать осмотическим силам, стремящимся вывести калий из клетки, могут только электрические силы. Равновесие может быть достигнуто только при равенстве работы осмотических и электрических сил (Аэ):
Для каждого иона может быть вычислена работа, необходимая для преодоления равновесия электрических сил, возникающих вследствие разделения зарядов по обе стороны мембраны:
где F — количество электрических зарядов в моле вещества (постоянная Фарадея); E — выраженная в вольтах разность электрических потенциалов, возникающая в результате разделения зарядов по обе стороны мембраны клетки.
Объединив формулы, легко получить известное уравнение Нернста, или равновесный (диффузионный) потенциал:

Подставив в формулу равновесия сил реальные значения при комнатной температуре, мы получим реальный заряд мембраны, который называют мембранным потенциалом. Обычно он колеблется в различных клетках от -60 до -90 мВ. При прямых измерениях зарядов мембран нервных клеток получены идентичные результаты. Обычная нервная клетка позвоночного обладает постоянным потенциалом мембраны около -75 мВ. Надо отметить, что подвижны ионы калия, натрия и хлора, поэтому при точных вычислениях мембранных потенциалов надо учитывать другие ионы.
Заряд мембраны и её способность изменять проницаемость для ионов под влиянием различных причин — уникальное эволюционное достижение. Этим свойством обладают многие клетки. Однако нервные клетки используют его для восприятия, передачи и хранения информации. Если при местном воздействии на нервную клетку возникает локальный участок с изменённым зарядом, то сигнал распространяется по мембране нервной клетки. Его обычно называют пассивным, а сам потенциал — градуальным. Это означает, что возникшее локальное возбуждение распространяется по мембране электротонически, что приводит к его постепенному затуханию. Обычно такие сигналы распространяются на небольшие расстояния, хотя у членистоногих известны нейроны, передающие такие сигналы на десятки миллиметров. Градуальные потенциалы образуют светочувствительные клетки сетчатки насекомых и позвоночных, многих периферических рецепторов и даже мотонейронов мышц стенки тела круглых червей.
Потенциал покоя есть у всех клеток на этой планете. Однако в нервной системе он является только отправной точкой для получения, хранения, обработки и передачи сигналов. Информация передаётся в нервных клетках при помощи динамического изменения заряда мембраны клетки. Это изменение может быть быстрым или медленным. Если оно происходит быстро и изменение заряда мембраны клетки превышает определённый порог, то возникает потенциал действия. Нейроны, формирующие потенциал действия, называют спайковыми. Потенциал действия отличается от градуального потенциала тем, что не затухает по мере движения от места возникновения. Причиной этого является способность мембраны активно распространять местную перезарядку мембраны, если изменение её потенциала достигнет определённого уровня. В обычной ситуации мембрана нервной клетки с потенциалом покоя -75 мВ должна изменить свой заряд до +40 мВ. Такое изменение приводит к формированию незатухающего потенциала действия, который распространяется по мембране клетки. Величину заряда мембраны, с которой начинается распространение сигнала, называют пороговой. Поскольку отростки нервных клеток достаточно хорошо изолированы глиальными клетками, а потенциал распространяется по мембране, его распространение скачкообразное (сальтаторное). В местах, где глиальные клетки плотно прилежат к нейронным мембранам, проведение происходит скачком: от одного перехвата Ранвье до другого. В перехватах мембрана отростков открыта, что позволяет осуществлять распространение сигнала путём открывания и закрывания калиевых и натриевых ионных каналов.
Принцип передачи сигнала по мембране нервной клетки довольно прост. В его основе лежит высокая плотность ионных каналов в мембране нервных клеток. Количество натриевых каналов может достигать в перехватах Ранвье 12 000 на 1 мкм 2 . Большое количество ионных каналов в мембранах нервных клеток является основой для распространения потенциалов действия. Натриевые и калиевые каналы расположены довольно плотно, что позволяет формировать локальные встречные потоки ионов при движении сигнала. Потенциал действия образуется в нейроне у аксонного холмика и начинается с открытия натриевых ионных каналов и проникновения натрия внутрь клетки. Поскольку в покое мембрана нейрона имеет заряд внутренней поверхности около -70 мВ, начинается небольшой сдвиг потенциала. Заряд мембраны локально уменьшается и постепенно доходит до нуля, а затем и до +40 мВ.
Изменение знака заряда мембраны приводит к закрыванию натриевых каналов и открыванию калиевых. Поток ионов калия восстанавливает исходный отрицательный заряд. Этот процесс называют кратковременной реверсией потенциала, или потенциалом действия. Возникнув в аксонном холмике, он распространяется по мембране клетки. Волна изменения заряда мембраны двигается с высокой скоростью, а весь процесс реверсии и восстановления занимает миллисекунды. Однако скорости движения потенциалов действия существенно различаются. На это влияют степень миелинизации, диаметр нервных волокон и многие другие факторы.
Скорости проведения потенциалов действия в нервной системе у разных видов животных различаются в десятки раз. Самые высокие скорости известны у аксонов человека и креветок (120–200 м/с), а самые низкие у актиний и медуз (0,1–0,5 м/с). По сути дела процессы передачи сигналов происходят в нервной системе со скоростью, которая отражает динамику и продолжительность жизни организма. Скорость нервных процессов определяет активность животного и внутреннее восприятие времени.
Надо отметить, что передаваемая информация кодируется частотой, последовательностью и продолжительностью активности импульсов, что обеспечивает высокую точность переносимой информации. Эти информационные сигналы различаются не только по принципам кодировки, но и по источникам — типам нервных клеток. Существует морфологическая и физиологическая классификация нейронов. По ведущим функциям нейроны физиологически подразделяются на сенсорные (афферентные), моторные, или двигательные (эфферентные), вставочные (ассоциативные, соединяющие афферентные и эфферентные) и нейросекреторные (гормональные клетки нервной системы). Эти типы не абсолютно жёсткие, а каждая клетка отчасти обладает всеми перечисленными функциями.
Читайте также:
