
Что такое нервные цепи
Диапроектор
НАГЛЯДНЫЕ ПОСОБИЯ.
Нервные цепи и сети. 15 мин.
Определение нервного центра по И.П.Павлову. 20 мин.
Введение. 5 мин.
Протокол №_19
Е.Б.ФИЛИППОВА
СТАРШИЙ ПРЕПОДАВАТЕЛЬ
кандидат биологических наук старший научный сотрудник
по НОРМАЛЬНОЙ ФИЗИОЛОГИИ
на тему: ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
для курсантов, слушателей и студентов 2 курсов
факультетов подготовки военных и гражданских врачей
Обсуждена и одобрена на заседании кафедры
Уточнено (дополнено):
СОДЕРЖАНИЕ
2. Синаптическая передача возбуждения в нервной системе 20 мин.
4. Процессы возбуждения и торможения в
нейронных цепях и сетях 15 мин.
5. Основные функциональные свойства
нервных центров 10 мин.
Заключение. 5 мин.
ЛИТЕРАТУРА
а) Использованная при подготовке текста лекции.
1. Физиология человека: Учебник: В 2-х т./Под ред. В.М. Покровского.- М.: Медицина,2003.
2.Физиология человека: Учебник / Под ред. Г.И. Косицкого. - М.: Медицина,1985.
3.Физиология человека: Учебник: В 3-х т. Под ред. Р. Шмидта, Г. Тевса. – 2-е изд.- М.: Мир,1996. Т.1
4. Гибадулин Т. В., Голубев В.Н. Свойства нервных центров. – С. Пб.: Изд-во ВмедА, 1987.
б) Рекомендуемая для самостоятельной работе по теме.
1.Основы физиологии человека: Учебник / Под ред. Б. И. Ткаченко.-
2. Коробков, А.В. Атлас по нормальной физиологии. /А. В. Коробков, С. А. Чеснокова.-М.: Высшая школа,1987
ТЕХНИЧЕСКИЕ СРЕДСТВА ОБУЧЕНИЯ
2. Мультимедийный проектор
На предыдущей лекции мы рассмотрели строение, свойства и функции отдельных нейронов, а также связи между нейронами в центральной нервной системе. Необходимо отметить, что в целостном организме нейроны не функционируют изолированно, а образуют нейронные цепи, ядра, сети и центры различной величины и численности.
Наиболее простой формой связи является нервная цепь, в которой возбуждение последовательно передается от одного нейрона к другому. Нервная цепь может содержать всего два нейрона с одним синапсом между ними, соответственно рефлекс, реализуемый через такое нервный центр, носит название моносинаптического. С увеличением числа нейронов и синапсов между ними время рефлекторной реакции на раздражитель увеличивается, поскольку каждый синапс имеет синаптическую задержку проведения. Такие рефлексы называют полисинаптическими. Сигналы по нервным сетям распространяются в одну сторону – за счет одностороннего проведения в синапсах.
Анатомическое понятие ядра означает совокупность нейронов, сосредоточенных в одном месте ЦНС, и связанных морфологически с исполнительным органом. Раздражение ядра приводит к изменению функции соответствующего органа. Например, возбуждение нейронов двигательных ядер передних рогов спинного мозга приводит к сокращению определенных групп мышц. Морфологическое понятие нервного центра означает скопление нейронов в определенном объеме, имеющих определенные, морфологически закрепленные связи с другими скоплениями нервных клеток и выполняющими определенные функции. Примером могут служить пирамидные клетки коры головного мозга и двигательные ядра передних рогов спинного мозга, которые, в свою очередь, связаны с определенными группами мышечных клеток. В ЦНС выделяют большое число таких центров: двигательные центры спинного мозга, центры глотания, рвоты, дефекации и др.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Нервные волокна, проводящие сигналы от рецепторов, передают их в центры головного мозга через несколько синаптических переключений. В синапсах происходит суммация и торможение сигналов от одного рецептора, а также взаимодействие сигналов от разных рецепторов.
Представим себе нервную цепь, состоящую из трех рецепторов и двух следующих за ними более высоких синаптических уровней. Средний рецептор усиливает свою активность при стимуляции и возбуждает те три нейрона, с которыми он образует синаптические связи. В результате такой дивергенции возбужденный участок, узко ограниченный на уровне рецепторов, расширяется, стимул становится менее четко локализованным. В то же время дивергенция обеспечивает проведение действия даже слабых стимулов от малого числа рецепторов к высшим уровням по многим волокнам, так что сигналы проходят параллельно через многие синапсы. В случае конвергенции каждый нейрон получает афференты от многих других нейронов, при этом происходит пространственная суммация синаптических потенциалов в этом нейроне. Конвергенция усиливает действия слабых стимулов, и даже слабое возбуждение может вызвать потенциал действия.
Если бы распространение нейронной активации продолжалось неопределенно долго, то вскоре произошло бы возбуждение всего головного мозга и различение качеств и места стимулов стало бы невозможным. Это предотвращается торможением, которое осуществляют тормозные вставочные нейроны. В некоторых сенсорных системах вышележащие центры тоже способны осуществлять торможение. Это так называемое центральное торможение может действовать на уровне самих рецепторов. Например, в органе слуха центрально управляемое торможение действует на разных синаптических уровнях, регулируя чувствительность органа.
Симнапс (греч. уэнбшйт, от ухнЬрфейн - обнимать, обхватывать, пожимать руку) - место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачинервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться.

Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном.
Структура синапса
Типичный синапс - аксодендритический химический. Такой синапс состоит из двух частей: пресинаптической, образованной булавовидным расширением окончанием аксона передающей клетки ипостсинаптической, представленной контактирующим участком цитолеммы воспринимающей клетки (в данном случае - участком дендрита). Синапс представляет собой пространство, разделяющее мембраны контактирующих клеток, к которым подходят нервные окончания. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём посредством прохождения ионов из одной клетки в другую.
Между обеими частями имеется синаптическая щель - промежуток шириной 10-50 нм между постсинаптической и пресинаптической мембранами, края которой укреплены межклеточными контактами.
Часть аксолеммы булавовидного расширения, прилежащая к синаптической щели, называется пресинаптической мембраной. Участок цитолеммы воспринимающей клетки, ограничивающий синаптическую щель с противоположной стороны, называется постсинаптической мембраной, в химических синапсах она рельефна и содержит многочисленные рецепторы.
В синаптическом расширении имеются мелкие везикулы, так называемые синаптические пузырьки, содержащие либо медиатор (вещество-посредник в передаче возбуждения), либо фермент, разрушающий этот медиатор. На постсинаптической, а часто и на пресинаптической мембранах присутствуют рецепторы к тому или иному медиатору.
Классификации синапсов
В зависимости от механизма передачи нервного импульса различают
· электрические - клетки соединяются высокопроницаемыми контактами с помощью особых коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе - 3,5 нм (обычное межклеточное - 20 нм). Так как сопротивление внеклеточной жидкости мало (в данном случае), импульсы проходят не задерживаясь через синапс. Электрические синапсы обычно бывают возбуждающими.
Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
· смешанные синапсы: Пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре - и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Наиболее распространены химические синапсы.
Химические синапсы можно классифицировать по их местоположению и принадлежности соответствующим структурам:
· аксо-дендритические - с дендритами, в т. ч.
· аксо-шипиковые - с дендритными шипиками, выростами на дендритах;
· аксо-соматические - с телами нейронов;
· аксо-аксональные - между аксонами;
· дендро-дендритические - между дендритами;
В зависимости от медиатора синапсы разделяются на
· аминергические, содержащие биогенные амины (например, серотонин, дофамин);
· в том числе адренергические, содержащие адреналин или норадреналин;
· холинергические, содержащие ацетилхолин;
· пуринергические, содержащие пурины;
· пептидергические, содержащие пептиды.
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По знаку действия:
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях.), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор - глицин) и ГАМК-ергические синапсы (медиатор - гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов:
1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала;
2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение. Синапс холинергический (s. cholinergica) - синапс, медиатором в котором является ацетилхолин.
В некоторых синапсах присутствует постсинаптическое уплотнение - электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические - симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. "Не-шипиковые" синапсы называются "сидячими". Например, сидячими являются все ГАМК-ергические синапсы.
Механизм функционирования химического синапса [править
При деполяризации пресинаптической терминали открываются потенциал-чувствительные кальциевые каналы, ионы кальция входят в пресинаптическую терминаль и запускают механизм слияния синаптических пузырьков с мембраной. В результате медиатор выходит в синаптическую щель и присоединяется к белкам-рецепторам постсинаптической мембраны, которые делятся на метаботропные и ионотропные. Первые связаны с G-белком и запускают каскад реакций внутриклеточной передачи сигнала. Вторые связаны с ионными каналами, которые открываются при связывании с ними нейромедиатора, что приводит к изменению мембранного потенциала. Медиатор действует в течение очень короткого времени, после чего разрушается специфическим ферментом. Например, в холинэргических синапсах фермент, разрушающий медиатор в синаптической щели - ацетилхолинэстераза. Одновременно часть медиатора может перемещаться с помощью белков-переносчиков через постсинаптическую мембрану (прямой захват) и в обратном направлении через пресинаптическую мембрану (обратный захват). В ряде случаев медиатор также поглощается соседними клетками нейроглии.
Открыты два механизма высвобождения: с полным слиянием везикулы с плазмалеммой и так называемый "поцеловал и убежал" (англ. kiss-and-run), когда везикула соединяется с мембраной, и из неё в синаптическую щель выходят небольшие молекулы, а крупные остаются в везикуле. Второй механизм, предположительно, быстрее первого, с помощью него происходит синаптическая передача при высоком содержании ионов кальция в синаптической бляшке.
Следствием такой структуры синапса является одностороннее проведение нервного импульса. Существует так называемая синаптическая задержка - время, нужное для передачи нервного импульса. Её длительность составляет около - 0,5 мс.
Так называемый "принцип Дейла" (один нейрон - один медиатор) признан ошибочным. Или, как иногда считают, он уточнён: из одного окончания клетки может выделяться не один, а несколько медиаторов, причём их набор постоянен для данной клетки.
Нервная система человека и животных может быть представлена как система нейронных цепочек, передающих возбуждающие и тормозные сигналы (нервная сеть). Эти элементарные нейронные цепи служат, например, для усиления слабых сигналов, уменьшения слишком интенсивной активности, выделения контрастов, поддержания ритмов или сохранения рабочего состояния нейронов путем регулировки их входов. Такие нейронные цепи построены из стандартных элементов, которые выполняют наиболее часто повторяющиеся операции и могут быть включены в схемы самых разнообразных нервных структур.
Существуют значительные количественные различия нервных сетей у разных видов позвоночных и беспозвоночных. Так, у человека нервная система включает около 10 10 элементов, у примитивных беспозвоночных - около 10 4 нейронов, Однако в строении и функционировании всех нервных систем имеются общие черты. Практически во всех отделах центральной нервной системы обнаружены дивергенция нервных путей, конвергенция нервных путей и различные варианты тормозных связей между элементами нервных цепочек.
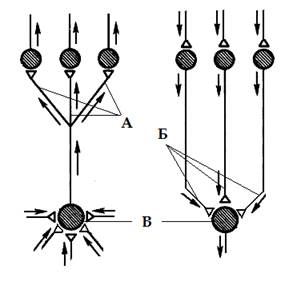
Дивергенция и конвергенция путей. Дивергенция (расхождение) пути (рис 2.А) - возникает в результате контактирования одного нейрона с множеством нейронов более высоких порядков. Так, например, происходит разделение аксона чувствительного нейрона, входящего в спинной мозг, на множество веточек (коллатералей), которые направляются к разным сегментам спинного мозга и в головной мозг, где происходит передача сигнала на вставочные и далее - на моторные нервные клетки. Дивергенция пути сигнала наблюдается так же у вставочных и у эффекторных нейронов.
Дивергенция пути обеспечивает расширение сферы действия сигнала, благодаря ей, информация поступает одновременно к разным участкам ЦНС. Это называют иррадиацией возбуждения (или торможения). Дивергенция настолько обычное явление, что можно говорить о принципе дивергенции в нейронных цепях.
| Конвергенция - это схождение многих нервных путей к одним и тем же нейронам (рис 2.Б). Например, у позвоночных на каждом мотонейроне спинного мозга и ствола головного мозга образуют | Рис 2. Дивергенция (А), конвергенция (Б) и пространственная суммация (В) нервных путей в центральной нервной системе. |
нервные окончания тысячи сенсорных, а также возбуждающих и тормозных вставочных нейронов разных уровней. Мощная конвергенция обнаруживается и на нейронах ретикулярной формации ствола мозга, на многих корковых нейронах у позвоночных и, видимо, на командных нейронах.
Конвергенция многих нервных путей к одному нейрону делает этот нейрон интегратором соответствующих сигналов. Вероятность возбуждения такого нейрона-интегратора зависит не от каждого пришедшего стимула в отдельности, а от суммы и направления стимулов, действующих одновременно, то есть суммы всех синаптических процессов происходящих на его плазматической мембране. Другими словами, вероятность распространения возбуждения через нейрон-интегратор определяется алгебраическим сложением величин возбуждающих и тормозных входов на нем, активных в данный момент. Такое сложение является результатом или пространственной или временной суммации. Пространственная суммация – результат сложения нервных импульсов приходящих одновременно к нейрону через разные синапсы (рис 2.В), временная суммация – сложение приходящих поочередно, через один синапс с небольшими интервалами времени. В обоих случаях нейрон интегратор, называют общим путем для конвергирующих на него нервных сигналов, а если речь идет о мотонейроне, т. е. конечном звене нервного пути к мускулатуре, говорят об общем конечном пути.
Результат суммации заключается в возможности изменения направления распространения возбуждения в ЦНС, (то есть не строго в пределах одной рефлекторной дуги), а значит и в изменении характера ответной реакции организма в ответ на действие раздражителя. Ответ организма, реализуемый в результате, становится более адекватным внешним условиям и состоянию нервной системы. Пример такого выбора ответа можно видеть, если речь идет о конвергенции не на одном нейроне, а на группе нейронов совместно регулирующих общую функцию, что в ЦНС не редкость. Наличие конвергенции множества путей на одной группе мотонейронов лежит в основе феноменов пространственного облегчения и окклюзии.
Пространственное облегчение - это превышение эффекта одновременного действия двух относительно слабых афферентных возбуждающих входов в ЦНС над суммой их раздельных эффектов. Т.е. при раздельном действии афферентных сигналов возбуждение возникает в меньшем числе эфферентных нейронов и эффект оказывается слабее. Феномен объясняется суммацией совместно возникающих ВПСП до критического уровня деполяризации в группе мотонейронов, в которых при раздельной активации входов ВПСП оказывались слишком слабыми для генерации ответа.
Окклюзия - это явление, противоположное пространственному облегчению. В этом случае эффект окажется выше если афферентные сигналы действуют порознь, а при их совместном действии возбуждается меньшая группа мотонейронов. Причина окклюзии состоит в том, что здесь афферентные входы в силу конвергенции частично связаны с одними и теми же мотонейронами, и каждый может возбуждать их, как и оба входа вместе.
Таким образом, если эффект нескольких стимулов, поступающих одновременно или в быстрой последовательности будет выше, чем сумма эффектов отдельных стимулов то это явление называется облегчением; если же эффект на сочетание стимулов меньше, чем сумма ответов на отдельные стимулы, то такое явление - окклюзия.
Такое явление следует учитывать, например, при тренировке различных функцинальных показателей скелетных мышц.
Простые тормозные и усиливающие цепи.
Тормозные цепи, виды торможения. Торможение, как и возбуждение, - активный процесс, оно возникает в результате сложных физико-химических изменений в тканях. Благодаря процессу торможения достигается ограничение распространения возбуждения в ЦНС и обеспечивается координация рефлекторных актов, внешне этот процесс проявляется ослаблением функции какого-либо органа.
К настоящему времени анализ тормозных явлений в ЦНС позволил выделить две формы разновидности постсинаптическое и пресинаптическое торможение.
Постсинаптическое торможение развивается не постсинаптических мембранах межнейронных синапсов и связано с гиперполяризацией постсинаптической мембраны под влиянием медиаторов, которые выделяются при возбуждении специальных тормозных нейронов. При этом, локально возникающая на постсинаптической мембране гиперполяризация - тормозной постсинаптический потенциал (ТПСП) – затрудняет электротоническое распространение возбуждающих постсинаптических потенциалов (ВПСП) от других синапсов, к аксонному холмику. В результате в зоне аксонного холмика не происходит выведение мембранного потенциала на критический уровень. Потенциал действия не образуется, нейрон не возбуждается.
Постсинаптическое торможение активно используется в нейронных сетях, и в зависимости от вариантов связывания нейронов друг с другом выделяют несколько его видов: реципрокное (прямое), параллельное, возвратное, латеральное (рис.3)
Реципрокное торможение (рис 3.А) – это взаимное (сопряженное) торможение центров антагонистических рефлексов, обеспечивающее координацию этих рефлексов. Классический пример реципрокного торможения - это торможение мотонейронов мышц-антагонистов у позвоночных. Торможение осуществляется с помощью специальных тормозных вставочных нейронов. При активации путей, возбуждающих, например, мотонейроны мышц-сгибателей, мотонейроны мышц-разгибателей тормозятся импульсами вставочных клеток.
Возвратное торможение (рис. 3.Б) - это торможение нейронов собственными импульсами, поступающими по возвратным коллатералям к тормозным клеткам. Возвратное торможение наблюдается, например, в мотонейронах спинного мозга позвоночных. Эти клетки отдают возвратные коллатерали в мозг к тормозным вставочным клеткам Реншоу, которые имеют синапсы на этих же мотонейронах. Торможение обеспечивает ограничение ритма мотонейронов, позволяющее чередовать сокращение и расслабление скелетной мышцы, что важно для нормальной работы двигательного аппарата.
Такую же роль играет возвратное торможение и в других нервных сетях.
Параллельное торможение (рис. 3.В) – играет сходную с возвратным роль, но в этом случае возбуждение блокирует само себя, посылая тормозной сигнал на нейрон который одновременно и активирует.
Это возможно, если возбуждающий импульс сам не должен вызвать возбуждения на нейроне-мишени, но его роль важна при пространственной суммации, в комбинации с другими сигналами.
Латеральное торможение (рис. 3.Г) – это торможение нервных клеток, расположенных по соседству с активной, которое этой клеткой и инициируется. При этом вокруг возбужденного нейрона возникает зона, в которой развивается очень глубокое торможение.
Латеральное торможение наблюдается, например, в конкурирующих сенсорных каналах связи. Оно наблюдается у соседних элементов сетчатки позвоночных, а также в их зрительных, слуховых и других сенсорных центрах. Во всех случаях латеральное торможение обеспечивает контраст, т. е. выделение существенных сигналов или их границ из фона.
Пресинаптическое торможение развивается в аксо-аксональных синапсах, образованных на пресинаптических окончаниях нейрона.
В основе пресинаптического торможения лежит развитие медленной и длительной деполяризации пресинаптического окончания, что и приводит к развитию торможения. В деполяризованном участке нарушается процесс распространения возбуждения и поступающие к нему импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуде, не обеспечивают выделения достаточного количества медиатора – нейрон не возбуждается.
Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные нейроны, аксоны которых и образуют синапсы на пресинаптических окончаниях аксона-мишени.
Разновидности пресинаптического торможения изучены недостаточно, вероятно они те же, что и для постсинаптического торможения. Точно известно о наличии параллельного и латерального пресинаптического торможения (рис. 4).
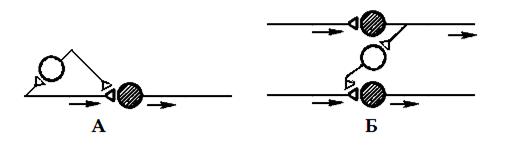
Рис. 4. Разновидности пресинаптического торможения: А – параллельное, Б – латеральное. Темные нейроны – возбуждающие, светлые – тормозные.
В реальной действительности взаимоотношения возбуждающих и тормозных нейронов значительно сложнее, чем представлено на рисунках, тем не менее, все варианты пре- и постсинаптического торможения можно объединить в две группы. Во-первых, когда блокируется собственный путь самим распространяющимся возбуждением с помощью вставочных тормозных клеток (параллельное и возвратное торможение), во-вторых, когда блокируются другие нервные элементы под влиянием импульсов от соседних возбуждающих нейронов с включением тормозных клеток (латеральное и прямое торможение).
Кроме того, тормозные клетки сами могут быть заторможены другими тормозными нейронами, это может облегчить распространение возбуждения.
Роль процесса торможения.
- Оба известных вида торможения со всеми их разновидностями выполняют, прежде всего, охранительную роль. Отсутствие торможения привело бы к истощению медиаторов в аксонах нейронов, утомлению, истощению и прекращению деятельности ЦНС.
- Торможение играет важную роль в обработке поступающей в ЦНС информации. Особенно ярко выражена эта роль у пресинаптического торможения. Оно более точно регулирует процесс возбуждения, поскольку этим торможением могут быть полностью заблокированы отдельные нервные волокна. К одному возбуждающему нейрону могут приходить сотни и тысячи различных импульсов по разным путям, но число дошедших до нейрона импульсов определяется пресинаптическим торможением.
- Поскольку блокада торможения ведет к широкой иррадиации возбуждения и судорогам, следует признать, что торможение является важным фактором обеспечения координационной деятельности ЦНС.
Усиливающие цепи и механизмы усиления. Нейронные сети имеют не только тормозные механизмы, препятствующие распространению возбуждения, но и системы, усиливающие приходящий к ним сигнал. Рассмотрим некоторые из них.
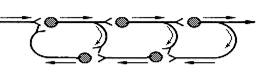
Самовозбуждающиеся нервные цепи (цепи с положительной обратной связью) (рис.5). Некоторые данные свидетельствуют о том, что в мозгу животных и человека существуют замкнутые самовозбуждающиеся цепочки нейронов, в которых нейроны соединены синапсами возбуждающего действия. Возникнув в ответ на внешний сигнал, возбуждение в такой цепочке циркулирует, иначе реверберирует, до тех пор, пока или какой-либо внешний тормоз не выключит одно из звеньев цепи, или в ней не наступит утомление. Выходные пути от такой цепочки (ответвляющиеся по коллатералям аксонов нервных клеток - участников цепи) во время работы передают равномерный поток импульсов, создающий ту или иную настройку в нервных клетках-мишенях. Ее функции могут состоять в том, чтобы обеспечивать длительное поддержание индуцированной однажды активности.
сигнал, который включил в ней циркуляцию (реверберацию) импульсов. Считают, что это возможный механизм (или один из механизмов) краткосрочной памяти, однако этому практически нет экспериментальных доказательств.
Синаптическая потенциация — увеличение амплитуды постсинаптического потенциала, если интервал между последовательным возникновением потенциалов действия в пресинаптической мембране невелик, то есть происходит частая и ритмическая активация синапса. Явление потенциации связывают с накоплением ионов кальция в пресинаптическом окончании, который дополнительно вбрасывается туда при каждом новом стимуле и не успевает полностью удаляться между частыми стимулами. Вследствие этого, каждый новый пресинаптический потенциал вызывает высвобождение большего числа квантов медиатора.
Такую же природу имеет и посттетаническая потенциация. В этом случае увеличение числа квантов медиатора, высвобождаемых нервным импульсом, после предшествующего ритмического раздражения приводит к увеличению синаптической реакции нейрона на одиночное раздражение пресинаптических путей. Посттетаническая потенциация может длиться от нескольких минут до нескольких часов в различных структурах мозга. Предполагают, что постсинаптическая потенциация играет важную роль в пластических перестройках функций синапсов, и лежит в основе механизмов организации условных рефлексов и памяти. Например, особенно длительная посттетаническая потенциация обнаружена в гиппокампе – структуре, которая, играет важную роль в явлениях памяти и научения.
Ритмическая стимуляция может приводить и к снижению активности синапсов. Процесс снижения постсинаптических потенциалов во время или по окончании тетанической стимуляции по сравнению с исходной амплитудой называется синаптической депрессией; по аналогии с потенциацией, различают тетаническую и посттетаническую депрессию. Возможно, синаптическая депрессия имеет место во многих участках нервной системы и является нейронным коррелятом привыкания (габитуации). У беспозвоночных габитуация простых поведенческих реакций прямо соответствует депрессии участвующих синапсов; то же самое относится и к флексорному рефлексу у кошки. Таким образом, синаптическая депрессия, так же как синаптическая потенциация, составляет элементарный процесс научения.

Нервная система
Раздражимость или чувствительность – характерная черта всех живых организмов, означающая их способность реагировать на сигналы или раздражители.
Сигнал воспринимается рецептором и передается с помощью нервов и (или) гормонов к эффектору, который осуществляет специфическую реакцию или ответ.
Животные имеют две взаимосвязанные системы координации функций – нервную и гуморальную (см. таблицу).
Нервная регуляция
Гуморальная регуляция
Электрическое и химическое проведение (нервные импульсы и нейромедиаторы в синапсах)
Химическое проведение (гормоны) по КС
Быстрое проведение и ответ
Более медленное проведение и отстроченный ответ (исключение - адреналин)
В основном кратковременные изменения
В основном долговременные изменения
Специфический путь распространения сигнала
Неспецифический путь сигнала (с кровью по всему телу)к специфической мишени
Ответ часто узко локализован (например, один мускул)
Ответ может быть крайне генерализованным (например, рост)
Нервная система состоит из высокоспециализированных клеток со следующими функциями:
- восприятие сигналов – рецепторы;
- преобразование сигналов в электрические импульсы (трансдукция);
- проведение импульсов к другим специализированным клеткам – эффекторам, которые получив сигнал, дают ответ;
Связь между рецепторами и эффекторами осуществляют нейроны .
Нейрон – это структурно – функциональная единица НС.

Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. Нейрон имеет сложное строение и узкую специализацию. Нервная клетка содержит ядро, тело клетки и отростки (аксоны и дендриты).
В головном мозге человека насчитывается около 90—95 миллиардов нейронов. Нейроны могут соединяться друг с другом, образуя биологические нейронные сети.
Нейроны разделяют на рецепторные, эффекторные и вставочные.
Тело нейрона: ядро (с большим количеством ядерных пор) и органеллы (ЭПС, рибосомы, аппарат Гольджи, микротрубочки), а также из отростков (дендриты и аксоны).
Нейроглия – совокупность вспомогательных клеток НС; составляет 40% общего объема ЦНС.
- Аксон – длинный отросток нейрона; проводит импульс от тела клетки; покрыт миелиновой оболочкой (образует белое вещество мозга)
- Дендриты - короткие и сильно разветвлённые отростки нейрона; проводит импульс к телу клетки; не имеют оболочки

Важно! Нейрон может иметь несколько дендритов и обычно только один аксон.
Важно! Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами.
- чувствительные – передают возбуждение от органов чувств в спинной и головной мозг
- двигательные – передают возбуждение от головного и спинного мозга к мышцам и внутренним органам
- вставочные – осуществляют связь между чувствительными и двигательным нейронами, в спинном и головном мозге
Нервные отростки образуют нервные волокна.
Пучки нервных волокон образуют нервы.
Нервы – чувствительные (образованы дендритами), двигательные (образованы аксонами), смешанные (большинство нервов).
Синапс – это специализированный функциональный контакт между двумя возбудимыми клетками, служащий для передачи возбуждения

У нейронов синапс находится между аксоном одной клетки и дендритом другой; при этом физического контакта не происходит – они разделены пространством - синаптической щель.
Нервная система:
- периферическая (нервы и нервные узлы) – соматическая и автономная
- центральная (головной и спинной мозг)
В зависимости от характера иннервации НС:
- Соматическая – управляет деятельностью скелетной мускулатуры, подчиняется воле человека
- Вегетативная (автономная) – управляет деятельностью внутренних органов, желез, гладкой мускулатуры, не подчиняется воле человека
Соматическая нервная система – часть нервной системы человека, представляющая собой совокупность чувствительных и двигательных нервных волокон, иннервирующих мышцы (у позвоночных — скелетные), кожу, суставы.
Она представляет часть периферической нервной системы, которая занимается доставкой моторной (двигательной) и сенсорной (чувственной) информации до центральной нервной системы и обратно. Эта система состоит из нервов, прикрепленных к коже, органам чувств и всем мышцам скелета.
- спинномозговые нервы – 31 пара; связаны со спинным мозгом; содержат как двигательные, так и сенсорные нейроны, поэтому смешанные;
- черепномозговые нервы – 12 пар; отходят от головного мозга, иннервируют рецепторы головы (за исключением блуждающего нерва – иннервирует сердце, дыхание, пищеварительный тракт); бывают сенсорными, моторными (двигательными) и смешанными
Рефлекс – это быстрый автоматический ответ на раздражитель, осуществляемый без осознанного контроля головного мозга.
Рефлекторная дуга – путь, проходимый нервными импульсами от рецептора до рабочего органа.
- в ЦНС – по чувствительному пути;
- от ЦНС – к рабочему органу – по двигательному пути
- рецептор (окончание дендрита чувствительного нейрона) – воспринимает раздражение
- чувствительное (центростремительное) нервное волокно – передает возбуждение от рецептора к ЦНС
- нервный центр – группа вставочных нейронов, расположены на разных уровнях ЦНС; передает нервные импульсы с чувствительных нейронов на двигательные
- двигательное (центробежное) нервное волокно – передает возбуждение от ЦНС к исполнительному органу

Простая рефлекторная дуга: два нейрона – чувствительный и двигательный (пример – коленный рефлекс)
Сложная рефлекторная дуга: три нейрона – чувствительный, вставочный, двигательный (благодаря вставочным нейронам происходит обратная связь между рабочим органом и ЦНС, что позволяет вносить изменения в работу исполнительных органов)
Вегетативная (автономная) нервная система – управляет деятельностью внутренних органов, желез, гладкой мускулатуры, не подчиняется воле человека.
Делится на симпатическую и парасимпатическую.

Обе состоят из вегетативных ядер (скопления нейронов, лежащих в спинном и головном мозге), вегетативных узлов (скопления нейронов, нейронов, за пределами НС), нервных окончаний (в стенках рабочих органов)
Путь от центра до иннервируемого органа состоит из двух нейронов (в соматической - один).
Место выхода из ЦНС
От спинного мозга – в шейный, поясничный, грудной отделы
От ствола головного мозга и ствола крестцового отдела спинного мозга
Местоположение нервного узла (ганглия)
По обе стороны спинного мозга, за исключением нервных сплетений (непосредственно в этих сплетениях)
В иннервируемых органах или вблизи них
Медиаторы рефлекторной дуги
В предузловом волокне –
в послеузловом - норадреналин
В обоих волокнах - ацетилхолин
Названия основных узлов или нервов
Солнечное, легочное, сердечное сплетения, брыжеечный узел
Общие эффекты симпатической и парасимпатической НС на органы:
- Симпатическая НС – расширяет зрачки, угнетает слюноотделение, повышает частоту сокращений, расширяет сосуды сердца, расширяет бронхи, усиливает вентиляцию легких, угнетает перистальтику кишечника, угнетает секрецию пищеварительных соков усиливает потоотделение, удаляет с мочой лишний сахар; общий эффект – возбуждающий, повышает интенсивность обмена, снижает порог чувствительности; активизирует во время опасности, стресса, контролирует реакции на стресс
- Парасимпатическая НС – сужает зрачки, стимулирует слезотечение, уменьшает частоту сердечных сокращений, поддерживает тонус артериол кишечника, скелетных мышц, снижает кровяное давление, уменьшает вентиляцию легких, усиливает перистальтику кишечника, расширяет артериолы в коже лица, увеличивает выделение с мочой хлоридов; общий эффект – тормозящий, снижает или не влияет на интенсивность обмена, восстанавливает порог чувствительности; доминирует в состоянии покоя, контролирует функции в повседневных условиях
Центральная нервная система (ЦНС) – обеспечивает взаимосвязь всех частей НС и их координированную работу
У позвоночных ЦНС развивается из эктодермы (наружного зародышевого листка)
ЦНС – 3 оболочки:
- твердая мозговая (dura mater) - снаружи;
- мягкая мозговая оболочка (pia mater) – прилегает непосредственно к мозгу.
Головной мозг расположен в мозговом отделе черепа; содержит
- белое вещество - проводящие пути между головным мозгом и спинным, между отделами головного мозга
- серое вещество - в виде ядер внутри белого вещества; кора покрывающая большие полушария и мозжечок
Масса головного мозга – 1400-1600 грамм.

5 отделов:
- продолговатый мозг– продолжение спинного мозга; центры пищеварения, дыхания, сердечной деятельности, рвота, кашель, чихание, глотание, слюноотделение, проводящая функция
- задний мозг – состоит из варолиевого моста и мозжечка; варолиев мост связывает мозжечок и продолговатый мозг с большими полушариями; мозжечок регулирует двигательные акты (равновесие, координация движений, поддержание позы)
- промежуточный мозг– регуляция сложных двигательных рефлексов; координация работы внутренних органов; осуществление гуморальной регуляции;
- средний мозг – поддержание тонуса мыщц, ориентировочные, сторожевые, оборонительные рефлексы на зрительные и звуковые раздражители;
- передний мозг (большие полушария) – осуществление психической деятельности (память, речь, мышление).
Промежуточный мозг включает таламус, гипоталамус, эпиталамус
Таламус – подкорковый центр всех видов чувствительности (кроме обонятельного), регулирует внешнее проявление эмоций (мимика, жесты, изменение пульса, дыхания)
Гипоталамус – центры вегетативной НС, обеспечивают постоянство внутренней среды, регулируют обмен веществ, температуру тела, чувство жажды, голода, насыщения, сна, бодрствования; гипоталамус контролирует работу гипофиза
Эпиталамус – участие в работе обонятельного анализатора
Передний мозг имеет два больших полушария: левое и правое
- Серое вещество (кора) находится сверху полушарий, белое – внутри
- Белое вещество – это проводящие пути полушарий; среди него – ядра серого вещества (подкорковые структуры)
Кора больших полушарий – слой серого вещества, 2-4 мм в толщину; имеет многочисленные складки, извилины
Каждое полушарие разделено бороздами на доли:
- лобная – вкусовая, обонятельная, двигательная, кожно- мускульная зоны;
- теменная – двигательная, кожно- мускульная зоны;
- височная – слуховая зона;
- затылочная – зрительная зона.
Важно! Каждое полушарие отвечает за противоположную сторону тела.
- Левое полушарие – аналитическое; отвечает за абстрактное мышление, письменную и устную речь;
- Правое полушарие – синтетическое; отвечает за образное мышление.
Спинной мозг расположен в костном позвоночном канале; имеет вид белого шнура, длина 1м; на передней и задней сторонах есть глубокие продольные борозды
В самом центре спинного мозга – центральный канал, заполненный спинномозговой жидкостью.
Канал окружен серым веществом (имеет вид бабочки), который окружен белым веществом.
- В белом веществе – восходящие (аксоны нейронов спинного мозга) и нисходящие пути (аксоны нейронов головного мозга)
- Серое вещество напоминает контур бабочки, имеет три вида рогов.
- передние рога – в них расположены двигательные нейроны (мотонейроны) – их аксоны иннервируют скелетные мышцы
- задние рога – содержат вставочные нейроны – связывают чувствительные и двигательные нейроны
- боковые рога – содержат вегетативные нейроны – их аксоны идут на периферию к вегетативным узлам
Спинной мозг – 31 сегмент; от каждого сегмента отходит 1 пара смешанных спинномозговых нервов, имеющих по паре корешков:
- передний (аксоны двигательных нейронов);
- задний (аксоны чувствительных нейронов.
Функции спинного мозга:
- рефлекторная – осуществление простых рефлексов (сосудодвигательных, дыхательных, дефекации, мочеиспускания, половых);
- проводниковая – проводит нервные импульсы от и к головному мозгу.

Повреждение спинного мозга приводит к нарушению проводниковых функций, вследствие чего – паралич.
Читайте также:
