
Что такое пролонгирование возбуждения в нервном центре
центре?
103.Что такое пролонгирование возбдужения в нервном центре?
Этим термином обозначается явление, при котором рефлекторный ответ сохраняется после прекращения действия раздражителя, т.е раздражение прекратилось, а, например, мышца продолжает сокращаться.
Существует 2 механизма этого феномена:
1. Следовая деполяризация мембраны нейрона, возникающая еще на фоне сохраняющегося ВПСП, создает условия для ритмической генерации потенциалов действия. Это кратковременное последствие.
2. Циркуляция возбуждения по замкнутым цепям нейронов в рефлекторном центре (принцип Лоренто-де-Но).
До прихода тормозного импульса или утомления одного из синапсов возбуждения может достаточно долго циркулировать по этим цепям. Бала выдвинута гипотеза о тем, что кратковременная память обусловлена реверберацией возбуждения в цепях положительной обратной связи.
104. Что такое феномен облегчения и каков его механизм?
Явление облегчения. Отдельные нейроны являются общими для каких-то двух рефлексов. При изолированной реализации одного из рефлексов эти нейроны не дают ответной реакции, так как раздражение для них оказывается подпороговым. При совместной реализации двух рефлексов их подпороговые эффекты суммируются и достигают пороговой величины. В результате суммации ответная реакция оказывается больше простой суммы изолированных эффектов.
Пул нейронов, расположенных в передних рогах спинного мозга состоит из высоковозбудимых (1,2) и низковозбудимых неронов (3). При раздражении только 1 афферентного нервного волокна в состояние возбуждения приходит нейрон 1, а в нейроне 3 возникает деполяризация, не достигающая критического уровня потенциала (т.е. его возбудимость увеличивается). При этом на выходе регистрируется сигнал равный 120 мВ.
При раздражении только II афферентного волокна возбуждается нейрон 2, а в нейроне 3 возникает увеличение возбудимости и на выходе регистрируется потенциал, например, равный 120 мВ. При одновременном раздражении I и II афферентных волокон в состояние возбуждения переходят нейроны 1,2,3. При данном воздействии регистрируется суммарный потенциал 320 мВ, т.е. не равный арифметической сумме отдельных потенциалов (120+120=240 мВ). Это обусловлено тем, что при одновременном раздражении волокон I и II дополнительно включается нейрон 3. Таким образом облегчение – это усиление поступающего сигнала.

временное
105. Что такое суммация возбуждения в нервном центре и ее механизм?
Особенностью ЦНС является то, что, как правило одни потенциал действия не вызывает возбуждение эффекторного нейрона. Только на ритмический раздражитель одного рецептора или одновременное раздражение нескольких нейронов возникает возбуждение эффекторного нейрона.
В зависимости от ситуации суммация бывает: 1) временная или 2) пространственная.
В случае временной суммации происходит суммирование квантов медиатора на постсинаптической мембране. Если в область постсинаптической мембраны ввести электрод и соединить его с усилителем постоянного тока и осциллографом, то можно наблюдать, что на одиночный раздражитель возбуждение нейрона не возникает, но на постсинаптической мембране возникает небольшой по амплитуде ВПСП.
С точки зрения возникновения возбуждения данный ВПСП является недостаточным, так как не может достигнуть критического уровня деполяризации. Если последующий импульс накладывается на первый, то есть суммируется и общий ВПСП достигает критического уровня. Это приводит к развитию потенциала действия. Таким образом 1) временная суммация – это суммация квантов медиатора, в первую очередь, в области аксонального холмика нейрона 2) временная суммация ВПСП обусловлена тем, что ВПСП продолжается дольше, чем рефрактерный период аксона, из окончания которого выделяется медиатор.
Пространственная суммация - это явление, когда раздельная стимуляция каждого из двух аксонов вызывает подпороговый ВПСП, однако при одновременной стимуляции обоих аксонов возникает потенциал действия, т.е. процесс, который не может быть обеспечен одиночным ВПСП.
Пространственная суммация может быть линейная и нелинейная. Если стимуляция будет пропорциональная каждому возбуждающему постсинаптическому потенциалу (ВПСП), то она называется линейной. Подобная ситуация возникает, когда возбуждающие синапсы на мембране одного нейрона удалены друг от друга на достаточном удалении.
Если синапсы находятся близко друг возле друга, силовые линии соседних ВПСП уменьшают их ВПСП и общий постсинаптический потенциал не будет пропорционален каждому ВПСП.
Пространственная суммация лежит в основе эффекта облегчения и окклюзии.
106. Что такое торможение? Классификация торможения.
Если на зрительный бугор поместить кристаллы соли (NaCl), то возникает торможение – удлинение времени рефлекса (замедление времени выдёргивания лапки из кислоты). Это наблюдение позволило И.М.Сеченову высказать мнение о явлении торможения в ЦНС. В последующем такой вид торможения получил название сеченовского торможения, или центральное торможение. Периферическое торможение было открыто ещё в 1845 году братьями Вебер (торможение деятельности сердца при раздражении блуждающих нервов).
Торможение – это активный процесс, связанный с возбуждением и ограничивающий его. Торможение это такой же врождённый процесс как и возбуждение, но общим для всех видов торможения является отсутствие способности к распространению по мембране нейрона и его отросткам. Различают два принципиально различных механизма торможения в ЦНС (рис. 20): первичное и вторичное. Первичное торможение развивается в клетках, примыкающих к тормозному нейрону. Оно инициировано возбуждением специальных тормозных нейронов, которые выделяют тормозные медиаторы. Вторичное торможение возникает в тех же нейронах, которые генерируют возбуждение.
107. Что такое первичное торможение?
Первичное торможение развивается в клетках, примыкающих к тормозному нейрону. Оно инициировано возбуждением специальных тормозных нейронов, которые выделяют тормозные медиаторы.
Первичное – возникает в специальных тормозных структурах под влиянием процесса возбуждения и проявляется подавлением другого возбуждения, развивающегося в соседних с этой структурой клетках. Следовательно, для тормозимой клетки этот процесс является первичным, т.е. без предварительного возбуждения.
| Постсинаптическое торможение (постторможение) | Пресинаптическое торможение |
| Этот вид торможения связан со специальными структурами (синапсами), которые выделяют тормозные медиаторы (ГАМК, Глицин) |
108. Что такое вторичное торможение и его виды?
Вторичное – это торможение возникает в той же самой клетке, в которой наблюдалось первичное возбуждение, т.е. торможение – это результат возбуждения.
Ко вторичному торможению относится пессимальное торможение и торможение вслед за возбуждением.
Пессимальное торможение – заключается в том, что в единицу времени к нейрону поступает очень большое количество потенциалов действия. В результате этого может происходить инактивация потенциал-зависимых натриевых каналов или десинситизация (потеря чувствительности) рецепторов, расположенных на постсинаптической структуре (десинситизация рецепторов обусловлена тем, что при большой частоте поступления медиатора он не успевает разрушаться и тем самым освобождать рецепторы для взаимодействия с новыми порциями медиатора).
Торможение вслед за возбуждением обусловлено тем, что в некоторых нейронах после потенциала действия развивается длительная следовая гиперполяризация, снижающая возбудимость клетки на долгое время.
Нейроны ЦНС для осуществления сложных и многообразных функций объединяются в нервные центры. Нервный центр - это совокупность нейронов, принимающих участие в осуществлении конкретного рефлекса (мигания, глотания, кашля и т. д.) . В целом организме при формировании сложных адаптивных процессов происходит функциональное объединение нейронов, расположенных на различных уровнях ЦНС. Такое объединение (нервный центр в широком смысле слова) позволяет осуществлять наиболее адекватное для конкретных условий осуществления рефлекторной деятельности.
Нервные центры обладают рядом характерных функциональных свойств, обусловленных объединением нейронов в нейронные сети и наличием межнейрональных синапсов.
Основные из этих свойств:
1. Возбуждение в нервных центрах распространяется односторонне - от рецептора к эффектору, что обусловливается свойством химических синапсов односторонне проводить возбуждение от пресинаптической мембраны к постсинаптической.
2. Возбуждение в нервных центрах проводится медленнее, чем по нервному волокну. Это обусловлено замедленным проведением возбуждения через синапс (синаптическая задержка) .
3. В нервных центрах осуществляется суммация возбуждений. Различают два вида суммации:
• временная или последовательная, если импульсы возбуждения приходят к нейрону по одному и тему же пути через один синапс с интервалом меньше, чем время полной реполяризации постсинаптической мембраны. В этих условиях ВПСП на постсинаптической мембране суммируются и доводят ее деполяризацию до уровня, достаточного для генерации нейроном потенциала действия;
• пространственная или одновременная - наблюдается в том случае, когда импульсы возбуждения поступают к нейрону одновременно через разные синапсы (рис. 10).
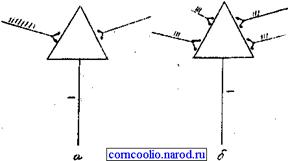
Рис. 10. Схема временной (а) и пространственной (б) суммации возбуждений в центральной нервной системе.
4. Трансформация ритма возбуждения - изменение количества импульсов возбуждения, выходящих из нервного центра, по сравнению с числом импульсов, приходящих к нему. Различают два вида трансформации:
• понижающая трансформация, в основе которой, в основном, лежит явление суммации возбуждений (пространственной и временной), когда в ответ на несколько возбуждений, пришедших к нервной клетке, в последней возникает только одно возбуждение;
• повышающая трансформация, в ее основе лежат механизмы умножения (мультипликации), способные резко увеличить количество импульсов возбуждения (рис. 11).
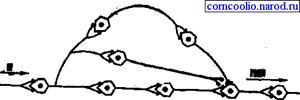
Рис. 11. Схема умножения (мультипликации) возбуждения в центральной нервной
5. Рефлекторное последействие - заключается в том, что рефлекторная реакция заканчивается позже прекращения действия раздражителя. Это явление обусловлено двумя причинами:
• длительной следовой деполяризацией мембраны нейрона, на фоне которой могут возникать несколько потенциалов действия, обеспечивающих кратковременное рефлекторное последействие;
• пролонгированием выхода возбуждения к эффектору в результате циркуляции (реверберации) возбуждения в нейронной сети типа "нейронной ловушки" (рис. 12). Возбуждение, попадая в такую сеть, может длительное время циркулировать в ней, обеспечивая длительное рефлекторное последействие. Возбуждение в такой цепочке может циркулировать до тех пор, пока какое-либо внешнее воздействие затормозит этот процесс или в ней наступит утомление.
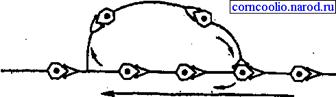
Рис. 12. Схема пролонгирования возбуждения (нейронной ловушки) в центральной нервной системе.
6. Нервные центры, как и синапсы, обладают высокой чувствительностью к недостатку кислорода.
7. Нервные центры, как и синапсы, обладают высокой чувствительностью к действию различных химических веществ, особенно ядов. На одном нейроне могут располагаться синапсы, обладающие различной чувствительностью к различным химическим веществам.
Поэтому можно подобрать такие химические вещества, которые избирательно будут блокировать одни синапсы, оставляя другие в рабочем состоянии. Это делает возможным корректировать состояния и реакции как здорового, так и больного организма.
8. Нервные центры, как и синапсы, обладают быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми.
9. Нервные центры, как и синапсы, обладают низкой лабильностью.
10. В нервных центрах легко возникает процесс торможения.
11. Нервные центры обладают тонусом, который выражается в том, что даже при отсутствии специальных раздражении, они постоянно посылают импульсы к рабочим органам.
12. Нервные центры, как и синапсы, обладают низкой аккомодационной способностью, т. е. они способны реагировать на раздражающие факторы, медленно нарастающие по силе.
13. Нервные центры обладают пластичностью - способностью изменять собственное функциональное назначение и расширять свои функциональные возможности.
14. Посттетаническая потенциация - явление усиления рефлекторного ответа после длительного ритмического раздражения нервного центра. Оно обусловлено сохранением определенного уровня ВПСП на нейронах центра, что облегчает проведение последующих возбуждений через синапсы.
Последействие и пролонгирование возбуждения — связаны с длительными следовыми потенциалами в нейронах, улучшением синаптического проведения, наличием кольцевых нейронных цепей и реверберацией возбуждения.
Трансформация ритма возбуждений
Трансформация ритма возбуждений, т.е. увеличение или уменьшение частоты нервных импульсов и эфферентных проводниках (на выходе) по сравнению с частотой афферентной импульсации (на входе центра), что связано с механизмом синаптической передачи (трансформация ритма как свойство синапса) и интегративной деятельностью нейронов;
Спонтанная (фоновая) электрическая активность
Спонтанная (фоновая) электрическая активность — периодическое генерирование импульсов возбуждения (потенциалов действия) нервными клетками центра в состоянии покоя, т.е. без специфичес кого раздражения рецептивного поля рефлекса.
Тонус нервного центра
Тонус нервного центра — состояние некоторого уровня активности нейронов, обеспечивающей их готовность к рефлекторной деятельности и проявляющейся в постоянной эфферентной импульсации низкой частоты к органам-эффекторам.
Пластичность нервных центров
Пластичность нервных центров — способность перестраивать функциональные свойства для более эффективной регуляции функций, осуществления новых, ранее несвойственных этому центру рефлексов или восстановления функций после повреждения части нейронов центра.
Утомление нервных центров
Утомление нервных центров — снижение эффективности их деятельности в виде повышения порогов возбуждения, связанное с утомлением синапсов и метаболическими сдвигами типа энергетического истощения в нервных клетках.
3. Иррадиация и конвергенция возбуждения. Облегчение и окклюзия.
Иррадиация возбуждения
Это активное распространение возбуждения в ЦНС, особенно при сильном и длительном раздражении. Возможность иррадиации в ЦНС обусловлена наличием в ней многочисленных ответвлений отростков (аксонов, дендритов) нервных клеток и цепей интернейронов, которые соединяют между собой различные нервные центры. Важную роль в иррадиации возбуждения в структурах мозга играет ретикулярная формация.
Усиление раздражения или повышение возбудимости ЦНС сопровождается усилением иррадиации возбуждения в ней. Тормозные нейроны и синапсы препятствуют иррадиации возбуждения или ограничивают ее.
Конвергенция возбуждения
На каждом из нейронов ЦНС конвергирует (сходятся) различные афферентные волокна. Таких афферентных входов для большинства нейронов много десятков и даже тысяч. Так, на мотонейронах заканчиваются в среднем 6000 коллатералей аксонов, которые поступают от периферических рецепторов и различных структур мозга, образуя возбуждающие и тормозные синапсы.
Свойство облегчения проведения и окклюзии нервного импульса — результат конвергенции (схождения) нервных импульсов от разных аксонов к одной нервной клетке. Для генерации нервного импульса может быть недостаточно возбуждения, поступающего к нервной клетке по отростку одного аксона. В этом случае возбуждение от другого аксона, поступающее к той же нервной клетке, облегчает генерацию нервного импульса.
4. Утомление нервных центров. Тонус нервных центров. Пластичность нервных центров.
Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением. Тонус определяется тем, что в покое в отсутствие специальных внешних раздражений определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки. Пластичность - функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
5. Временная и пространственная суммация постсинаптических потенциалов.
Пространственная суммация - суммация в результате действия нескольких возбуждающих постсинаптических потенциалов , возникших одновременно в разных синапсах одного и того же нейрона . Временная суммация - часто повторяющееся высвобождение медиатора из синаптических пузырьков одной и той же синаптической бляшки под действием интенсивного стимула, вызывающего отдельные возбуждающие постсинаптические потенциалы , которые следуют так часто один за другим во времени, что их эффекты суммируются и вызывают в постсинаптическом нейроне потенциал действия .
6. Виды торможения в ЦНС. Постсинаптическое торможение. Тормозные медиаторы.
Пресинаптическое торможение, как свидетельствует название, локализуется в пресинаптических элементах и связано с угнетением проведения нервных импульсов в аксональных (пресинаптических) окончаниях.
Постсинаптическое торможение (ГПСП) обусловлено выделением пресинаптическим окончанием аксона тормозного медиатора, который снижает или тормозит возбудимость мембран сомы и дендритов нервной клетки, с которой он контактирует. Оно связано с существованием тормозных нейронов, аксоны которых образуют на соме и дендритах клеток нервных окончаний, выделяя тормозные медиаторы - ГАМК и глицин. Под влиянием этих медиаторов возникает торможение возбуждающих нейронов.
Постсинаптическое торможение локальное. Развивается оно градуально, способное к суммации, не оставляет после себя рефрактерности. Является более оперативным, четко адресованным и универсальным тормозным механизмом.
7. Тормозные постсинаптические потенциалы (ТПСП). Пресинаптическое торможение. То́рмозный постсинапти́ческий потенциа́л — это разновидность постсинаптического потенциала, которая приводит к тому, что активность постсинаптического нейрона понижается, и менее вероятным становится возникновение потенциала действия. Пресинаптическое торможение - это уменьшение или прекращение высвобождения медиатора из пресинаптических нервных окончаний. При этом не происходит генерации тормозного постсинаптического потенциала .
8. Возвратное торможение. Роль клеток Реншоу.
Возвратноеторможение. Оно реализуется через тормозные клетки Реншоу, открытые в спинном мозге. Аксоны мотонейронов передних рогов отдают коллатераль на тормозной нейрон Реншоу, аксоны которого возвращаются на тот же мотонейрон, образуя на нем тормозные синапсы. Таким образом формируется контур с отрицательной обратной связью, позволяющий стабилизировать частоту разрядов мотонейрона.
Одна из самых важных функций клеток Реншоу, общих для всех типов мотонейронов, — это стабилизация частоты их работы при поддержании позы или удержании нагрузки. То есть КР сглаживают колебания на выходе МН. Когда входной поток к МН возрастает, МН импульсируют сильнее, частота работы КР увеличивается и они притормаживают МН.
9. Латеральное торможение как один из видов возвратного торможения.
Латеральное — тормозная клетка тормозит расположенные рядом нейроны. Подобные явления развиваются между биполярными и ганглиозными клетками сетчатки, что создаёт условия для более четкого видения предмета.
10. Реципрокное торможение и его роль в управлении движениями.
реципрокноеторможение – осуществляется для координации активности мышц, противоположных по функции.
Различают девять Основные свойства нервных центров:
Пространственная и временная суммация основана на свойстве каждого нейрона в центре к суммации как возбуждения, так и торможения. Поскольку каждый нервный центр имеет много параллельно расположенных афферентных или входных волокон от рецептивного поля рефлекса, слабые раздражения нескольких участков рецептивного поля, в отдельности не способные реализовать рефлекс, вызывают в нейронах центра несколько ВПСП, которые суммируются, приводя к формированию на мембране нервной клетки потенциалов действия, распространяющихся по эфферентным проводникам, вызывая рефлекторную реакцию. Это явление называют пространственной суммацией. При увеличении частоты афферентных сигналов в единицу времени амплитуда ВПСП нарастает до критического уровня из-за повышения эффективности синаптического проведения, что также вызывает возбуждение нейронов и рефлекторный ответ на слабые частые раздражения. Это явление называют временной суммацией;
Центральная задержка рефлекса, характеризуется временем распространения информации в структурах нервного центра, главным образом в синапсах, где скорость проведения сигнала существенно меньше, чем в нервных проводниках. Поэтому, центральная задержка рефлекса зависит от количества синапсов между нейронами центра и представляет собой сумму синаптических задержек;
Посттетаническая потенциация — увеличение амплитуды ВПСП после серии частых (тетанизирующих) ритмических возбуждений, что связано с временной суммацией частых ВПСП и активацией синаптического проведения из-за увеличения числа квантов медиатора. Длительность состояния потенциации синапсов может достигать нескольких часов, что играет роль в процессах обучения и памяти:
Последействие и пролонгирование возбуждения — связаны с длительными следовыми потенциалами в нейронах, улучшением синаптического проведения, наличием кольцевых нейронных цепей и реверберацией возбуждения. Все эти процессы также играют роль в процессах обучения и памяти;
Трансформация ритма возбуждений, т.е. увеличение или уменьшение частоты нервных импульсов и эфферентных проводниках (на выходе) по сравнению с частотой афферентной импульсации (на входе центра), что связано с механизмом синаптической передачи (трансформация ритма как свойство синапса) и интегративной деятельностью нейронов;
Тонус нервного центра — состояние некоторого уровня активности нейронов, обеспечивающей их готовность к рефлекторной деятельности и проявляющейся в постоянной эфферентной импульсации низкой частоты к органам-эффекторам. Тонус нервных центров обусловлен небольшим уровнем афферентных сигналов от различных рецептивных полей (т.е. имеет рефлекторную природу), а также действием на нейроны метаболитов и других гуморальных раздражителей из клеточной микросреды. Проявлением тонуса нервных центров является спонтанная электрическая активность нейронов и некоторая фоновая активность эффекторов, например, тонус скелетной мускулатуры, гладких мышц сосудов и т.п.;
Утомление нервных центров — снижение эффективности их деятельности в виде повышения порогов возбуждения, связанное с утомлением синапсов и метаболическими сдвигами типа энергетического истощения в нервных клетках. Утомление формируеся при чрезмерной продолжительности действия раздражителей или их интенсивности, напряженном умственном труде или физической работе.
Нервный центр — совокупность нейронов, принимающих участие в осуществлении конкретного рефлекса (мигание, глотание, кашель и др.). Нейронной основой нервного центра служат локальные и иерархические нервные сети. Нервные центры обладают рядом характерных функциональных свойств, обусловленных объединением нейронов в нейронные сети и наличием межнейрональных синапсов. Свойства нейронов, связи между ними и синаптические структуры определяют:
- 1) суммацию возбуждений в нервных центрах:
- • временную (последовательную), когда импульсы возбуждения приходят к нейрону по одному и тему же пути через один синапс и обеспечивают суммацию В ПСП на постсинаптической мембране до уровня, достаточного для генерации нейроном потенциала действия;
• пространственную (одновременную), которая наблюдается при поступлении импульсов возбуждения к нейрону одновременно через разные синапсы (рис. 7.1.19);

Рис. 7.1.19. Суммация возбуждений в ЦНС: о — временная; 6 — пространственная
- 2) трансформацию ритма возбуждений — изменение числа импульсов возбуждения, выходящих из нервного центра, по сравнению с числом импульсов, приходящих к нему. Различают два вида трансформации:
- • понижающую, основанную на явлении суммации возбуждений (пространственной и временной);
- • повышающую, которую опосредуют механизмы умножения (мультипликации), способные резко увеличить число импульсов возбуждения (рис. 7.1.20);

Рис. 7.1.20. Умножение (мультипликация) возбуждения в ЦНС

Рис. 7.1.21. Пролонгирование возбуждения в ЦНС
- 4) тонус, который выражается в том, что даже при отсутствии специальных раздражений нервный центр постоянно посылает импульсы к рабочим органам;
- 5) пластичность — способность нервного центра изменять собственное функциональное назначение и расширять свои функциональные возможности;
- 6) одностороннее распространение возбуждения в нервных центрах — от рецептора к эффектору, обусловливаемое свойством химических синапсов односторонне проводить возбуждение от пре- синаптической к постсинаптической мембране;
- 7) задержку проведения возбуждения, обусловленную замедлением его проведения через синапсы.
- 8) высокую чувствительность к недостатку кислорода и действию различных химических веществ, особенно ядов. На одном нейроне могут располагаться синапсы, обладающие избирательной чувствительностью к различным химическим веществам;
- 9) быструю утомляемость (в отличие от нервных волокон, которые считаются практически неутомляемыми) и низкую лабильность;
- 10) низкую аккомодационную способность, т.е. способность реагировать на медленно нарастающие по силе раздражители.
Нервные центры имеют ряд общих свойств, которые во многом определяются структурой и функциями синаптических образований.
1. Одностороннее проведение возбуждения в нервном центре.
Распространение возбуждения обусловлено особенностями соединения нейронов в нервном центре. Поскольку в физиологических условиях возбуждение по нейрону распространяется от тела нейрона по его аксону, а синапсы проводят возбуждение только в одном направлении – от пресинаптической мембраны к постсинаптической, то возбуждение в нервном центре распространяется от входа через центральную часть к его выходным нейронам. Иногда часть нейронов может образовать замкнутую цепь, то есть нейронную ловушку, получившую название реверберационного круга. Возбуждение, распространившееся по реверберационному кругу, может циркулировать довольно долго, до тех пор, пока не изменится функциональное состояние какого-либо нейрона этой цепи.
2. Центральная задержка.
Центральной задержкой называется время от начала действия раздражителя на входе до появления возбуждения на выходе нервного центра. По отношению к рефлекторной дуге это время получило название латентного периода рефлекса. Центральная задержка определяется числом нейронов в центральной части нервного центра и временем распространения возбуждения через синапсы. В химических синапсах время выделения медиатора составляет около 0,3 мс, его действие на постсинаптическую мембрану, возникновение возбуждающего постсинаптического потенциала (ВПСП), его нарастание до критического уровня деполяризации (КУД) составляет 1 – 3 мс, таким образом время передачи возбуждения в одном синапсе составляет 2 – 3 мс.
Время центральной задержки будет состоять из времени проведения возбуждения по всем синапсам (отсюда можно предположить число нейронов в нервном центре) и времени распространения возбуждения по аксонам нейронов, которое очень мало, так как скорость распространения возбуждения составляет 10 – 70 м / с. Поскольку в состав нервного центра могут быть включены различные цепи с различным числом нейронов, обладающих разной возбудимостью, то время центральной задержки может зависеть также и от силы поступающего на вход раздражителя. Установлено, что чем сильнее раздражитель, тем это время короче, то есть распространение возбуждения осуществляется по более коротким цепям нейронов, обладающим, как правило, более высокой возбудимостью. Самой короткой такой цепью является моносинаптическая дуга коленного рефлекса, центральная задержка которого составляет около 3 мс.
3. Суммация возбуждений.
Это свойство заключается в том, что нервный центр может приходить в состояние возбуждения при действии на вход центра подпороговых раздражений, следующих друг за другом через определенные промежутки времени или одновременно с нескольких входных нейронов. Обязательным условием является то, что каждое приходящее возбуждение в отдельности является подпороговым для нервного центра. Это свойство нервных центров было впервые описано И.М.Сеченовым в 1863 году, а затем подробно изучено Ч.Шеррингтоном, И.П.Павловым, Дж.Экклсом.
Механизм, лежащий в основе процесса суммации, связан с интегративными функциями нервного центра и аналогичен механизму суммации отдельного нейрона. Суммация возбуждений зависит от свойств сомы, места и свойств синапса в нервном центре, от величины КУД, числа потенциалов действия и может быть пространственной и временной. Суть их заключается в следующем. Каждый нервный центр имеет множество афферентных, или входных волокон от различных участков рецептивного поля рефлекса. Слабые раздражители одного или нескольких участков рецептивного поля в отдельности не способны реализовать рефлекс, но, действуя на нервный центр одновременно, они генерируют ВПСП, которые суммируются, приводя к формированию на мембране нервных клеток потенциалов действия, распространяющихся по эфферентным проводникам, вызывая рефлекторную реакцию. Это явление называется пространственной суммацией. При увеличении частоты подпороговых афферентных сигналов, приходящих к одному и тому же входному нейрону, суммация ВПСП на постсинаптической мембране приводит к достижению КУД, что также вызывает возбуждение нервного центра и возникновение рефлекторной реакции на слабые, но частые стимулы. Это явление временной суммации.
4. Посттетаническая потенциация.
Величина ответной реакции нервных центров зависит не только от раздражения, действующего в настоящий момент, но и от предшествующих раздражений. Об этом убедительно свидетельствует свойство потенциации. Проявляется это свойство после действия частых ритмических предшествующих раздражений. Нервный центр приходит в такое состояние, что ранее подпороговый для него раздражитель становится надпороговым и вызывает возбуждение, а сила ответа нервного центра на некоторое время увеличивается.
Считают, что тетанизирующее ритмическое воздействие на нервный центр приводит к увеличению проницаемости пресинаптических мембран нейронов, увеличению вследствие этого выброса медиатора в синаптическую щель, что в свою очередь повышает возбудимость постсинаптической мембраны нейронов и всего нервного центра. Другим механизмом может служить особенность развития следовых процессов потенциалов действия у некоторых нейронов ЦНС, когда положительный и отрицательный следовые потенциалы длятся десятки и сотни миллисекунд.
Ответная реакция нервного центра не заканчивается, как правило, с окончанием возбуждения, поступающего на вход. Напротив, длительность ответа может занимать значительное время. Это свойство получило название последействия. Оно может быть тем продолжительнее, чем сильнее и дольше действовало возбуждение на входе. Принято считать, что механизмы этого явления могут быть различными. Если в состав нервного центра входят нейроны с длительной следовой деполяризацией, то нейрон при одиночном возбуждении может генерировать в течение нескольких десятков миллисекунд ритмический разряд импульсов, урежающийся по мере ослабления деполяризации.
Вторым механизмом может быть так называемая мультипликация ответа при распространении возбуждения в нервном центре по нейронным цепям, включающим различное число нейронов. Тогда на вход нервного центра из-за центральной задержки возбуждение от каждой такой нейронной цепи придет неодновременно, что и увеличит время ответной реакции, то есть последействия. Длительное же последействие возникает вследствие циркуляции нервных импульсов по замкнутым нейронным цепям. При таком включении нейронов возбуждение одного из них передается на другой (или другие), а по коллатералям их аксонов вновь возвращается к первой клетке и так далее. Благодаря существованию подобных кольцевых связей возбуждение может длительно циркулировать в нервном центре до тех пор, пока не наступит утомление одного из синапсов или же активность нейронов не будет приостановлена приходом тормозного импульса.
6. Трансформация ритма.
При подаче ритмического раздражения на вход нервного центра может возникнуть ситуация, когда на его выходе импульсация будет иметь новый ритм, который может быть больше или меньше исходного. Такое свойство нервного центра изменять ритм возбуждения получило название трансформации ритма. Механизмы, лежащие в основе трансформации ритма могут быть различными.
Во-первых, если в нервный центр включены нейроны, которые на один пришедший импульс отвечают серией (пачкой) импульсов, то на выходе такого нервного центра ритм возбуждения будет выше, чем на входе. Аналогичная ситуация возникает тогда, когда в нервном центре возбуждаются несколько нейронных цепей, включающих различное число нейронов.
7. Тонус нервного центра.
Электрофизиологические исследования показали, что не только при возбуждении нервного центра на входе можно получить поток импульсов на выходе, но и в состоянии относительного покоя из нервных центров на периферию поступают ритмические разряды импульсов. Такие центры обладают так называемой пейсмекерной активностью, а их постоянное возбуждение носит название тонуса нервных центров. Понятие тонуса нервных центров существовало уже в конце 19-го века. В поддержании тонуса нервных центров могут участвовать различные механизмы. С одной стороны, это постоянная афферентная импульсация от периферических рецепторов, поступающая в ЦНС, с другой - гуморальные воздействия, например, углекислый газ, гормоны, нейропептиды. Например, в исследованиях немецкого физиолога К.Гейманса было доказано существование постоянных тонических влияний на сердце со стороны центров, находящихся в продолговатом мозге. Происхождение этого тонуса во многом определяется пейсмекерной активностью нейронов ретикулярной формации, гуморальными факторами (изменение концентраций ионов калия и кальция, рН, содержанием углекислого газа в ликворе), и лишь в незначительной степени зависит от афферентных влияний, исходящих от рецепторов.
8. Пластичность нервного центра.
9. Утомляемость нервного центра.
В отличие от аксонов нейронов нервные центры легко утомляемы. Утомление нервного центра выражается в постепенном снижении импульсации на выходе при длительном поступлении нервной импульсации на вход. Считается, что это явление связано прежде всего с нарушением передачи возбуждения в межнейронных синапсах. Такое нарушение может быть обусловлено истощением синтезированного медиатора в нервных окончаниях, уменьшением чувствительности к медиатору постсинаптической мембраны, что может быть связано с истощением ферментов, инактивирующих медиатор, а также с уменьшением энергетических ресурсов нейронов.
10. Низкая лабильность нервного центра.
Как и все возбудимые структуры, нервные центры обладают лабильностью. Однако, в отличие от аксонов нейронов, где лабильность достигает 500 Гц, нервные центры обладают низкой лабильностью, порядка 15 – 50 Гц и менее. Это связано со сложной структурой нервного центра, большим числом нейронов в его составе, а также межнейронных синапсов, особенностями распространения возбуждения внутри нервного центра.
11. Высокая чувствительность нервных центров к недостатку кислорода, специфическим ядам и фармакологическим средствам.
Нервные клетки отличаются интенсивным обменом веществ и большим потреблением кислорода. Мозг человека поглощает приблизительно 40 – 50 мл кислорода в минуту. Потребляя большое количество кислорода, нервные клетки высокочувствительны к его недостатку. Особенно чувствительны у человека нейроны коры больших полушарий головного мозга, уже через 5 – 6 мин без кислорода они подвергаются необратимым изменениям и гибнут. Нервные центры ствола мозга менее чувствительны к недостатку кислорода: их функции восстанавливаются даже после 15 – 20 мин полного прекращения поступления кислорода, а центры спинного мозга могут восстанавливать свои функции даже через 20 – 30 мин после полного прекращения поступления к ним кислорода.
Заключение
Нервные клетки и синапсы обладают избирательной чувствительностью к некоторым ядам и фармакологическим средствам. Например, апоморфин избирательно возбуждает нервный центр рвоты, а лобелин возбуждает дыхательный центр. Другие вещества, получившие название ганглиоблокаторов, угнетают передачу в нервных центрах автономной нервной системы. Некоторые вещества влияют на огромные популяции нейронов, например, кардиазол избирательно действует на двигательные нейроны коры больших полушарий, а мескалин оказывает влияние на зрительные центры головного мозга. Эти явления, очевидно, связаны с тем, что химические процессы, происходящие в данных нейронах и синапсах, сходны с действием химических веществ на указанные структуры. К настоящему времени открыты и частично получены синтетически многие химические вещества, которые могут блокировать передачу возбуждения в синапсах ЦНС или возбуждать их искусственным путем.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Читайте также:
