
Что такое временная суммация нервных центров
Нервный центр – это совокупность нейронов, обеспечивающих регуляцию какого-либо конкретного физиологического процесса или функции.
Нервный центр в узком смысле – это совокупность нейронов, без которых данная конкретная функция не может регулироваться. Например, без нейронов дыхательного центра продолговатого мозга дыхание прекращается. Нервный центр в широком смысле — это совокупность нейронов, которые участвуют в регуляции конкретной физиологической функции, но не являются строго обязательными для ее осуществления! Например, в регуляции дыхания кроме нейронов продолговатого мозга участвуют нейроны пневмотаксического центра варолиевого моста, отдельные ядра гипоталамуса, кора больших полушарий и другие образования головного мозга.
Все нейроны нервного центра разделяют на 2 неравные по количеству и качеству группы.
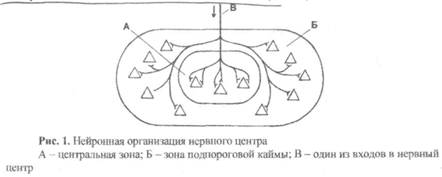
Первая группа – нейроны центральной зоны. Это наиболее возбудимые нейроны, которые возбуждаются в ответ на поступление порогового (для нервного центра) сигнала. Таких нейронов около 15-20%, и они не обязательно располагаются в середине нервного центра, как это изображено на рис.1. Особенностью их является то, что они имеют на своем теле больше синаптических терминалей от сенсорных и вставочных нейронов.
Вторая группа – нейроны подпороговой каймы. Это менее возбудимые нейроны, которые не возбуждаются в ответ на поступление пороговых им-пульсов, но при действии более сильных раздражителей они возбуждаются и включаются в работу нервного центра, обеспечивая ее усиление. Таких нейронов большинство (80-85%), и они не обязательно располагаются на периферии нервного центра, но все имеют значительно меньше синаптических терминалей от сенсорных и вставочных нейронов по сравнению с нейронами центральной зоны.
На рис. 1 нейроны центральной зоны условно поставлены в центр внутреннего круга (А), а нейроны подпороговой каймы – в пространство между внутренним и наружным кругами (Б). Таким образом, если к нервному центру по афферентному входу (В) придет пороговый импульс, то возбудятся три нейрона центральной зоны, а на десяти нейронах подпороговой каймы потенциалы действия не возникнут, но появится местная деполяризация – возбуждающий постсинаптический потенциал (ВПСП).
От структуры нервного центра зависят его свойства, а они, в свою очередь, влияют на процесс проведения возбуждения через нервный центр, на его скорость и степень выраженности. От свойств нервных центров во многом зависит процесс распространения возбуждения по ЦНС, что имеет важное значение в интегративной деятельности организма.
Свойства нервных центров обусловлены описанной выше нейронной организацией нервного центра, а также химическим способом передачи возбуждения в синапсах. При электрическом способе передачи возбуждения нервные центры не имели бы подобных свойств.
Свойства нервных центров: 1 одностороннее проведение возбуждения; 2 задержка проведения возбуждения; 3 суммация; 4 облегчение; 5 окклюзия; 6 мультипликация; 7 трансформация; 8 последействие; 9 посттетаническая потенциация; 10 утомление; 11 тонус; 12 высокая чувствительность к изменению состояния внутренней среды организма; 13 пластичность.
3) Суммация – это возникновение возбуждения в нервном центре при поступлении к нему нескольких допороговых импульсов, каждый из которых в отдельности не может возбуждения (рис. 2). Фактически этот процесс происходит на нейронах подпороговой каймы. Различают два вида суммации: пространственную и временною.
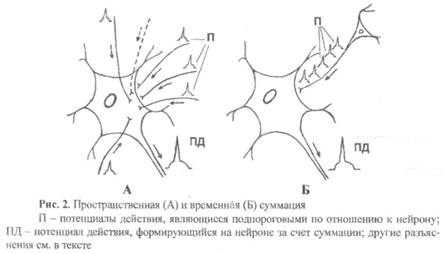
Пространственная суммация возникает в том случае, когда к нервному центру (к его нейронам) приходят одновременно, несколько допороговых импульсов. На рисунке 2А видно, что к нейрону подпороговой каймы, имеющему пороговый потенциал 30 мВ одновременно по пяти различным афферентным входам (их аксоны обозначены сплошной линией) приходят пять импульсов, каждый из которых деполяризует мембрану нейрона на 5 мВ (то есть возникают пять отдельных ВПСП). В этом случае возбуждение нейрона не наступает, так как суммарная деполяризация мембраны нейрона составляет лишь 25 мВ (суммированный ВПСП мал для достижения КУД). Но если к нейрону придет еще один подобный импульс по шестому входу (его аксон обозначен пунктирной линией), то суммированный ВПСП будет достаточен по величине и мембрана нейрона в зоне аксонного холмика деполяризуется до критического уровня, в результате чего нейрон из состояния покоя перейдет в состояние возбуждения. На постсинаптической мембране происходит суммация ВПСП в пространстве.
Временная (последовательная) суммация возникает в том случае, когда к нейронам нервного центра по одному афферентному входу приходит не один, а серия импульсов с очень небольшими по времени межимпульсными промежутками (рис. 2Б). Два механизма временной суммации:
1) интервалы между отдельными импульсами настолько малы, что за это время медиатор, выделившийся в синаптическую щель, не успевает полностью разрушиться и вернуться в пресинаптический аппарат. В этом случае возникает постепенное накопление медиатора до критического объема, необходимого для возникновения достаточного по амплитуде ВПСП, а значит, и для возникновения ПД;
2) интервалы между отдельными импульсами настолько малы, что возникший за это время на постсинаптической мембране ВПСП не успевает исчезнуть и усиливается за счет новой порции медиатора – суммируется. На постсинаптической мембране происходит суммация ВПСП во времени.
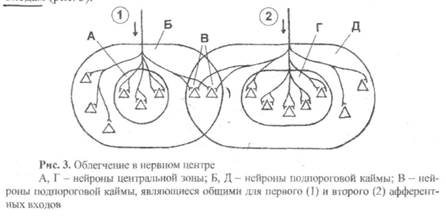
4) Облегчение – это увеличение количества возбужденных нейронов в нервном центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному, а по двум или более афферентным входам. На рис. 3 рассмотрен случай, когда при отдельном раздражении первого афферентного входа возбуждается только три нейрона центральной зоны (А), а на пяти нейронах подпороговой каймы (Б) возникают ВПСП. Если раздражать отдельно только второй афферентный вход, то возбуждены будут пять нейронов (Г), а четыре нейрона подпороговой каймы (Д) не возбудятся. Раздражая и первый, и второй афферентные входы одновременно (!), мы ожидаем вовлечения в процесс возбуждения восьми нейронов. И они, естественно, будут возбуждаться, но кроме них (сверх ожидания!) могут возбуждаться еще некоторые нейроны подпороговой каймы. Это произойдет потому, что один или несколько нейронов подпороговой каймы являются общими как для первого, так и для второго афферентных входов (в нашем случае это два нейрона, обозначенные буквой В), и при одновременном поступлении возбуждения к этим нейронам дни возбудятся за счет возникновения пространственной суммации.
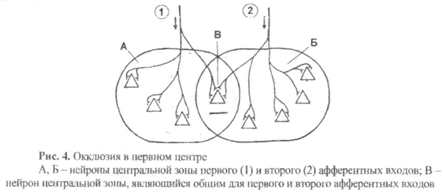
5) Окклюзия – это уменьшение количества возбужденных нейронов в нервной центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному. а по двум или более афферентным входам (рис. 4).
6) Мультипликационное возбуждение (мультипликация) заключается в том, что по разветвлениям аксона вставочного нейрона возбуждение поступает одновременно не на один, а на несколько моторных нейронов (рис. 6). В связи с этим эффект на рабочем органе усиливается в несколько раз, или в работу вовлекаются не одна, а несколько рабочих структур, Это свойство особенно ярко проявляется в ганглиях автономной (вегетативной) нервной системы.
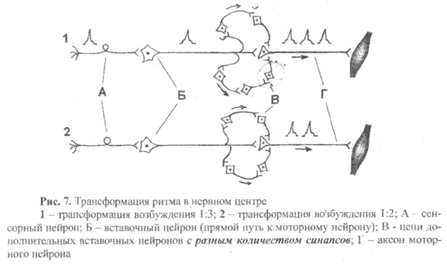
7) Трансформация ритма возбуждения – это изменение частоты импульсов на выходе из нервного центра по сравнению с частотой импульсов на входе в нервный центр.
а) наличием дублирующих цепей вставочных нейронов, связывающих сенсорные и моторные нейроны;
б) разным количеством синапсов в каждой из этих цепей.


Например, на рис.7 представлены два варианта трансформации, которые, на первый взгляд, не отличаются друг от друга, так как в том и в другом случае показаны две дополнительные цепи вставочных нейронов (кроме прямого пути), с помощью которых возбуждение может передаваться по цепи нейронов А—Б—В. Рассмотрим эти схемы.
Вариант 1. Верхняя цепь состоит из двух дополнительных вставочных нейронов, а значит, по сравнению с прямым путем передачи возбуждения с нейрона Б на нейрон В, имеет два дополнительных синапса. Поэтому возбуждение, проходя по верхней цепи, задержится на 2 мс (время синаптической задержки в одном синапсе составляет
1 мс) и придет на нейрон В после того, как пройдет возбуждение по прямому пути. В нижней цепи три дополнительных вставочных нейрона (то есть три дополнительных синапса), значит, возбуждение будет доходить до нейрона В еще дольше, чем по верхней цепи (задержка составит 3 мс). Следовательно, по нижней цепи возбуждение на нейрон В придет после того, как пройдет возбуждение по верхней цепи. В результате на один импульс, пришедший по сенсорному нейрону А, на моторном нейроне В возникнет три потенциала действия (трансформация 1:3).
Вариант 2. В этом случае и верхняя и нижняя цепи вставочных нейронов состоят из двух дополнительных нейронов. Возбуждение по обеим цепям придет к нейрону В одновременно в виде одного потенциала действие, который появится на нейроне В только после прохождения возбуждения к нему от нейрона Б по прямому пути. В этом варианте мы тоже получим трансформацию ритма, но уже в соотношении 1:2.
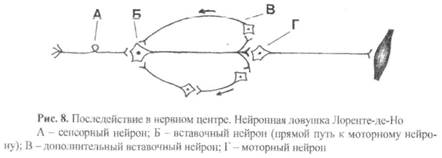
8) Последействие – это продолжение возбуждения моторного нейрона в течение некоторого времени после прекращения действия раздражителя.
Другими причинами последействия могут быть:
а) возникновение высокоамплитудного ВПСП, в результате которого возникает не один, а несколько потенциалов действия то есть ответ длится большее время;
б) длительная следовая деполяризация постсинаптической мембраны, в результате чего возникают несколько потенциалов действия, вместо одного.
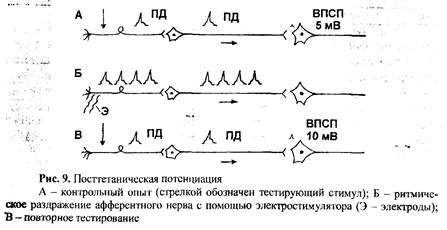
9) Посттетаническая потенциация (синаптическое облегчение) – это улучшение проведения в синапсах после короткого раздражения афферентных путей.
Если в качестве контроля вызвать одиночное раздражение афферентного нерва тестирующим раздражителем (рис. 9А), то на моторном нейроне мы получим ВПСП вполне определенной амплитуды (в нашем случае 5 мВ). Если после этого тот же афферентный нерв раздражать некоторое время серией частых импульсов (рис. 9Б), а потом вновь подействовать тестирующим раздражителем (рис. 9В), то величина ВПСП будет больше (в нашем случае 10 мВ). Причем она будет тем больше, чем более частыми импульсами мы раздражали афферентный нерв.
Длительность синаптического облегчения зависит от свойств синапса и характера раздражения: после одиночных стимулов оно выражено слабо, после раздражающей серии потенциация (облегчение) может продолжаться от нескольких минут до нескольких часов. Объясняется он тем, что при частом раздражении афферентного волокна в его пресинаптической терминали (окончании) накапливаются ионы кальция, а значит, улучшается выделение медиатора. Кроме того, показано, что частое раздражение нерва приводит к усилению синтеза медиатора, мобилизации пузырьков медиатора, к усилению синтеза белков-рецепторов на постсинаптической мембране и увеличению их чувтствительности. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах.
10) Утомление нервного центра (посттетаническая депрессия, синаптическая депрессия) – это уменьшение или прекращение импульсной активности нервного центра в результате длительной стимуляции его афферентными импульсами (или произвольного вовлечения его в процесс возбуждения посредством импульсов, идущих из коры больших полушарий). Причинами утомления нервного центра могут быть:
• истощение запасов медиатора в афферентном или вставочном нейроне;
• снижение возбудимости постсинаптической мембраны (то есть мембраны моторного или вставочного нейрона) из-за накопления, например, продуктов метаболизма.
Утомляемость нервных центров продемонстрировал Н.Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения п. tibialis и п. peroneus. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги. Синаптическая депрессия при длительной активации центра выражается в снижении постсинаптических потенциалов.
11) Тонус нервного центра – это длительное, умеренное возбуждение нервного центра без видимо утомления Причинами тонуса могут быть:
• потоки афферентных импульсов, постоянно поступающие с неадаптирующихся рецепторов;
• гуморальные факторы, постоянно присутствующие в плазме крови;
• спонтанная биоэлектрическая активность нейронов (автоматия);
• циркуляция (реверберация) импульсов в ЦНС.
12) Нервный центр состоит из нейронов, а они очень чувствительны к изменению состава внутренней среды организма, что и отражается на свойствах нервных центров. Наиболее важными факторами, влияющими на работу нервных центров, являются: гипоксия; недостаток питательных веществ (например, глюкозы); изменение температуры; воздействие продуктов метаболизма; воздействие различных токсических и фармакологических препаратов.
Разные нервные центры имеют неодинаковую чувствительность к воздействию названных факторов. Так, нейроны коры больших полушарий наиболее чувствительны к гипоксии, недостатку глюкозы, продуктам метаболизма; клетки гипоталамуса – к изменению температуры, содержанию глюкозы, аминокислот, жирных кислот и др.; различные участки ретикулярной формации выключаются разными фармакологическими препаратами, различные нервные центры избирательно активируются или тормозятся разными медиаторами.
13) Пластичность нервного центра означает его способность изменять при определенных обстоятельствах свои функциональные свойства. В основе этого явления лежит поливалентность нейронов нервных центров. Особенно ярко проявляется это свойство при всевозможных повреждениях ЦНС, когда организм компенсирует утраченные функции за счет сохранившихся нервных центров. Особенно хорошо свойство пластичности выражено в коре больших полушарий. Например, центральные параличи, связанные с патологией двигательных центров коры, иногда полностью компенсируются, и ранее утраченные двигательные функции восстанавливаются.
Дата добавления: 2015-01-21 ; просмотров: 21329 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Нервным центром называется совокупность нейронов в различных отделах ЦНС, обеспечивающих регуляцию какой-либо функции организма. Например, бульбарный дыхательный центр.
Для проведения возбуждения через нервные центры характерны следующие особенности:
1. Одностороннее проведение. Оно идет от афферентного, через вставочный, к эфферентному нейрону. Это обусловлено наличием межнейронных синапсов.
2. Центральная задержка проведение возбуждения, т.е. по ЦНС возбуждение идет значительно медленнее, чем по нервному волокну. Это объясняется синаптической задержкой. Так как больше всего синапсов в центральном звене рефлекторной дуги, там скорость проведения наименьшая. Исходя из этого время рефлекса – это время от начала воздействия раздражителя до появления ответной реакции. Чем длительнее центральная задержка, тем больше время рефлекса. Вместе с тем оно зависит от силы раздражителя. Чем оно больше, тем время рефлекса короче и наоборот. Это объясняется явлением суммации возбуждений в синапсах. Кроме того, оно определяется и функциональным состоянием ЦНС. Например, при утомлении нервного центра длительность рефлекторной реакции увеличивается.
3. Пространственная и временная суммация. Временная суммация возникает как в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда возбуждающего постсинаптического потенциала. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторных нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются, и в мембране нейрона генерируется распространяющийся потенциал действия.
4. Трансформация ритма возбуждения – изменение частоты нервных импульсов при прохождении через нервный центр. Частота может снижаться или повышаться. Например, повышающая трансформация – увеличение частоты обусловлено дисперсией и мультипликацией возбуждения в нейронах. Первое явление возникает в результате разделения нервных импульсов на несколько нейронов, аксоны которых образуют затем синапсы на одном нейроне. Второе – генерацией нескольких нервных импульсов при развитии возбуждающего постсинаптического потенциала на мембране одного нейрона. Понижающая трансформация объясняется суммацией нескольких возбуждающих постсинаптических потенциалов и возникновением одного потенциала действия в нейроне.
5. Посттетаническая потенциация – это усиление рефлекторной реакции в результате двигательного возбуждения нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы, выделяется большое количество нейромедиаторов в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
6. Последействие – это запаздывание окончания рефлекторного ответа после прекращения действия раздражителя. Связано с циркуляцией нервных импульсов по замкнутым цепям нейронов.
7. Тонус нервных центров – состояние постоянной повышенной активности. Он обусловлен постоянным поступлением к нервному центру нервных импульсов от периферических рецепторов, возбуждающим влиянием на нейроны продуктов метаболизма и других гуморальных факторов. Например, проявлением тонуса соответствующих центров является тонус определенной группы мышц.
8. Автоматия (спонтанная активность) нервных центров. Периодическая или постоянная генерация нейронами нервных импульсов, которые возникают в них самопроизвольно, т.е. в отсутствии сигналов от других нейронов или рецепторов. Обусловлена колебаниями процессов метаболизма в нейронах и действием на них гуморальных факторов.
9. Пластичность нервных центров. Это их способность изменять функциональные свойства. При этом центр приобретает возможность выполнять новые функции или восстанавливать старые после повреждения. В основе пластичности нервного центра лежит пластичность синапсов и мембран нейронов, которые могут изменять свою молекулярную структуру.
10. Низкая физиологическая лабильность и быстрая утомляемость. Нервные центры могут проводить импульсы лишь ограниченной частоты. Их утомление объясняется утомлением синапсов и ухудшением метаболизма нейронов, истощение состава медиаторов, длительность их синтеза.
Суммация возбуждения в ЦНС. Это явление было открыто русским физиологом И.М. Сеченовым в 1868г. в опыте на лягушке. Раздражение конечности лягушки слабыми редкими импульсами не вызывало реакцию, а более частые раздражения такими же слабыми импульсами сопровождались ответной реакцией – лягушка совершала прыжок. Различают временную (последовательную) и пространственную суммацию.
Временная суммация обусловлена тем, что ВПСП от предыдущего импульса еще продолжается, когда приходит следующий импульс. Поэтому данный вид суммации называют также последовательной суммацией, которая играет важную физиологическую роль, потому что многие нейронные процессы имеют ритмический характер и, таким образом, могут суммироваться, давая начало надпороговому возбуждению в нейронных объединениях нервных центров.
Пространственная суммация. Раздельная стимуляция каждого из двух аксонов вызывает подпороговый ВПСП, тогда как при одновременной стимуляции обоих аксонов возникает ПД – процесс, который не может быть обеспечен одиночным ВПСП. Пространственная суммация связана с такой особенностью распространения возбуждения, как конвергенция. Последствие – продолжение возбуждения нервного центра после прекращения поступления к нему импульсов по афферентным нервным путям. Последствие вызывает ряд причин:
а) длительное существование ВПСП (полисинаптического или высокоамплитудного), когда при одном ВПСП возникает несколько ПД;
б) многократные появления следовой деполяризации, что свойственно нейронам ЦНС;
в) циркуляция возбуждения по замкнутым нейронным цепям.
Последствие играет важнейшую роль в процессах обучения (кратковременная память).
Фоновая активность нервных центров (тонус)объясняется:
а) спонтанной активностью нейронов ЦНС;
б) гуморальными влияниями биологически активных веществ (метаболитов, гормонов, медиаторов и т.д.), циркулирующих в крови и влияющих на возбудимость нейронов;
в) афферентной импульсацией от различных рефлексогенных зон;
г) суммацией миниатюрных потенциалов, возникающих в результате спонтанного выделения квантов медиатора из аксонов, образующих синапсы на нейронах;
д) циркуляцией возбуждения в ЦНС.
Значение фоновой активности нервных центров заключается в обеспечении некоторого исходного уровня деятельного состояния центра и эффекторов.
Трансформация ритма возбуждения– это изменение числа импульсов, возникающих в нейронах центра на выходе относительно числа импульсов, поступающих на вход данного центра. Трансформация ритма возбуждения возможна как в сторону увеличения, так и в сторону уменьшения, поскольку для возбуждения нейрона нужен поток афферентных импульсов.
Большая чувствительность ЦНС к изменениям внутренней среды, например к изменению содержания глюкозы в крови, газового состава крови, температуры, к вводимым с лечебной целью различным фармакологическим препаратам. В первую очередь реагируют синапсы нейронов. Особенно чувствительны нейроны ЦНС к недостатку глюкозы и кислорода. При снижении содержания глюкозы в 2 раза ниже нормы могут возникнуть судороги. К тяжелым последствиям для ЦНС приводит недостаток кислорода в крови. Прекращение кровотока всего лишь на 10с вызывает очевидные нарушения функций мозга, человек теряет сознание, а прекращение кровотока на 8-12 мин вызывает необратимые нарушения деятельности мозга, т.е. погибают многие нейроны, в первую очередь корковые, что ведет к тяжелым последствиям.
Пластичность нервных центровспособность нервных элементов к перестройке функциональных свойств:
Синаптическое облегчение (потенциация) – улучшение проведения в синапсах после короткого раздражения афферентных путей. Длительность синаптической потенциации зависит от свойств синапса и характера раздражения: после одиночных стимулов она выражена слабо, после раздражающей серии облегчение в ЦНС может продолжаться от нескольких минут до нескольких часов. Главной причиной возникновения синаптического облегчения является накопление Са 2+ в пресинаптических окончаниях, поскольку Са 2+ , входящий в нервное окончание во время ПД, накапливается там, т.к. ионная помпа не успевает выводить его. Кроме того, при частом использовании синапсов ускоряется синтез рецепторов и медиатора, а также мобилизация пузырьков, однако при редком использовании синапсов синтез медиаторов уменьшается.
Значение синаптического облегчения заключается в том, что оно создает предпосылки для улучшения процессов переработки информации на нейронах нервных центров, что крайне важно, например, выработки двигательных навыков и условных рефлексов. Повторное возникновение явление облегчения в нервном центре может вызвать переход центра из обычного состояния в доминантное.
Образование временных связей, обеспечивающих формирование условных рефлексов, чему способствует синаптическое облегчение и доминантное состояния двух центров. Например, сочетание звука колокольчика с подачей мясного корма вызывает у экспериментальной собаки слюноотделение. После повторения этого воздействия звучание только колокольчика вызывает такое же слюноотделение, как и мясо. В основе механизма выработки условного рефлекса лежит явление доминанты.
Доминанта – стойкий господствующий очаг возбуждения в ЦНС, подчиняющий себе в данный момент функции других нервных центров. Явление доминанты открыл А.А. Ухтомский в 1923г. Если раздражать корковую двигательную зону на фоне избыточного повышения возбудимости другого нервного центра, то сгибания конечности может не произойти. Вместо сгибания конечности раздражения двигательной зоны может вызвать реакцию тех эффекторов, деятельность которых контролируется господствующим, т.е. доминирующим, в данный момент в ЦНС нервным центром. Доминантный очаг возбуждения обладает рядом особых свойств, главными из которых является инерционность, стойкость, повышенная возбудимость, способность притягивать к себе иррадиирующие по ЦНС возбуждения и оказывать угнетающее влияние на центры-конкуренты и другие нервные центры.
Значение доминантного очага возбуждения в ЦНС заключается в том, что на его базе формируется конкретная приспособительная деятельность, в том числе и условно-рефлекторная, ориентированная на достижение полезных результатов, необходимых для устранения причин, поддерживающих тот или иной нервный центр в доминантном состоянии.
Синаптическая депрессия (утомление центра) развивается при длительной посылке импульсов к центру. Утомляемость нервных центров продемонстрировал Н.Е. Введенский в опыте над лягушкой. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение на раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги. Синаптическая депрессия (утомляемость синапсов) при длительной активации центра выражается в снижении числа постсинаптических потенциалов. Это обусловлено расходованием медиатора, накоплением метаболитов, в частности закислением среды при длительном проведении возбуждения по одним и тем же нейронным центрам.
Компенсация нарушенных функций после повреждения того или иного центра – также результат проявления пластичности ЦНС. Отмечалось, что парализованные конечности у больных постепенно начинает вовлекаться в двигательную активность, при этом нормализуется тонус ее мышц. Нарушенная двигательная функция частично, а иногда полностью восстанавливается за счет большей активности сохранившихся нейронов и вовлечении в эту функцию других (рассеянных) нейронов с подобными функциями в коре большого мозга. Этому способствуют регулярные пассивные и активные движения.
Возбуждений, конвергенция, дивергенция, окклюзия, реверберация.
Пространственная и временная суммация: основана на свойстве каждого нейрона в центре к суммации как возбуждения, так и торможения. В основе лежит конвергенция. Поскольку каждый нервный центр имеет много параллельно расположенных афферентных или входных волокон от рецептивного поля рефлекса, слабые раздражения нескольких участков рецептивного поля, в отдельности не способные реализовать рефлекс, вызывают в нейронах центра несколько ВПСП, которые суммируются, приводя к формированию на мембране нервной клетки потенциалов действия, распространяющихся по эфферентным проводникам, вызывая рефлекторную реакцию. Это явление называют пространственной суммацией. При увеличении частоты афферентных сигналов в единицу времени амплитуда ВПСП нарастает до критического уровня из-за повышения эффективности синаптического проведения, что также вызывает возбуждение нейронов и рефлекторный ответ на слабые частые раздражения - временная суммация;
Конвергенция - способность возбуждений сходиться к одним и тем же промежуточным и конечным нейронам получила название конвергенции возбуждений. Процесс конвергенции характерен не только для однотипных нервных клеток.
Дивергенция (расхождение) - расхождение импульсаций от одного нейрона сразу на многие нейроны. На основе дивергенции происходит иррадиация возбуждения и становится возможным быстрое вовлечение в ответную реакцию многих центров, расположенных на разных уровнях ЦНС.
Окклюзия (запирание) - это сложение пороговой импульсации. Окклюзия осуществляется (так же как и пространственная суммация) в конвергирующей системе соединения нейронов. Одновременной активации нескольких (минимум двух) рецепторов сильным или сверхсильным раздражителями к одному нейрону будут конвергировать несколько пороговых или сверхпороговых импульса. На этом нейроне будет происходить окклюзия, т.е. эти два раздражителя он ответит с той же максимальной силой, что и на каждый из них отдельности. Феномен окклюзии состоит в том, что количество возбужденных нейронов при одновременном раздражении афферентных входов обоих нервных центров оказывается меньше, чем арифметическая сумма возбужденных нейронов при отдельном раздражении каждого афферентного входа в отдельности.
Явление окклюзии приводит к снижению силы ответной реакции. Окклюзия имеет охранительное значение, предотвращая перенапряжение нейронов при действии сверхсильных раздражителей.
Ревербация - циркуляция возбуждения замкнутыми нейронами и их цепями в ЦНС. Возбуждение одного из нейронов, входящих в эту цепь, передается на другой (или другие), а коллатералям аксонов снова возвращается к нервной клетки и т.д.
Реверберация возбуждения наблюдается в так называемом рефлекторном последействии, когда рефлекторный акт заканчивается не сразу после прекращения, а через некоторый период.
6. Координация рефлекторной деятельности в ЦНС. Принципы координации:
Доминанта, общий конечный путь, реципрокное торможение, обратная афферентная
Связь.
Координация - согласованную деятельность ЦНС, благодаря взаимодействиию нейронов и нервных процессов (возбуждения и торможения) в ЦНС. Благодаря координации возможно точное выполнение в данный момент различных сложных рефлекторных актов, к которым относятся двигательные, секреторные, сосудистые и другие компоненты. Координация происходит во всех отделах ЦНС, в любом нервном центре.
Согласованная деятельность обеспечивается за счет ряда механизмов:
1. Принцип доминанты. Он был сформулирован А.А.Ухтомским как основной принцип работы нервных центров.
Доминантный очаг возбуждения характеризуется следующими свойствами:
· стойкостью возбуждения, т.е. может сохраняться длительное время;
· способностью к суммации возбуждений, притягивая на себя возбуждение с других центров;
· способностью тормозить субдоминантные очаги возбуждения других нервных центров.
2. Принцип общего конечного пути. Эффекторные нейроны ЦНС, например, мотонейроны спинного мозга, могут вовлекаться в осуществление различных реакций организма возбуждениями, приходящими к ним от большого числа афферентных и промежуточных нейронов, для которых они являются конечным путем (путем от ЦНС к эффектору).
3. Принцип обратной связи. За счет импульсов, поступающих по этому каналу, происходит оценка правильности исполнения поставленной задачи, а если она не выполнена, то вносятся коррекции для достижения результата. Велико значение механизмов обратной связи в поддержании гомеостаза. Так, например, поддержание постоянного уровня кровяного давления осуществляется за счет изменения импульсной активности барорецепторов сосудистых рефлексогенных зон, в результате чего измененяеся тонус вазомоторных симпатических нервов и таким образом нормализуют кровяное давление.
4. Принцип реципрокности. Он отражает характер отношений между центрами ответственными за осуществление противоположных функций (вдоха и глотания, выдоха и выдоха, сгибания и разгибания конечностей и т.д.). Например, активация проприорецепторов мышцы-сгибателя одновременно возбуждает центр мышц сгибателей и тормозит центр мышц разгибателей. Реципрокное торможение играет важную роль в координации двигательных актов.
Последнее изменение этой страницы: 2017-02-19; Нарушение авторского права страницы
Читайте также:
