
Чувствительные нервные окончания реагирующие нa изменение температуры
Классификация
1. По месту восприятия раздражения:
1.1. Экстерорецепторы, воспринимающие раздражения из внешней среды;
1.2. Интерорецепторы – восприятие раздражений внутренних органов;
1.3. Проприорецепторы – восприятие информации от опорно-двигательного аппарата.
2. В зависимости от природы раздражения, воспринимаемого рецептором:
2.1. Механорецепторы – механические раздражители, перемещение частей тела;
2.2. Хеморецепторы – химические раздражители.
2.3. Терморецепторы – изменения температуры.
2.4. Ноцирецепторы – чувство боли.
3. По способу восприятия:
3.1. Контактные рецепторы.
3.2. Дистантные рецепторы (рецепторные клетки сетчатки глаза, органа слуха, обоняния).
4. Морфологическая классификация основана на особенностях их структурной организации.
4.1. Свободные нервные окончания состоят только из конечных ветвлений дендрита чувствительного нейрона. Они встречаются в эпителии (кожа и серозные оболочки) и в соединительной ткани, обеспечивая восприятие температурных, механических и болевых сигналов.
4.2. Несвободные нервные окончания – содержат все компоненты нервного волокна (осевой цилиндр и клетки глии). Они могут быть инкапсулированными (имеющими особую соединительнотканную капсулу) и неинкапсулированные.
Неинкапсулировпнные нервные окончания представлены обязательными дисками Меркеля и встречаются в дерме кожи и собственной пластинки слизистых оболочек.
Инкапсулированные нервные окончания имеют общий принцип строения: их основу составляют ветвления дендрита, которые окружены леммоцитами и покрыты соединительнотканной капсулой. К этому виду окончаний относят: пластинчатые тельца (Фатер-Пачини), обязательные тельца (Мейснера), тельца Руффини, колбы Краузе, генитальные тельца Догеля, нервно-мышечные и нервно-сухожильные веретена.
Пластинчатые тельца (Фатер-Пачини) – в соединительной ткани внутренних органов и кожи. Имеют вид округлых образований (1-5 мм), воспринимают давление и вибрацию.
Структурными компонентами являются:
1) внутренняя колба (луковица), образованная видозимененными леммоцитами, в которую проникают нервные волокна;
2) наружная колба – слоистая соединительнотканная капсула из фибропластов и коллагеновых волокон, образующих концентрические пластины, между которыми имеется жидкость.
При деформации пластин давление передается на нервные окончания и вызывает деполяризацию мембраны.
Осязательные тельца (Мейснера) – в сосочковом слое дермы, имеют эллипсоидную форму и небольшие размеры (50-140 мкм). Внутренняя колба состоит из глиальных клеток, лежащих перпендикулярно длинной оси тельца, между которыми располагаются веточки дендритов. Снаружи находится очень тонкая, переходящая в периневрит слоистая капсула – наружная колба. Незначительная деформация капсулы передается глиоцитам и далее на дендриты.
Тельца Руффини лежат в соединительнотканной части кожи и капсулах суставов: они воспринимают давление и имеют вид веретеновидных структур длиной 1-2 мм. Внутреннюю колбу образуют глиальные клетки между которыми располагаются терминали дендритов с расширениями на концах. Капсула хорошо выражена.
Колбы Краузе – мелкие (40-150 мкм) округлые тельца, являющиеся механорецепторами и Холодовыми рецепторами. Они лежат в дерме кожи, слизистой оболочке полости рта, подгортанника, в конъюнктиве глаза. Внутренняя колба образована плоскими глиоцитами между которыми тонкие ветви дендрита образуют сплетения в виде клубочка. Наружная капсула очень тонкая.
Генитальные тельца Догеля – находятся в особо чувствительных областях кожи (наружные половые органы, молочные железы). Они по строению похожи на колбы Краузе, но в отличие от них в тельце входит несколько отростков от нейроцитов, что вызывает сильную иррадиацию возбуждения.
Нервно-мышечные веретена – рецепторы растяжения поперечнополосатых мышц - нервные окончания, обладающие как чувствительной, так и двигательной иннерваций. Наружная капсула веретена окружает несколько тонких интрафузальных мышечных волокон. Волокна, лежащие за пределами капсулы, называются экстрафузальными. Интрафузальные мышечные волокна двух видов:
- волокна с ядерной сумкой – в расширенной центральной части которых содержатся скопления ядер (ядерная сумка);
- волокна с ядерной цепочкой – более тонкие, ядра лежат по всей длине волокна в виде цепочки в центральной части (ядерная цепь).
Чувствительные нервные волокна образуют кольцеспиральные окончания (отростки нервных клеток закручены вокруг центральной части волокна по спирали) на волокнах обоих типов и гроздьевидные окончания у краев волокон с ядерной цепочкой.
Двигательные нервные волокна образуют мелкие нервномышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус, регулируя длину волокон. Всё свободное пространство между мышечными волокнами заполнено жидкостью и ограничено тонкой капсулой. Изменение тонуса мышцы ведет к изменению давления жидкости и передается на дендриты. Кольцеспиральные окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, гроздьевидные окончания – только на изменение длины. Число веретен в мышце зависит от её функции и тем выше, чем более точными движениями она обладает.
Нервно-сухожильные веретена – рецепторы растяжения, располагаются в месте соединения мышцы с сухожилием, веретеновидные структуры длиной 0,5-1 мм. Каждое веретено имеет капсулу из фиброцитов, которая охватывает группу сухожильных пучков, оплетенных окончаниями нервных волокон. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.
Рефлекторная дуга – это цепь нейронов, связанных синапсами и обеспечивающих проведение нервного импульса к рабочему органу. Простая рефлекторная дуга состоит из двух нейронов – чувствительного и двигательного. В сложной рефлекторной дуге между ними включены вставочные, или ассоциативные нейроны. Возбуждение по рефлекторной дуге передается только в одном направлении.
| | | следующая лекция ==> | |
| Нервные окончания | | | Поверхность в пространстве |
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Температурный анализатор является частью соматосенсорного анализатора, хотя его можно рассматривать и как самостоятельный вид анализатора.
Температурная сенсорная система (температурный анализатор) у теплокровных животных предназначена для оценки температуры внешней и внутренней среды организма, на основе которой производится поддержание температурного гомеостаза ядра и, в определенной степени, оболочки тела.
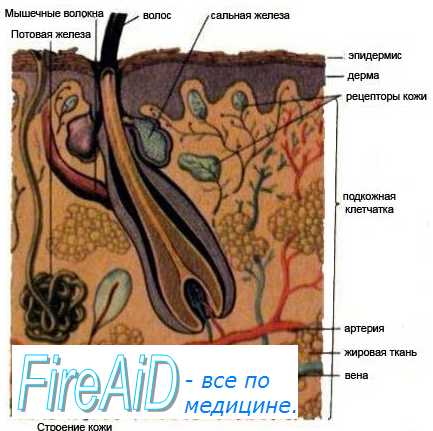
Характеристика периферического отдела температурного анализатора.Терморецепторы расположены на различных участках кожи, слизистых, на роговице глаза, во внутренних органах (в желудке, кишечнике, матке, мочевом пузыре, в дыхательных путях), в скелетных мышцах, кровеносных сосудах, в том числе в артериях, в аортальной и каротидной зонах, во многих крупных венах. Кроме того, терморецепторы имеются в коре больших полушарий, гипоталамусе, в ретикулярной формации ствола мозга, в среднем и спинном мозге. Полагают, что терморецепторы ЦНС – это, скорее всего, нейроны, которые одновременно выполняют роль терморецептора и афферентного нейрона.
Кожные терморецепторы распределены неравномерно. Больше всего терморецепторов на коже лица и шеи. В среднем на 1 мм 2 поверхности кожи приходится I терморецептор.
Все кожные терморецепторы принято подразделять на тепловые и холодовые, а последние, в свою очередь, на собственно холодовые (специфические), реагирующие только на изменение температуры, и тактильно-холодовые, или неспецифические, которые одновременно могут отвечать и на изменение температуры, и на давление. В то же время существует представление о том, что различия температурных ощущений обусловлены различной глубиной залегания в толще кожи единых температурных рецепторов.
Холодовые рецепторы располагаются на глубине 0,17 мм от поверхности кожи, т. е. в базальном слое эпидермиса. Общее число таких рецепторов достигает 250 тысяч. Они реагируют на изменение температуры с коротким латентным периодом. При этом частота генерации потенциалов действия линейно зависит от температуры в пределах от 41° С до 10° С: чем ниже температура, тем выше частота импульсации. Оптимальная чувствительность находится в диапазоне от 15° до 30° С, а по некоторым данным – до 34° С. Следует иметь ввиду, что в некоторых условиях холодовые рецепторы могут быть возбуждены и теплом (выше 45° С). Этим объясняется возникновение острого ощущения холода при быстром погружении в горячую ванну.
Среди холодовых и тепловых рецепторов имеются разные по чувствительности популяции рецепторов: одни реагируют на изменение температуры, равное 0,1° С (высокочувствительные рецепторы), другие – на изменение температуры, равное 1° С (рецепторы средней чувствительности), третьи – на изменение в 10° С (высокопороговые, или рецепторы низкой чувствительности).
В узком нейтральном диапазоне, который соответствует нормальной температуре кожи того или иного представителя гомойотермных (теплокровных) животных в состоянии теплового комфорта (равновесия) тепловые и холодовые рецепторы имеют низкий уровень активности, а даже небольшой сдвиг (на 0,2 о С) в ту или другую сторону воспринимается быстро и точно. Это способствует высокой эффективности терморегуляции.
При очень высоких температурах многие терморецепторы сигнализируют также острую боль.
Проводниковый и корковый отделы температурного анализатора.Информация от кожных терморецепторов к таламусу идет по спинно-таламическому пути, описание которого было дано выше при изложении соматосенсорной системы. При этом часть импульсации идет по афферентным волокнам группы А-дельта, а часть – по афферентным волокнам группы С. Поэтому к таламусу и к коре больших полушарий они доходят с разной скоростью (но не более 30 м/с). Предполагают, что импульсы от холодовых рецепторов идут по волокнам группы А-дельта, а от тепловых рецепторов – по волокнам группы С.
Часть информации от кожных терморецепторов достигает вентробазальных ядер таламуса, часть – вентральных неспецифических ядер. Кроме того, импульсация от терморецепторов доходит до нейронов ствола мозга, а также до нейронов гипоталамуса, где находятся высшие вегетативные центры терморегуляции.
Нейроны таламуса, воспринимающие информацию от кожных терморецепторов, лишь частично дают проекции в соматосенсорную зону коры. При этом на уровне таламических ядер и нейронов коры отсутствует точное представительство терморецепторов поверхности тела, хотя известно, что рецептивные поля большинства терморецепторов локальны.
Восприятие температуры. Информация от таламических нейронов, воспринимающих сигналы терморецепторов кожных покровов и слизистых оболочек, а также от терморецепторов внутренних органов, частично поступает в первую сенсомоторную зону коры больших полушарий (S-1). Вместе с лимбической системой этот поток импульсов обеспечивает формирование теплоощущения (тепло, холодно, температурный комфорт, температурный дискомфорт). Окружающая среда всегда имеет некоторую температуру, поэтому деятельность терморецепторов характеризуется отсчетом температуры относительно нормальной температуры тела: все, что оказывается ниже этой температуры, кажется холодным, все, что выше – теплым и горячим. Таким образом, терморецепторы обнаруживают тепловое излучение только косвенно, по его влиянию на температуру кожи.
Важным фактором, определяющим температурные ощущения человека, является абсолютное значение температуры. В то же время начальная интенсивность температурных ощущений зависит от разницы температуры кожи и температуры действующего раздражителя, его площади и места приложения. Так, если руку держали в воде температуры 27° С, то в первый момент при переносе руки в воду, нагретую до 25°С, она кажется холодной, однакоуже через несколько секунд становится возможной истинная оценка абсолютной температуры воды.
Известен также метод количественной термоэстезиометрии, позволяющий определить количество холодовых и тепловых точек на поверхности кожи. С этой целью применяется термоэстезиметр – небольшой конусообразный стеклянный сосуд (он предварительно заполняется льдом или горячей водой с температурой 50 о С), широкая часть которого закрыта пробкой, а в вершину впаян стержень из металла с высокой теплопроводностью. При исследовании на участок кожи накладывается бумажный трафарет с квадратным отверстием площадью 1 см 2 , к которому прикасаются стержнем термоэстезиметра. Подсчет холодовых или тепловых точек производят по зигзагообразной линии в квадрате трафарета на основании ответов исследуемого на 50 касаний, которые наносятся, начиная с левого верхнего угла трафарета.
Для оценки процессов адаптации терморецепторов кожи опускают кисть руки в горячую (40 о С) или холодную (10 о С) воду и определяют время адаптации терморецепторов, т.е. время, в течение которого ощущение тепла или холода ослабевает.
Адаптацию к теплу и холоду, а также относительность температурных ощущений (явление контрастности) можно наблюдать в следующем опыте: в три сосуда наливают воду с температурой 15° С, 30 о С и 45° С. Одну руку опускают в воду с температурой 15° С, другую – в воду с температурой 45° С. Подержав руки некоторое время в разных сосудах, их одновременно погружают сосуд, содержащий воду, нагретую до 30 °С. При этом одной рукой вода ощущается как горячая, а другой – как холодная. Этот опыт показывает, что на температурные ощущения влияет не только температура сама по себе, но и адаптация к ней. Этот же эксперимент можно провести в ином порядке, например, опускают обе руки (или кончики пальцев) в воду с температурой 25 о С. Убедившись, что ощущение в обеих руках одинаково, одну руку переносят в воду с температурой 40 о С, другую – с температурой 10 о С. Через несколько минут обе руки переносят в воду с температурой 25 о С. При этом возникает ощущение контраста (рука, находившаяся в холодной воде, ощущает тепло, другая рука, находившаяся в горячей воде, ощущает холод).
Восприятие теплового (инфракрасного) излучения. Некоторые животные могут непосредственно воспринимать инфракрасное излучение с помощью особых сенсорных систем, реагирующих на радиацию этого типа. Такими специализированными рецепторами инфракрасных лучей являются лицевые ямки на голове некоторых змей. Например, у гремучей змеи с каждой стороны головы между ноздрей и глазом расположено по одной ямке. Змеи, обладающие лицевыми ямками, питаются преимущественно теплокровной добычей, и, если к голове змеи приблизить предмет, температура которого отличается от температуры окружающей среды, это сразу вызывает повышение активности нерва, иннервирующего рецепторы ямки. Лицевые ямки очень чувствительны и реагируют на изменение температуры всего на 0,1 С. Кроме того, лицевые ямки, по-видимому, обеспечивают стереоскопическое восприятие, что способствует точной локализации добычи.
Температурная чувствительность. Тепловые рецепторы. Холодовые рецепторы. Температурное восприятие.
Ощущения тепла или холода, вызванные прикосновением к телу человека нагретых или холодных предметов, теплого или охлажденного воздуха, возникают вследствие раздражения одной из двух разновидностей температурных рецепторов, представленных в коже свободными нервными окончаниями. Среди них различают тепловые и холодовые рецепторы, которым соответствуют две субмодальности температурного восприятия внешних воздействий. Температурные рецепторы распределены в коже неравномерно и образуют специфические тепловые и холодовые точки площадью около 1 мм2 и менее, с наибольшей плотностью на лице и ладонях, причем Холодовых точек примерно в десять раз больше, чем тепловых.
Импульсная активность терморецепторов возрастает в зависимости от направления и скорости изменений температуры поверхности кожи, составляющей в условиях температурного комфорта около 33 °С. При ее снижении растет активность Холодовых рецепторов с максимумом в диапазоне от 26 до 17 °С, что субъективно расценивается как действие на кожу холода. При повышении температуры кожи от 34 до 45 С увеличивается импульсная активность тепловых рецепторов, что субъективно ощущается как возрастающее по интенсивности действие на кожу тепла. При уменьшении температуры кожи ниже 17 °С и увеличении выше 45 °С к температурным ощущениям прибавляются болевые.
Тоническая активность температурных рецепторов при неизменной температуре сохраняется в течение некоторого времени, а затем постепенно уменьшается. В таком случае, как и при медленных изменениях действующей на рецепторы температуры, происходит их адаптация, в связи с которой постепенно ослабевают ощущения тепла или холода. Но в ответ на резкие ступенчатые изменения действующей температуры (температурный контраст) возникает залповая физическая активность терморецепторов, которая отражает произошедшие изменения температуры. Наличие адаптации терморецепторов наряду с их способностью реагировать залповой активностью на контрастные изменения действующей температуры проявляются в уменьшении порога тепловой чувствительности при пониженной температуре кожи и в уменьшении порога холодовой чувствительности при повышении ее температуры.
Спиноталамический путь ведет к задним вентролатеральным ядрам таламуса, на нейронах которых конвергируют проводящие пути температурной и тактильной чувствительности. Благодаря такой конвергенции у человека проявляется способность устанавливать, на какую часть поверхности тела подействовал температурный стимул. Спиноретикулярный тракт служит для проведения возбуждения от терморецепторов к ретикулярной формации и принадлежит к неспецифической части сенсорной системы. Ретикулярная формация вместе с неспецифическими ядрами таламуса активирует одновременно многие регионы коры, определяя общую активность мозга и уровень бодрствования. Поэтому повышение притока афферентной информации к ней от терморецепторов, например во время приема холодного душа, оказывает на человека тонизирующее влияние.
терморецепторы (термо- + рецептор) - см. Рецепторы температурные.
1) Орфографическая запись слова: терморецепторы2) Ударение в слове: терморец`епторы3) Деление слова на слоги (перенос слова): терморецепторы4) Фонетиче. смотреть
Этимология. Происходит от греч. thermos - теплый. Категория. Вид рецепторов. Специфика. Рецепторы, расположенные на кожной поверхности и во внутренних органах и служащие для контроля за температурными изменениями. Выделяют холодовые, с оптимумом чувствительности 28-38 градусов, и тепловые, с оптимумом 35-43 градуса. При этом кожные холодовые рецепторы представлены значительно большим количеством, чем тепловые, и располагаются они более поверхностно. Наибольшая плотность распределения терморецепторов на лице, наименьшая - на подошве ног. Существуют особые терморецепторы, контролирующие температуру крови, расположенные в гипоталамической области мозга. смотреть
термоцепторы, нервные окончания (Рецепторы) в различных тканях и органах, специфически реагирующие на изменения температуры тела изменением час. смотреть
Терморецепторы - рецепторы, расположенные на кожной поверхности и во внутренних органах и служащие для контроля за температурными изменениями. Выделяют холодовые, с оптимумом чувствительности 28 - 38 градусов, и тепловые, с оптимумом 35 - 43 градуса. При этом кожные холодовые рецепторы представлены значительно большим количеством, чем тепловые, и располагаются они более поверхностно. Наибольшая плотность распределения терморецепторов на лице, наименьшая - на подошве ног. Существуют особые терморецепторы, контролирующие температуру крови, расположенные в гипоталамической области мозга. смотреть
Ударение в слове: терморец`епторыУдарение падает на букву: еБезударные гласные в слове: терморец`епторы
ТЕРМОРЕЦЕПТОРЫ, в анатомии - чувствительные нервные окончания, расположенные в коже и в глубине тканей тела, которые фиксируют изменения температуры и . смотреть
ТЕРМОРЕЦЕПТОРЫ (от термо . и рецепторы), чувствительные нервные окончания, реагирующие на изменения температуры тела и окружающей среды.
ТЕРМОРЕЦЕПТОРЫ (от термо. и рецепторы) - чувствительные нервные окончания, реагирующие на изменения температуры тела и окружающей среды.
(от термо. и рецепторы), чувствительные нерв. окончания, реагирующие на изменения темп-ры тела и окружающей среды.
терморец'епторы и термоц'епторы, -ов, ед. ч. -тор, -а
- (от термо. и рецепторы) - чувствительные нервныеокончания, реагирующие на изменения температуры тела и окружающей среды.
(Термо- + рецептор)см. Рецепторы температурные.
Рецепторы, реагирующие на изменение температуры окружающей среды и тела человека.
терморецепторы терморец`епторы и термоц`епторы, -ов, ед. -тор, -а
терморецепторы терморец`епторы, -ов, ед. -тор, -а
(термо- + рецептор) см. Рецепторы температурные.
ТЕРМОРЕЦЕПТОРЫ (от термо . и рецепторы), чувствительные нервные окончания, реагирующие на изменения температуры тела и окружающей среды.
ТЕРМОРЕЦЕПТОРЫ (от термо. и рецепторы), чувствительные нервные окончания, реагирующие на изменения температуры тела и окружающей среды.
Представляют собой терминальные разветвления дендритов нейроцита. КЛАССИФИКАЦИЯ. Существует несколько принципов классификации рецепторных нервных окончаний.
1. По месту восприятия раздражителя.Рецепторные нервные окончания делятся на три группы: экстерорецепторы,воспринимающие раздражение из внешней среды; интерорецепторы,служащие для восприятия раздражений из внутренней среды организма; проприорецепторы,воспринимающие информацию от опорно-двигательного аппарата.
2. В зависимости от специфичности раздражения, воспринимаемого рецептором.Выделяют: механорецепторы,воспринимающие механические раздражители, перемещения частей тела; хеморецепторывоспринимают химические раздражители; терморецепторыулавливают изменения температуры, а ноцирецепторывоспринимают чувство боли.
3. По способу восприятия раздражителявыделяют контактные рецепторы,приходящие в состояние возбуждения при непосредственном воздействии на участок тела, и дистантные рецепторы,воспринимающие раздражитель, удаленный от организма (рецепторные клетки сетчатки глаза, органа слуха, обоняния).
4. Морфологическая классификация.В зависимости от строения все рецепторы делят на свободныеи несвободные.Свободные рецепторные нервные окончания состоят только из конечных ветвлений дендрита чувствительного нейрона, а несвободные нервные окончания кроме терминалей нервного отростка имеют также и клетки олигодендроглии (леммоци-ты), которые окружают терминали дендрита и участвуют в восприятии раздражения. В свою очередь, несвободные нервные окончания делятся на неинкапсулированные(не окруженные по периферии соединительнотканной капсулой) и инкапсулированные, имеющие такую капсулу. Свободные нервные окончания воспринимают в основном болевые раздражения. Большинство несвободных нервных окончаний являются механорецепто-рами. В последнее время, однако, выдвигается небеспочвенная точка зрения, что не существует разделения рецепторов в зависимости от вида воспринимаемого раздражения, все рецепторы способны воспринимать раздражители любой модальности, а характер ощущения зависит от силы раздражителя.
МОРФОЛОГИЯ РЕЦЕПТОРОВ. 1. Свободные нервные окончания.Внаибольшем количестве представлены в коже. Это механорецепторы на волосяных фолликулах, ноцицептивные (воспринимающие болевые раздражители) нервные окончания в эпидермисе (рис. 14.3). Их много также в многослойном плоском неороговевающем эпителии, серозной оболочке. В эпидермисе они представлены древовидными ветвлениями дендритов псевдоуниполярных нейронов спинальпых ганглиев.
2. Несвободные неинкапсулированные нервные окончанияпредставлены осязательными дисками Меркеля,а также нервными окончаниями соединительной ткани. Особенно много их в дерме. Осязательные диски Меркеля (рис. 14.3 б) состоят из отростка нервной клетки, который закан- чивается расширением в виде П. диска. Этот диск образует синапс с клеткой Меркеля, которая лежит в эпидермисе. В цитоплазме клетки Меркеля есть секреторные гранулы с нейромедиатором. Механическое раздражение вызывает выделение гранул из клеток Меркеля, их содержимое ведет к деполяризации отростка нейроцита.
 |
Несвободные неинкапсули-рованные окончания в соединительной ткани построены следующим образом. Осевой цилиндр освобождается от миелина и на значительном расстоянии окружается глиальными клетками, тесно с ними контактируя. Очень часто на поперечном разрезе видна билатеральная симметрия таких окончаний.
3. Несвободные инкапсулированные нервные окончанияпостроены по общему принципу. К этим окончаниям относятся нервные окончания в соединительной и мышечных тканях. Есть следующие разновидности этих окончаний: пластинчатые тельца Фатер-Пачини, осязательные тельца Мейснера, концевые колбы Краузе, генитальные тельца Догеля, тельца Руффини, нервно-мышечные и нервно-сухожильные веретенаи др.
 |
Наиболее распространены пластинчатые тельца Фатер-Пачини.Они встречаются в коже, молочной железе, в брыжейке, во внутреннил органах, около кровеносных сосудов, около суставов. Это крупные образования диаметром от 1 до 5 мм (рис. 14.4, 14.5). Имеют овальную форму и состоят из соединительнотканной капсулы, терминалей дендрита псевдоуниполярного нейрона и нейролеммоцитов (олигодендроглии). Дендрит при подходе ккапсуле теряет миелиновую оболочку и со всех сторон окружается нейролем-моцитами. Они формируют так называемую внутреннюю колбу.Эта колба снаружи покрыта соединительнотканной капсулой, которая часто называется наружной колбой.Капсула состой! из послойно параллельно лежащих кол-лагеновых волокон (образуют от 10 до 60 слоев) и клеток фиброцитов. В наружной капсуле встречаются кровеносные сосуды. Между наружной и внутренней колбами лежат специализированные отростчатые олигодендроглиоци-ты, контактирующие с осевым цилиндром. При давлении на тельце механическое воздействие во много раз усиливается слоями наружной колбы, что делает этот рецептор очень чувствительным. Давление смещает наружную колбу по отношению к внутренней. При этом раздражаются отростчатые олигоденд-роциты, передающие возбуждение на дендрит.
 |
В сосочковом слое дермы обнаруживаются осязательные тельца Мейс-нера.Они являются механорецепторами и по размеру меньше телец Фа-тер-Пачини (50—140 мкм). Имеют овальную форму (рис. 14.3 и 14.6). Снаружи находится очень тонкая слоистая капсула — наружная колба.Дендрит псевдоуниполярного нейрона теряет миелиновую оболочку, разветвляется, и его ветви входят внутрь капсулы по спирали. Перпендикулярно к ним лежат глиальные клетки, которые вместе с терминалами дендритов образуют внутреннюю колбу.Незначительная деформация капсулы передается глиоцитам, которые имеют синаптическую связь с дендритом.
Концевые колбы Краузеявляются барорецепторами и терморецепторами. Они лежат в дерме кожи, слизистых оболочках. Имеют небольшие (40—150 мкм) размеры. Также состоят из наружной капсулы и внутренней колбы. Внутренняя колба образована плоскими глиоцитами, между которыми проходят, формируя своеобразный клубочек, тонкие ветви дендрита. Наружная капсула очень тонкая.
Генитальные тельца Догелянаходятся в особо чувствительных областях кожи, в первую очередь, в области наружных половых органов, коже молочных желез. Они похожи по строению на колбы Краузе, но в в отличие от них в тельце входят несколько отростков от нескольких нейро-цитов. Поэтому раздражение генитального тельца вызывает сильную иррадиацию возбуждения.
Тельца Руффининаходятся в соединительной ткани кожи и в капсулах суставов. Воспринимают чувство давления. Имеют вид верстеновид-ных образований длиной до 2 мм. Осевой цилиндр во внутренней колбе разветвляется с образованием большого количества ветвей с булавовидными утолщениями на конце. Капсула хорошо выражена.
 |
В гладкой мышечной тканичувствительные нервные окончания также инкапсулированы, они контактируют с группой гладких миоцитов.
В скелетной мышечной ткани чувствительные нервные окончания называются нервно-мышечными веретенами.Представляют собой инкапсулированные нервные окончания (рис. 14.7, 14.8). Наружная соединительнотканная капсула нервно-мышечного веретена окружает несколько тонких так называемых интрафузальных мышечных волокон. В отличие от обычных мышечных волокон, лежащих снаружи и называемых экстрафузаль-ными,интрафузальные волокна тонкие, содержат мало миофибрилл и имеют светлую цитоплазму. Различают два вида интрафузальных мышечных волокон (рис. 14.8).1. ЯС-волокна.Ядра этих волокон лежат в центральной части мышечного волокна, образуя скопление в виде ядерной сумки(сокращенно ЯС). В месте расположения ядер волокно резко расширяется.
2. ЯЦ-волокна.Эти волокна имеют равномерную толщину, а ядра лежат но всей длине волокна в его центре, формируя ядерную цепь.
Вокруг данных двух видов интрафузальных волокон в их центральной части образуются специфические синапсы дендритов чувствительных нейронов в виде:
1) аннулоспиральных (кольцеспиральных)окончаний, в которых отростки нервных клеток закручены вокруг центральной части интрафузалыю-го волокна по спирали и на большом протяжении вступают в синаптичес-кую связь с ним; аннулосниральные окончания имеются как на ЯС-, так и на ЯЦ-волокнах.2) гроздьевидных окончаний,которые находятся только на ЯЦ-волокнах. При этом они формируются не в центральной части, а на периферии волокна.
 |
На интрафузальных волокнах имеются также двигательные нервные окончания,которые представлены аксонами у-мотонейроновпередних рогов спинного мозга. Они регулируют длину интрафузальных волокон и поддерживают их тонус. Все свободное пространство между мышечными волокнами заполнено жидкостью и ограничено тонкой капсулой. Всякое изменение тонуса мышцы ведет к изменению давления жидкости в полости капсулы. При этом давление передается на дендриты. Аннулосниральные окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, а гроздьевидные — только на изменение длины. Благодаря нервно-мышечным веретенам организм по-
стоянно получает информацию о степени сокращения мышц, что формирует представление о положении тела в пространстве.
МЕЖНЕЙРОННЫЕ СИНАПСЫ
Это особый вид нервных окончаний, когда разветвления отростков одних нервных клеток заканчиваются на других нервных клетках. При помощи синапсов возбуждение передается с одной нервной клетки на другую.
Классификация синапсов.Существует несколько подходов к классификации синапсов.
 |
i. По механизму передачи нервного импульса.Синапсы делятся на химические, электрическиеи смешанные. В химических синапсахвозбуждение передается при помощи химического вещества — нейро-медиатора.Эти синапсы являются наиболее распространенными в нервной системе высших животных. В электрических синапсахпотенциал действия передается прямо с мембраны одного нейрона на другой. Смешанные синапсыпредставляют собой сочетание признаков и химического, и электрического синапсов.
2. Морфологическая классификация синапсов.Учитывает особенности контактирующих участков иейроцитов. Различают аксо-соматичес-кие, аксо-дендритические, аксо-аксональные, дендро-дендричес-кие, сомато-соматическиесинапсы.
3. Физиологическая классификация. По вызываемому эффектуна нервную клетку синапсы делятся на возбуждающиеи тормозные.
4. Медиаторная классификация синапсов.По химическому типу медиатора есть синапсы холинергические, аминергические (адренергические, серотонинергические, дофаминергические); пуринергичес-кие, аминокислотные (медиаторами являются аминокислоты: ГАМК, глицин, глутамат, аспартати т.д.), пептидергические(см. ме-диаторную классификацию нейроцитов).
СТРОЕНИЕ СИНАПСОВ. Любой синапс состоит из трех частей: пре-синаптического полюса с пресинаптической мембраной, синапти-ческой щелии постсинаптического полюса с постсинаптической мембраной.
 |
Электрические синапсы.Эти синапсы построены по типу нексусов:две мембраны (пре- и постсинаптическая) соседних нейронов тесно сближаются друг с другом до расстояния в 2 нм, и это место контакта пронизано многочисленными коннексонами.Следовательно, синаптическая щель в электрическом синапсе практически отстутствует. Коннексоны представляют собой своеобразную пору через обе мембраны, которая по краям ограничена особыми белковыми молекулами коннексинами.Коннексоны пропускают не только ионы щелочных металлов, играющих важную роль в формировании электрических потенциалов, но и молекулы с ММ 1000—2000. Поэтому кроме электрического сопряжения коннексоны дают возможность нейронам обмениваться метаболитами. В отличие от химических синапсов, в которых проведение сигнала несколько задерживается, в электрических синапсах импульс проводится практически без задержки и в обе стороны. Значение электрических синапсов неизвестно. Предполагают, что оно связано с необходимостью быстрого сопряжения нервных клеток.
Химические синапсы.В отличие от электрических, химические синапсы передают нервные импульсы только в одном направлении и с задержкой (синаптическая задержка).Это наиболее распространенный у млекопитающих тин синапсов. Химические синапсы имеют все три отчетливо выраженные составные компоненты: нресинаптический и ностсинаптический полюсы и синаптичес-кую щель (рис 14.9) (в световом микроскопе синапсы видны в виде пугов-чатых утолщений на нейроцитах, см. рис. 13.1).
В пресинаптическом полюсенаходятся пресинаптические пузырьки с медиатором, митохондрии, агранулярная ЭПС, нейротрубочки и нейрофи-ламенты. Синаптические пузырьки имеют различное строение в зависимости от содержащегося в них медиатора. Так, пузырьки с ацетилхолином имеют мелкие размеры и электронно прозрачные. Синаптические пузырьки с но-радреналином крупнее и имеют в центре электронноплотную часть. Содержащие пептиды пузырьки имеют крупные размеры, плотную сердцевину и окружены периферическим светлым ободком.
Читайте также:
