
Чувствительным нервным окончанием ответственным за термовосприятие является
г. мотонейронов спинного мозга
Нейрофибриллы в нервных клетках соответствуют
а. продольным канальцам ЭПС
б. пучкам нейрофиламентов
в. миозиновым нитям
г. пучкам нейротрубочек
Нейроциты, аксоны которых образуют двигательные окончания в гладкой мышечной ткани, располагаются в
а. передних рогах спинного мозга
б. боковых рогах спинного мозга
в. вегетативных ганглиях
г. спинномозговых ганглиях
Нейроциты, дендриты которого образуют мышечные веретена, по функции относятся к
в. ассоциативному тормозному
По аксону транспортируется
в. белковых молекул
В аксоне присутствуют
г. тигроидное вещество
Оболочки безмиелиновых нервных волокон образованы
а. плазмолеммой шванновских клеток (нейролеммоцитов)
в. цитоплазмой шванновских клеток
г. спирально закрученной мембраной аксона
Оболочки миелиновых нервных волокон образованы
г. клетками микроглии
В миелиновом волокне ЦНС различают
а. перехват Ранвье
б. осевой цилиндр
г. шванновские клетки
Миелиновая оболочка нервных волокон образована
а. плазмолеммой шванновских клеток (нейролеммоцитов)
в. цитоплазмой шванновских клеток
г. спирально закрученной мембраной аксона
В процессе дегенерации и регенерации нервных волокон основная роль принадлежит клеткам
Какие клетки синтезируют белки миелина
Основным химическим компонентом миелина является
Чувствительное нервное окончание, ответственное за термовосприятие
а. пластинчатое тельце Пачини
б. осязательное тельце Майсснера
в. свободное нервное окончание
г. тельце Руффини
В состав пластинчатого тельца (барро-рецептора) входят
в. коллагеновые волокна
г. эластические волокна
Рецепторные нервные окончания располагаются в
б. соединительной ткани
Какие клетки в эпидермисе кожи образуют тактильные рецепторы
б. клетки Лангерганса
в. клетки Меркеля в комплексе с нервной терминалью
Свободные нервные окончания имеют характерные признаки
а. чувствительное, содержит поперечно-полосатые мышечные волокна
б. чувствительное , состоит только из ветвлений осевого цилиндра
в. двигательное, нервно-мышечный синапс
г. чувствительное, имеет соединительную капсулу
Какие из перечисленных нервных окончаний относят к несвободным
а. тельце Пачини (баррорецептор)
б. тельце Мейснера (осязательное)
в. тактильные клетки Меркеля в комплексе с нервной терминалью
Нервное окончание – пластинчатое тельце имеет характерные признаки
а. чувствительное, содержит поперечно-полосатые мышечные волокна
б. чувствительное, состоит только из ветвлений осевого цилиндра
в. двигательное, нервно-мышечный синапс
г. чувствительное, имеет соединительную капсулу
Медиатор в нервно-мышечном синапсе скелетной мышцы
Для нервно-мышечного синапса характерно
а. синаптические пузырьки с ацетилхолином
б. постсинаптическая мембрана с холинорецепторами
в. нервная терминаль окружена шванновской клеткой
г. синаптические везикулы выходят в синаптическую щель
Однонаправленное проведение нервного импульса в синапсе определяется
а. системой нейрофибрилл
б. наличием митохондрий
в. аксоплазматическим током веществ
г. наличием рецепторного белка на постсинаптической мембране
Для нейромышечного синапса характерен медиатор
Нервно-мышечное окончание имеет характерные признаки
а. чувствительное, содержит поперечно-полосатые мышечные волокна
б. чувствительное, состоит только из ветвлений осевого цилиндра
в. двигательное, нервно-мышечный синапс
г. чувствительное, имеет соединительную капсулу
После травматического сдавления конечности в ее нервном стволе при дегенерации имеются проявления
а. распада окончаний нервных волокон
б. разрушения миелина
в. гибели шванновских клеток в дистальном отрезке
г. фагоцитоза фрагментов поврежденных нервных волокон
Спинно-мозговой канал, желудочки мозга выстланы
г. клетками микроглии
Астроциты выполняют функции
в. участвуют в обмене медиаторов
г. участвуют в водно-солевом обмене
Поддерживающий аппарат серого вещества ЦНС образован
б. протоплазматическими астроцитами
в. волокнистыми астроцитами
Для клеток микроглии характерно
а. многочисленные лизосомы
б. участие в иммунном ответе в мозге
в. способность к обновлению
г. участие в транспорте глюкозы
Какие процессы происходят после локальной компрессии (сдавления) периферического нерва
а. разрушение осевых цилиндров
б. распад миелина
в. прорастание аксонов из центрального отрезка в периферический
г. ориентация регенерирующих аксонов по цепочкам из шванновских клеток
Эталоны ответов:1бг, 2а, 3в, 4а, 5абв, 6бг, 7в, 8г, 9аб, 10абв, 11абвг, 12в, 13абвг, 14а, 15в, 16а, 17в, 18в, 19абвг, 20абвг, 21в, 22б, 23абв, 24г, 25в, 26абв, 27г, 28а, 29в, 30абг, 31а, 32абвг, 33б, 34абв, 35абвг.
Область, раздражение которой приводит к возбуждению чувствительной единицы, называют рецепторным полем. Чем больше размер рецепторного поля, тем меньшей остротой сенсорной чувствительности обладает данная область: например, в верхней части руки рецепторные поля занимают площадь 2 см 2 , в области запястья — 1 см 2 , на подушечках пальцев — 5 мм 2 .
Чувствительные единицы переплетаются между собой, за счет чего становится возможным одновременное восприятие одним участком кожи разных видов чувствительности.
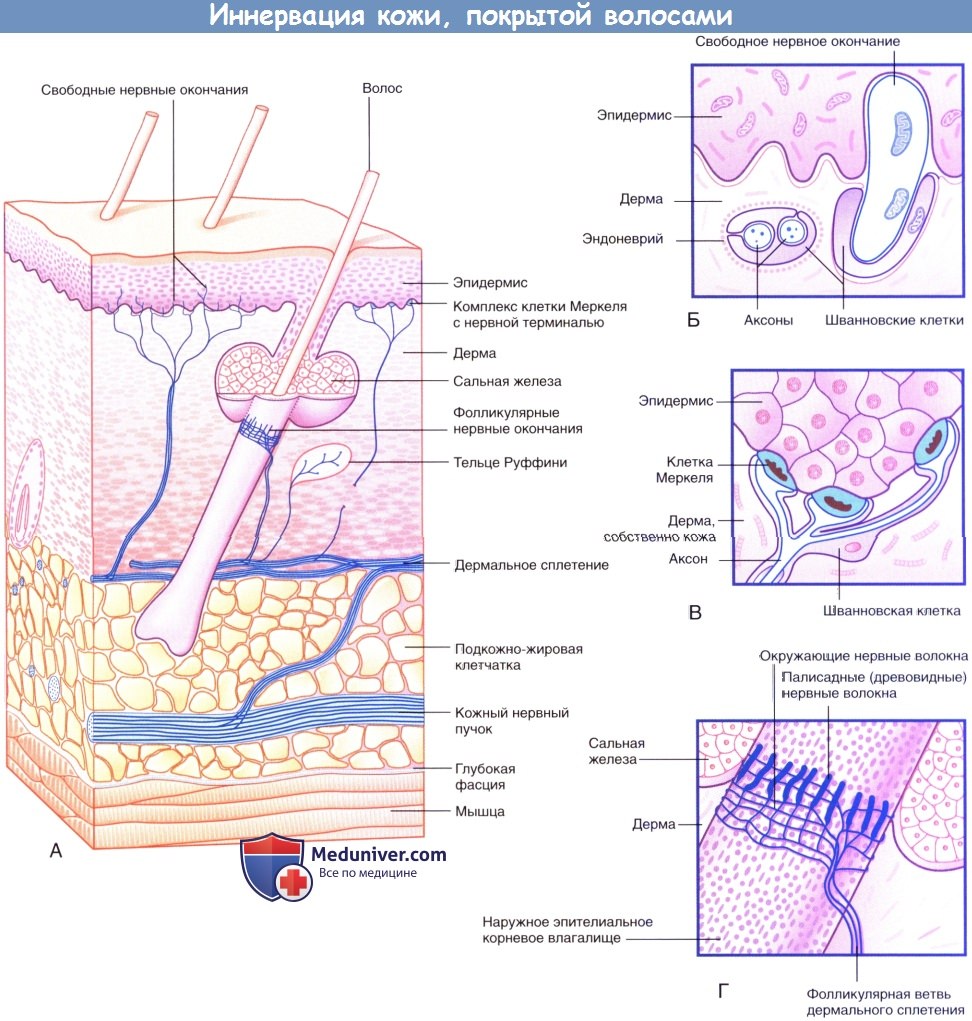
Иннервация кожи, покрытой волосами.
(А) Три морфологических типа чувствительных нервных окончаний в коже, покрытой волосами.
(Б) Свободные нервные окончания в базальном слое эпидермиса.
(В) Комплекс клетки Меркеля с нервной терминалью.
(Г) Палисадные и циркулярные нервные окончания на поверхности наружного корневого влагалища волоса.
б) Нервные окончания:
1. Свободные нервные окончания. По мере приближения к поверхности кожи многие чувствительные нервные волокна утрачивают периневральную, а затем и миелиновую оболочку (в случае ее наличия). Впоследствии нервные волокна разветвляются и формируют субэпидермальное нервное сплетение. Аксон освобождается от оболочек, сформированных шванновскими клетками, что позволяет ему, разветвляясь между коллагеновыми пучками дермы, образовывать дермальные нервные окончания, а внутри эпидермиса — эпидермальные нервные окончания.
2. Фолликулярные нервные окончания. Нервные окончания волосяного фолликула представлены палисадными нервными волокнами, образованными обнаженными терминалями миелинизированных нервных волокон, расположенными на поверхности наружного корневого влагалища волосяных фолликулов ниже уровня сальных желез, а также циркулярными нервными окончаниями. Каждая фолликулярная единица иннервирует несколько волосяных фолликулов и образует множественные перекресты. Фолликулярные единицы — быстро адаптирующиеся: они возбуждаются при изменении положения волос, однако при сохранении этого положения возбуждения не происходит. Человек, одеваясь, чувствует давление одежды, но затем за счет быстрой адаптации вскоре перестает ощущать ее прикосновение. Иннервация волос у других млекопитающих организована сложнее. Иннервация волосяных фолликулов осуществляется тремя типами механорецепторов, каждый из которых передает информацию определенным структурам мозга, что свидетельствует о важности выполняемой ими чувствительной функции.
3. Комплексы клетки Меркеля с нервной терминалью. Нервная терминаль, расширяясь в области базального слоя эпидермальных гребешков и бороздок, образует комплекс с осязательным тельцем овальной формы — клеткой Меркеля. Комплексы клетки Меркеля с нервной терминалью — медленно адаптирующиеся. В ответ на продолжительное давление (например, при удержании ручки или ношении очков) эти комплексы непрерывно генерируют нервные импульсы. Комплексы клетки Меркеля с нервной терминалью особенно хорошо распознают края удерживаемых в руке предметов.
4. Инкапсулированные нервные окончания. Капсулы описанных ниже свободных нервных окончаний состоят из трех слоев: наружный слой представлен соединительной тканью, средний — периневральным эпителием, а внутренний — видоизмененными шванновскими клетками (телоглией). Инкапсулированные нервные окончания являются механорецепторами, преобразующими механическое воздействие в нервный импульс.
• Тельца Мейснера в большом количестве находятся в подушечках пальцев и расположены вблизи бороздок эпидермиса. Тельца представляют собой клетки овальной формы, внутри которых аксоны располагаются зигзагообразно между уплощенными клетками телоглии. Тельца Мейснера— быстро адаптирующиеся, вместе с медленно адаптирующимися комплексами клетки Меркеля с нервной терминалью они обеспечивают точное восприятие текстур (например, текстуры ткани одежды или поверхности дерева), а также рельефных поверхностей (например, шрифта Брайля). Такие кожные рецепторы способны воспринимать изменение рельефа поверхности даже на высоту 5 нм.
• Тельца Руффини присутствуют как на гладкой коже, лишенной волос, так и на коже с волосами. Они воспринимают плавные скользящие касательные прикосновения и являются медленно адаптирующимися. Внутреннее строение телец сходно со строением сухожильных органов Гольджи: аксоны образуют разветвления в центральной части телец, представленной коллагеновыми волокнами.
• Тельца Пачини по величине соответствуют размерам рисового зерна. В области кисти имеется около 300 телец, которые преимущественно сконцентрированы на боковых участках пальцев и ладони. Тельца Пачини расположены подкожно, близко к надкостнице. Несколько слоев периневрального эпителия внутри соединительнотканной капсулы расположены овально и по форме напоминают луковицу в разрезе. В центральной части тельца Пачини несколько пластинок телоглии окружают единичный аксон, который, попадая в тельце, утрачивает миелиновую оболочку. Тельца Пачини — быстро адаптирующиеся рецепторы преимущественно вибрационной чувствительности. Эти структуры особенно восприимчивы к вибрации костной ткани: большое количество телец расположено в надкостнице длинных трубчатых костей.
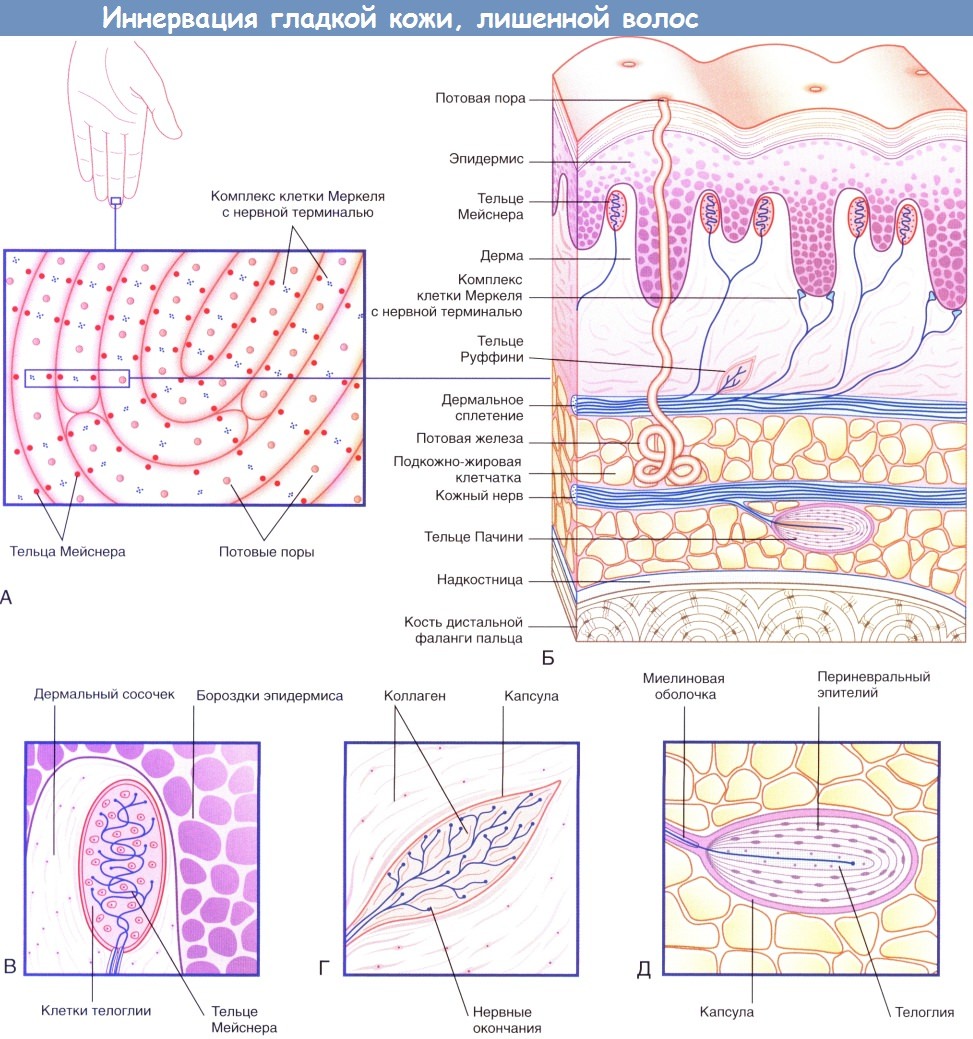
Иннервация гладкой кожи, лишенной волос.
(А) На подушечках пальцев располагаются нервные окончания двух видов.
(Б) На схеме строения участка кожи с изображения (А) представлены четыре типа чувствительных нервных окончаний.
(В) Тельца Мейснера.
(Г) Тельца Руффини.
(Д) Тельца Пачини.
Специалисты по физиологии чувствительности выделяют следующие виды рецепторов, локализованных в коже пальцев.
• Комплексы клетки Меркеля с нервной терминалью — медленно адаптирующиеся рецепторы I типа (MAP I).
• Тельца Мейснера — быстро адаптирующиеся рецепторы I типа (БАР I).
• Тельца Руффини — медленно адаптирующиеся рецепторы II типа (MAP II).
• Тельца Пачини — быстро адаптирующиеся рецепторы II типа (БАР II).
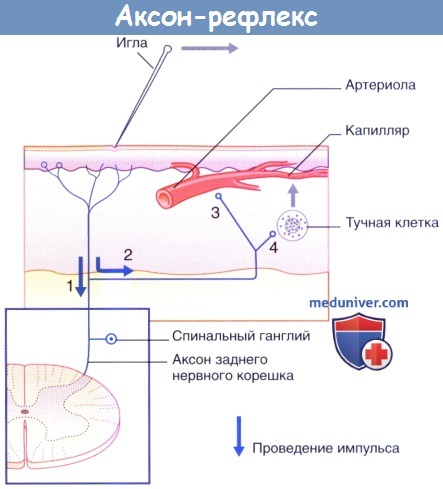
1. Полимодальные ноцицепторы преобразуют действие болевого раздражителя в нервные импульсы.
2. Аксоны посылают нервные импульсы в центральную нервную систему не только в обычном ортодромном направлении, но и в противоположном антидромном направлении от мест бифуркации к прилежащим участкам кожи. Ответная реакция ноцицептивных нервных окончаний на антидромную стимуляцию проявляется в высвобождении пептидных веществ, среди которых в большом количестве представлена субстанция Р.
3. Субстанция Р связывается с рецепторами на стенках артериол и вызывает их расширение, что приводит к появлению гиперемии.
4. Кроме того, субстанция Р связывается с рецепторами на поверхности тучных клеток, что приводит к высвобождению из них гистамина. Гистамин увеличивает проницаемость капилляров, за счет чего происходит местное накопление тканевой жидкости, обусловливающее возникновение бледного отечного валика.
д) Резюме. Направляющиеся к коже нервы разветвляются и образуют дермальное нервное сплетение. Чувствительные нервные волокна дермального сплетения разветвляются и перекрывают друг друга. Каждое стволовое нервное волокно и его рецепторы формируют чувствительную единицу. Область, иннервируемую стволовым нервным волокном, называют его рецептивным полем.
К чувствительным единицам со свободными нервными окончаниями относят рецепторы температурной чувствительности, а также механические и температурные рецепторы болевой чувствительности. Рецепторы волосяных фолликулов—быстро адаптирующиеся осязательные механорецепторы, которые активируются только при движении волос. Комплексы клеток Меркеля с нервными терминалями обеспечивают восприятие края предметов, их относят к медленно адаптирующимся.
Редактор: Искандер Милевски. Дата публикации: 13.11.2018
∙ Центральные отрезки нервных волокон на небольшом протяжении и всём протяжении периферического отрезка.
Периферические отрезки нервных волокон на всём протяжении.
2.1.11. Дегенерация нервного ствола сопровождается всеми проявлениями, за исключением:
∙ Распада окончаний нервных волокон.
Гибели швановских клеток в дистальном отделе.
2.1.12. Тормозной нейромедиатор:
2.1.13. Изменение длины мышечных волокон регистрируют:
∙ Осязательные эпителиоциты.Мышечные веретёна.
2.1.14. Чувствительным нервным окончанием, ответственным за термовосприятие является:
∙ Пластинчатое тельце Пачини.
Свободное нервное окончание.
2.1.15. В миелиновом нервном волокне периферической нервной системы присутствует все перечисленное, за исключением:
Нескольких осевых цилиндров.
2.1.16. В безмиелиновом нервном волокне периферической нервной системы присутствует все перечисленное, за исключением:
Одного осевого цилиндра.
∙ Нескольких осевых цилиндров.
2.1.17. Преганглионарные симпатические нейроны спинного мозга образуют ядра в:
2.1.18. Периневрий это:
∙ Слой соединительной ткани вокруг каждого нервного волокна.
∙ Содержит клетки, связанные плотными контактами.
Соединительная ткань, расположенная вокруг пучков нервных волокон.
∙ Контролирует проницаемость и поддерживает гомеостаз эндоневрия.
2.1.19. Какими синапсами заканчиваются преганглионарные волокна, вступая в соответствующий ганглий?
2.1.20. Какие нейроны не содержатся в ганглии интрамуральных сплетений?
2.1.21. Преганглионарные волокна заканчиваются на телах клеток ганглия парасимпатического отдела, образуя синапсы:
2.1.22. Однонаправленное проведение сигнала в области синапса определяется:
∙ Системой нейрофиламентов и нейротрубочек. ∙ Наличием митохондрий.
∙ Направлением аксонного транспорта.
Присутствием рецепторного белка в постсинаптической мембране.
2.1.23. В регенерации нервных волокон основная роль принадлежит:
2.1.24. Нейрон, дендриты которого образуют мышечные веретёна, по функции относится к нейрону:
2.1.25. Выберите верные утверждения, характеризующие оболочки периферических нервов:
Периневрий имеет пластинчатое строение и окружает отдельные пучки нервных волокон.
∙ Эндоневрий содержащих жировые клетки.
∙ Эпиневрий содержит кровеносные сосуды и чувствительные нервные структуры.
∙ Мелкие нервы не имеют периневрия.
2.1.26. Эпиневрий периферического нерва образован:
Волокнистой соединительной тканью.
∙ Ретикулярной соединительной тканью.
2.1.27. Периневрий образован:
Пластами плоского однослойного эпителия на базальной мембране, разделенного тонкими прослойками рыхлой соединительной ткани.
∙ Волокнистой соединительной тканью.
∙ Ретикулярной соединительной тканью.
2.1.28. Что меняется в составе нервных стволов в пожилом возрасте:
∙ Развитие соединительнотканных элементов.
∙ Миелинизация нервных волокон.
2.1.29. Скорость передачи импульса по миелиновым волокнам:
2.1.30. Скорость передачи импульса по безмиелиновым волокнам:
2.1.31. Срединная щель спинного мозга находится:
∙ Сзади между правой и левой половинами.
Спереди между правой и левой половинами.
∙ Делит спинной мозг на вентральную и досальную части. ∙ Проходит через центр спинного мозга.
2.1.32. Срединная соединительнотканная перегородка спинного мозга находится:
Сзади между правой и левой половинами.
∙ Спереди между правой и левой половинами.
∙ Делит спинной мозг на вентральную и досальную части.
∙ Проходит через центр спинного мозга.
2.1.33. Дорсальные рога серого вещества спинного мозга располагаются:
∙ Между срединной щелью и передним канатиком.
∙ Между передним и боковым канатиками.
∙ Между соединительнотканной перегородкой и задним канатиком.
Межу задним и боковым канатиками.
2.1.34. Вентральные рога серого вещества спинного мозга располагаются:
Между срединной щелью и передним канатиком.
Между передним и боковым канатиками.
Между соединительнотканной спайкой и задним канатиком.
Межу задним и боковым канатиками.
Нейроны серого вещества спинного мозга относятся к:
2.1.36. Центральный канал спинного мозга выстлан:
∙ Протоплазматическими астроцитами. ∙ Олигодендроглиоцитами.
2.1.37. Корешковые нейроны серого вещества спинного мозга располагаются в составе:
∙ Передних и задних рогов.Передних и боковых рогов.
∙ В составе всех участков.
∙ В составе задних рогов.
2.1.38. Пучковые клетки серого вещества спинного мозга располагаются в составе:
∙ Передних и задних рогов
∙ Передних и боковых рогов.
∙ В составе всех участков.В составе задних рогов.
2.1.39. Внутренние клетки серого вещества спинного мозга располагаются:
∙ В передних и задних рогах
∙ В передних и боковых рогах.В составе всех участков.
∙ В составе задних рогов.
2.1.40. Центральные ядра симпатического отдела нервной системы спинного мозга располагаются в:
2.1.41. Центральные ядра парасимпатического отдела нервной системы спинного мозга располагаются в:
2.1.42. Чувствительные нейроциты рефлекторных дуг локализуются:
В спинальных ганглиях.
∙ В передних корешках спинного мозга.
∙ В спинном мозге.
∙ В паравертебральных ганглиях.
2.1.43. Двигательные нейроциты рефлекторных дуг спинного мозга образуют ядра:
В передних рогах.
∙ В передних канатиках.
∙ В боковых рогах.
2.1.44. Нейроны серого вещества спинного мозга являются:
2.1.45. В промежуточной зоне серого вещества спинного мозга располагаются ядра:
2.1.46. Где располагаются тела двигательных нейронов, аксоны которых образуют нейромышечный синапс:
∙ Коре больших полушарий.Спинном мозге.
2.1.47. Серое вещество спинного мозга развивается из следующих слоев нервной трубки:
2.1.48. Передние корешки спинного мозга содержат:
∙ В основном безмиелиновые волокна.
2.1.49. Серое вещество спинного мозга содержит нейроны:
2.1.50. Белое вещество спинного мозга состоит из:
Миелиновых нервных волокон.
∙ Тел внутренних (собственных) нейронов.
2.1.51. Сколько оболочек имеет спинной мозг:
2.1.52. В составе серого вещества спинного мозга отсутствуют:
2.1.53. В белом веществе спинного мозга отсутствуют:
2.1.54. Паутинная оболочка мозга:
Содержит сеть тонких трабекул и цистерны с цереброспинальной жидкостью.
∙ Сосудистый слой оболочек мозга.
∙ Содержит слой эластических волокон.
∙ Непосредственно прилежит к ткани мозга и отграничена глальной мембраной.
2.1.55. Мягкая мозговая оболочка:
∙ Содержит сеть тонких трабекул и цистерны с цереброспинальной жидкостью.
Сосудистый слой оболочек мозга.
∙ Содержит слой эластических волокон.
∙ Непосредственно прилежит к ткани мозга и отграничена глальной мембраной.
2.1.56. Твердая мозговая оболочка:
∙ Содержит сеть тонких трабекул и цистерны с цереброспинальной жидкостью.
∙ Сосудистый слой оболочек мозга.
Образована плотной волокнистой соединительной тканью, содержащей слой эластических волокон.
∙ Непосредственно прилежит к ткани мозга и отграничена глальной мембраной.
2.1.57. Какую функцию выполняют псевдоуниполярные клетки спинномозгового узла?
∙ Они выполняют функции глиальных клеток.
Они являются чувствительными клетками соматической и вегетативной рефлекторной дуги.
∙ Они являются вставочными клетками любой рефлекторной дуги.
∙ Они обеспечивают двигательную иннервацию скелетных мышц.
2.1.58. Нейроны спинномозгового узла относятся к:
2.1.59. Дендриты псевдоуниполярных клеток находятся:
∙ В составе задних корешков спинного мозга. ∙ В составе передних корешков спинного мозга.
В составе смешанного нерва.
∙ В составе белого вещества спинного мозга.
2.1.60. Аксоны псевдоуниполярных клеток находятся:
В составе задних корешков спинного мозга.
∙ В составе передних корешков спинного мозга.
∙ В составе смешанного нерва.
∙ В составе центрального канала спинного мозга.
2.1.61. Тела псевдоуниполярных клеток находятся:
∙ В составе задних корешков спинного мозга.
∙ В составе передних корешков спинного мозга.
∙ В составе смешанного нерва.
На периферии спинномозгового ганглия.
2.1.62. Мантийными клетками в составе спинномозгового узла называют:
∙ Клетки, образующие оболочки нервных волокон узла.
Клетки нейроглии, окружающие тела псевдоуниполярных нейроцитов.
ЗНАЧЕНИЕ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ. ПОНЯТИЕ О НЕРВНЫХ ЦЕНТРАХ И ИХ СВОЙСТВА
Группа нейронов, необходимых для осуществления определенного рефлекса или для управления одной из функций организма, образует нервный центр . Центр ахиллова рефлекса (подошвенное сгибание стопы при ударе по ахиллову сухожилию) находится в I—II крестцовых сегментах спинного мозга. Дыхательный центр расположен в ретикулярной формации продолговатого мозга на дне IV желудочка и в спинном мозге.
Нервный центр — понятие физиологическое, а не анатомическое. В нервный центр могут входить нейроны (или группы нейронов), расположенные в разных отделах центральной нервной системы.
Нервные центры обладают рядом свойств, обусловленных особенностями механизма передачи возбуждения в синапсах.
Односторонность распространения возбуждения. В отличие от нервного волокна, в котором возбуждение распространяется в обе стороны от места раздражения, в нервном центре оно распространяется только в одном направлении — от центростремительного нейрона к центробежному. Это свойство обусловлено строением синапса: медиатор, осуществляющий передачу возбуждения, выделяется только в пресинаптическом окончании.
Замедление передачи возбуждения. В нервных центрах возбуждение замедляется. Это происходит потому, что в нервном центре может быть несколько нейронов и, следовательно, столько же сигналов. В каждом синапсе происходит синаптическая задержка возбуждения. В этой связи общая длительность задержки передачи возбуждения в нервном центре с одного синапса на другой зависит от количества вставочных нейронов: чем большее количество нейронов образует рефлекторную дугу, тем сильнее замедляется передача возбуждения в нервном центре этого рефлекса.
Суммация. Явление суммации в нервном центре впервые описал И. М. Сеченов (1863 г.). Это явление проявляется в накоплении (сложении) эффектов подпороговых раздражений. Одно подпороговое раздражение не вызывает ответной рефлекторной реакции: пресинап- тическое нервное окончание выделяет недостаточное количество медиатора. Несколько подпороговых раздражений в сумме дают нужный эффект: выделяется достаточное количество медиатора и возникает ответная рефлекторная реакция.
Различают временную и пространственную суммацию возбуждения в нервном центре. Временная суммация происходит под действием серии подпороговых раздражений, следующих друг за другом достаточно часто. Механизм временной суммации состоит в том, что каждое подпороговое раздражение повышает возбудимость нервного центра до тех пор, пока очередное из них не вызовет ответной рефлекторной реакции. Например, чихательный рефлекс возникает при длительном воздействии раздражителя на рецепторы слизистой оболочки полости носа.
Пространственная суммация возбуждения происходит при одновременном раздражении различных чувствительных нервов, передающих возбуждение в один и тот же нервный центр. Примером пространственной суммации возбуждения является рефлекторное сокращение полусухожильной мышцы при одновременном подпороговом раздражении мало- и большеберцового нервов. Подпороговое раздражение только одного из них сокращения не вызывает.
Пространственная суммация происходит вследствие схождения многих афферентных путей к одному нейрону (вставочному или эфферентному). Это явление называется конвергенцией.
Трансформация ритма возбуждения. Нервные центры способны трансформировать частоту и ритм поступающих импульсов. На одиночное раздражение, поступившее в нервный центр, последний может ответить серией импульсов. Если импульс поступает в нервный центр с частотой, превышающей лабильность этого центра, то последний ответит с частотой, соответствующей его возможностям, т. е. более редкими импульсами.
Последействие. Ответная рефлекторная реакция продолжается некоторое время и после прекращения действия раздражителя. Это явление называется рефлекторным последействием. Длительность рефлекторного последействия может во много раз превышать длительность действия самого раздражителя. Существует прямая зависимость: чем сильнее и дольше раздражение действует на рецептор, тем продолжительнее последействие. Причинами возникновения последействия являются следовая деполяризация и циркуляция нервных импульсов — наличие кольцевой связи между нейронами данного центра.
Утомляемость нервного центра. Нервное волокно практически неутомляемо. Утомление возникает в нервном центре центральной нервной системы, что обусловлено его низкой лабильностью. Такое утомление проявляется в постепенном уменьшении, а затем и прекращении рефлекторного ответа в случае продолжительного действия раздражителя. Утомление возникает в результате нарушения передачи возбуждений в синапсах.
Механизм фоновой активности предусматривает наличие кольцевой связи между нейронами, что обеспечивает передачу нервных импульсов с нейрона на нейрон. Фоновая активность нейронов повышает чувствительность центральной нервной системы к раздражениям, расширяет ее функциональные возможности, обеспечивает гибкость и пластичность. Изменение возбудимости центральной нервной системы. Центральная нервная система чрезвычайно чувствительна к различным воздействиям. При этом изменяется ее возбудимость. Она снижается при недостатке кислорода, при недостаточном кровообращении, в шоковом состоянии.
И. М. Сеченову принадлежит исключительная заслуга перед мировой наукой: он обнаружил в головном мозге центры, угнетающие спинномозговые рефлексы, и показал значение этих центров в рефлекторной координации двигательных актов.
Классический опыт И. М. Сеченова состоял в следующем. У лягушки перерезали головной мозг на уровне зрительных бугров. Переднюю часть мозга удаляли. После этого определяли время сгибательного рефлекса при раздражении лапки серной кислотой. Затем на зрительные бугры клали кристаллы поваренной соли и снова определяли продолжительность действия сгибательного рефлекса. Продолжительность рефлекса значительно увеличивалась, и через некоторое время реакция полностью исчезала.
После удаления раздражителя (кристалла соли) и промывания раздражаемого участка мозга физиологическим раствором реакция вновь возникала и продолжительность рефлекса восстанавливалась. Из этого опыта следует вывод: торможение — это активный процесс, возникающий, как и возбуждение, при раздражении любых участков центральной нервной системы. Значение открытия И. М. Сеченова состоит в том, что он установил одновременное существование в центральной нервной системе процессов возбуждения и торможения.
Торможение — это особый нервный процесс, внешне проявляющийся в уменьшении или полном исчезновении ответной реакции. Он представляет собой особую форму стойкого, неколеблющегося возбуждения, которое возникает вследствие сильного или длительного воздействия какого-либо раздражителя.
Различают торможение первичное и вторичное. Первичное торможение возникает с участием тормозных нейронов. Примером тормозных нейронов являются так называемые клетки Реншоу. Вторичное торможение возникает без участия тормозных нейронов. Оно является следствием сильного возбуждения нервной клетки. Возбуждение особенно легко сменяется торможением в участках нервной системы, обладающих низкой лабильностью.
Жизнь организма — согласованная работа всех его частей и приспособление к условиям среды — возможна благодаря центральной нервной системе. Она координирует все функции организма. Это обусловлено особенностями ее строения и функциональными свойствами. Существуют определенные закономерности координации нервных процессов.
Принцип общего конечного пути. Его открыл выдающийся английский физиолог Чарлз Скотт Шеррингтон. Суть этого принципа заключается в том, что к одному мотонейрону поступают импульсы от многих рецепторов, расположенных в различных частях тела. Этот процесс называется конвергенцией. Он обусловлен неодинаковым количеством афферентных и эфферентных нервных путей: первых примерно в пять раз больше, чем вторых. Из всех поступающих по различным путям в нейрон импульсов только некоторые, наиболее значимые в данный момент для организма, вызывают ответную реакцию. Конвергенция является одним из основных механизмов координации рефлек-торной деятельности.
Иррадиация возбуждения. Возбуждение, возникшее в одном из нервных центров под влиянием сильного и длительного раздражения, способно распространяться по центральной нервной системе, возбуждая новые участки. Распространение возбуждения называется иррадиацией (от лат. irradiare — сиять). Иррадиация возбуждения обусловлена наличием многочисленных связей между отдельными нейронами центральной нервной системы. Различают иррадиацию возбуждения избирательную и генерализованную.
Одновременная индукция характеризуется тем, что одновременно в одном центре возникает возбуждение, а в сопряженном центре — торможение (или наоборот). Примером может служить подтягивание на перекладине: в центре мышц-сгибателей возникает возбуждение, а в центре мышц-разгибателей — торможение. Последовательная положительная индукция проявляется в смене торможения возбуждением, а последовательная отрицательная — в смене возбуждения торможением. Принцип обратной связи. Воздействие работающего органа на состояние управляющего им нервного центра называется обратной связью.
Различают положительные и отрицательные обратные связи. Если импульсы, возникающие в результате какой-либо рефлекторной реакции, поступая в управляющий ею нервный центр, усиливают ее, — это положительная обратная связь; если же они угнетают эту реакцию, — это отрицательная обратная связь. Благодаря наличию обратной связи между нервным центром и управляемым им рабочим органом обеспечивается строгая согласованность их совместной деятельности и достигается наибольший эффект.
Доминанта — яркий пример взаимодействия возбудительного и тормозного процессов в центральной нервной системе. Наличие доминантного очага возбуждения резко изменяет обычные координационные отношения между этими процессами. Поступающие волны возбуждения, даже адресованные другим центрам, усиливают только его и вызывают характерную для него реакцию. В остальных нервных центрах в этот момент наступает торможение. Например, если в момент, предшествующий акту дефекации, раздражать у животного двигательные нервы, то вместо обычной ответной реакции — сгибания передней конечности — ускорится и усилится акт дефекации.
Доминантный очаг возбуждения характеризуется пятью признаками, определяющими характер его деятельности:
1) повышенной возбудимостью;
2) стойкостью возбуждения;
3) повышенной способностью к суммированию возбуждения;
4) инерцией, т. е. способностью длительно сохранять возбуждение после окончания действия раздражителя;
5) способностью вызывать сопряженные торможения.
Значение принципа доминанты А. А. Ухтомского заключается в установлении зависимости деятельности нервных центров и их взаимоотношений от исходного состояния. Будучи господствующим очагом возбуждения, нервный центр осуществляет специфическую ответную реакцию, угнетая другие центры. При этом он привлекает к себе все волны возбуждения, поступающие в центральную нервную систему и адресованные другим нервным центрам. Принцип доминанты играет большую роль в координирующей деятельности центральной нервной системы, в образовании условных рефлексов и двигательных навыков.
Нервные центры характеризуются пластичностью: в определенных условиях они перестраиваются и приобретают новые, не свойственные им ранее функции. Это доказывают специальные опыты. У животного перерезали подъязычный и диафрагмальный нервы, после чего дыхательные движения диафрагмы прекращались. Затем к центральному концу подъязычного нерва пришивали периферический конец диафрагмального. После заживления дыхательные движения диафрагмы восстанавливались. Из этого следует, что центр подъязычного нерва начинал управлять дыхательными движениями диафрагмы, т. е. приобретал новое функциональное значение.
Пластичность нервных центров позволяет перестраивать в широком диапазоне координационные отношения в центральной нервной системе. Это способствует наиболее совершенному приспособлению организма к изменяющимся условиям внешней и внутренней сред.
Не нашли то, что искали? Воспользуйтесь поиском:
Читайте также:
