
Цнс в организации движений
Вариант 1.
Системы управления движением:
· Пирамидная - двигательные центры коры больших полушарий в передней центральной извилине - формирует потоки нервных импульсов, идущих из двигательных центров коры в сегментам спинного мозга и скелетным мышцам, содержащих точные волевые посылки.
· Экстрапирамидная - премоторная зона неокортекса, базальные ганглии, субталамические ядра среднего мозга - создает тоническую предуготовленность мышц, необходимую для реализации произвольных движений, обеспечивает модуляцию команд, идущих по пирамидному тракту. Инстинктивные, автоматизированные движения.
Функциональные уровни управления:
2. Надсегментарный - стволовые структуры ГМ - двигательные ядра продолговатого и среднего мозга, отделы ретикулярной формации и мозжечка.
3. Высший (корково-подкорковый) - образования переднего мозга - базальные ганглии, сенсорно-моторные отделы неокортекса.
4. Блок регуляторно-трофического движения - отделы ВНС, иннервирующие скелетную мускулатуру. Способны оказывать опосредованное воздействие через изменение кровообращения.
+1 уровень, о котором говорил Юров, а Гриша не записал.
2) Функции спинного мозга. Спинальные рефлексы, их клиническое значение.
Функции СМ: осуществление двигательных и вегетативных рефлексов, проведение сенсорных сигналов от рецепторов к ГМ, передача информации от ГМ для координации деятельности мышц и органов.
Сухожильные рефлексы - частный случай рефлексов растяжения, резкое сокращение мышцы после легкого удара по её сухожилию.
Механизм: в момент удара происходит растяжение мышцы и возбуждение проприорецепторов, находящихся в мышечных волокнах, окруженных капсулой из соединительной ткани - интрафузальные. Проприорецептор - ПД - периферическая и центральная ветви первичного сенсорного нейрона в спинном ганглии - задний корешок - афферентные глутаматергические волокна - мотонейроны = моносинаптическая рефлекторная дуга.
Рефлексы растяжения мышцы - сокращение мышцы в ответ на ее растяжение. При быстром растяжении - фазический рефлекс - быстрое ответное сокращение. При медленном - тонический - сохранение неизменной длины мышцы при постоянном растяжении, поддержание мышечного тонуса.
Механизм: интрафузальные волокна тоньше и короче остальных, в их средней части находятся окончания первичного афферентного нейрона, которые реагируют на растяжение. Возбуждение передается по моносинаптичексой рефлекторной дуге к мотонейронам и мышцы сокращается.
При растяжении мышцы активируются α-мотонейроны (для экстрафузальных волокон) и ɣ-мотонейроны (для интрафузальных). Оба типа возбуждаются разными коллатералями одного сенсорного нейрона, поэтому оба типа мышечных волокон сокращаются вместе. Другие ветви афферентного нейрона активируют тормозные интернейроны, которые подавляют активность мотонейронов для мышц-антагонистов. Т.о. в механизме рефлекса растяжения запрограммирована одновременная согласованная деятельность нескольких мышцы.
Рефлекторная регуляция напряжения мышц.
Механизм: осуществляются с помощью сухожильных рецепторов Гольджи, расположенных между коллагеновыми пучками сухожильных волокон в области их прикрепления к мышце. + Рецепторы в суставах, высокочувствительные кожные рецепторы и нисходящие двигательные пути. Нарастание напряжения в мышце - натяжение сухожилий - деформация рецепторов - возрастание частоты ПД - тормозные интернейроны - гомонимные мотонейроны.
Кожные рецепторы - ощупывание хрупких или мелких предметов. Когда нужно согласовать силу мышечных сокращение с характером тактильных ощущений.
Сгибательные и разгибательные рефлексы.
Сгибательный рефлекс - немедленное сгибание конечности в ответ на болевое раздражение. Выраженность рефлекса пропорциональна силе раздражителя, т. е. чем больше стимул, тем больше мышцы задействовано. Перекрестный разгибательный рефлекс - разгибание коллатеральной конечности сопровождающее сгибательный рефлекс.
Механизм: полисинаптическая рефлекторная дуга = сенсорные нейроны - интернейроны - возбуждение мотонейронов мышц-сгибателей и угнетение мотонейронов мышц-разгибателей.
Способность суставов двигаться в разных плоскостях требует согласованных сокращение одних групп мышц при одновременном расслаблением других. Механизм: взаимодействие возбуждающих и тормозных интернейронов с мотонейронами мышц-синергистов и мышц-антагонистов.
Ритмический чесательный рефлекс.
Вызывается слабым раздражением кожи и включает: обнаружение раздраженного участка и ритмичные чесательные движения. Ритмичные чесательные движения обеспечиваются с помощью интернейронов, которые поочередно тормозят мотонейроны то сгибателей, то разгибателей. Чередуемые движения регулирует двигательный центр спинного мозга, состоящий из 2 полуцентров: сгибательного и разгибательного, которые реципрокно тормозят друг друга. Продолжительность торможения в цикле зависит от свойств тормозных синапсов, в которых происходит динамическое повышение порога возбуждения постсинаптических нейронов.
Участие в локомоции.
В управлении координированной деятельностью мышц при ходьбе и беге участвуют интернейроны и мотонейроны разных сегментов спинного мозга. Спинномозговые центры локомоции контролируются двигательными ядрами ствола, моторной корой и мозжечком. Для осуществления контроля используется сенсорная информация. Активность спинальных локомоторных центров полностью подчинена супраспинальным отделам и самостоятельно не проявляется.
Спинальные вегетативные рефлексы.
В зависимости от локализации различают:
· Висцеро-висцеральные проявляются при раздражении рецепторов внутренних органов. Проявляется в изменении деятельности этого же или другого внутреннего органа.
· Висцеродермальные. Раздражение рецепторов внутренних органов. Проявляется в изменении кожной чувствительности, потоотделения и электрического сопротивления кожи. Зоны Захарьина-Геда.
· Дерматовисцеральные. Раздражение рецепторов кожных участков - изменение деятельности внутренных органов, реакции сосудов.
Полисинаптические рефлекторные дуги. Нисходящий контроль осуществляется вегетативными центрами ствола мозга и гипоталамусом.
3) Ноцицептивная система, её биологическая роль, теории боли, классификация - виды боли.
Ноцицептивная система - многоуровневая сенсорная система, ответственная за восприятие и оценку повреждающего воздействия и формирование боли. Боль является одним из главных компонентов защитной системы организма. Это важнейший сигнал о повреждении ткани и развитии патологического процесса, постоянно действующий регулятор гомеостатических реакций, включая их высшие поведенческие формы.
1.Теория интенсивности (А. Гольдшейдер - 1894). Болевое ощущение возникает вследствие интенсивного раздражения различных сенсорных рецепторов и проведения болевых импульсов в определённые образования мозга.
2.Специфическая теория М. Фрея (1894). Боль возникает в результате возбуждения специфических полимодальных ноцицепторов.
3.Теория спиналъного воротного контроля (Р. Мелзак и П. Уолл - 1965). Открытие ворот для прохождения болевых импульсов в выше расположенные структуры ЦНС происходит на уровне желатинозной субстанции задних рогов спинного мозга. Открытие ворот происходит с участием субстанции Р, брадикинина, простагландинов и других ФАВ. Закрытие - энкефалины, тормозящие реализацию болевой афферентации с помощью субстанции Р и других алгогенных ФАВ.
4.Современная теория боли. Существуют специфические болевые рецепторы, специфические афферентные пути и специфические структуры головного мозга, формирующие болевое ощущение и реакции организма на него. Боль возникает вследствие превалирования активности ноцицептивной (алгогенной) системы над активностью постоянно функционирующей в здоровом организме антиноцицептивной (антиалгогенной) системы.
· Aδ-миелинизированные волокна. Проводят сигналы со скоростью от 5 до 30 м/с, служат для пропускания сигналов быстрой боли. Этот тип боли чувствуется за одну десятую доли секунды с момента возникновения болевого стимула.
· С-немиелинизированные волокна. Участвуют в передачи медленной боль. Скорость передачи от 0,5 до 2 м/с, —ноющая, пульсирующая, жгучая боль.
· Острая боль.
· Хроническая боль.
· Кожная боль -при повреждении кожи или подкожных тканей.
· Соматическая боль - в связках, сухожилиях, суставах, костях, кровяных сосудах.
· Внутренняя (висцеральная) боль - от внутренних органов тела. Сложно определить локализацию из-за малого количества рецепиторов.Нудящая и продолжительная боль. Часто ощущение боли приписывается участку тела, которое никак не связано с участком самого повреждения. Например, сердечная ишемия.
· Фантомная боль- ощущение боли в утраченной конечности.
· Психогенная боль. Диагностируют в отсутствие органического заболевания или в том случае, когда последнее не может объяснить характер и выраженность болевого синдрома. Имеет хронический характер и возникает на фоне психических расстройств.
Если Павлов создал физиологию элементов действий, то известный советский физиолог, специалист в области биомеханики движения, Николай Александрович Бернштейн создал физиологию, распространяющуюся на высшие формы психической деятельности. Если Павлов говорил о проторении нервных связей при образовании условных рефлексов, то в теории Бернштейна речь уже идет о построении сложного действия или навыка.
"В построении движений участвуют все уровни головного мозга. В стройном и согласованном ансамбле каждый из них ведет свою партию".
Центральная нервная система устроена иерархично, каждый вышележащий уровень подчиняет себе нижележащий: кора головного мозга, подкорковые — экстрапирамидные структуры, продолговатый мозг, спинной мозг, уровень сегмента спинного мозга и уровень простой рефлекторной дуги.
Уровни организации движений[править | править вики-текст]
уровень слепого выполнения движений (чистая физиология) обеспечивает настройку тонуса мышц (то есть способность мышц к релаксации, отношения мышц-антагонистов).
- Уровень D — Теменно-премоторный уровень (уровень действий), обеспечивающий целенаправленные, смысловые движения или серии движений, которые представляют собой манипуляции с предметами и требуют сложившихся в опыте представлений о форме, размерах и предназначении предметов, а также топологической схематизации пространства;
уровень слияния физиологического и психологического, он обеспечивает выработку сложных автоматизмов движения. Это уровень действия, уровень формирования двигательного навыка. На этом уровне формируются профессиональные движения. Применительно к осанке этот уровень отвечает за осознанное группирование тела — т.е осмысленную коррекцию положения тела.
(смысловой уровень) — обеспечивает смысловую коррекцию всех нижележащих уровней. Здесь формируются такие сложные действия-навыки, как пилотирование самолета, работа космонавта и другие. На этом уровне осуществляется самоконтроль осанки. Воздействуя на свои мысли, мы можем менять напряжение мышц и, расслабляя мышцы, сознательно корректировать привычную осанку и выработанную годами манеру двигаться. На этом уровне реализуются методы телесно ориентированной психотерапии (психосоматической коррекции) — высший уровень контроля осанки.
сознательное действие, автоматизируясь, переходит в уровень бессознательного, рефлекторного, и наоборот, рефлекторное может приобрести самостоятельное значение, войти в сферу сознания.
Это важный закон построения движения, он используется при коррекции ходьбы и осанки.
Индивидуально двигательный стереотип формируется под влиянием разнообразных факторов, имеющих отношение к становлению моторики. Эти факторы могут быть закономерными и случайными: вспомним школьника, склонившегося над низким столом и проводящего многие часы учебы в этой позе, лишенного своевременной педагогической поправки. В последующем накопившиеся двигательные ошибки суммируются с неизменно наступающими возрастными изменениями в осанке подростка, что в итоге способствует фиксации патологической цепи ошибок в формировании осанки, пластики, ловкости. Подобных ситуаций достаточно в профессиональной деятельности, когда люди с правильно сформированной моторикой в нерациональной рабочей позе подвергают ДС проверке на устойчивость с нередкими потерями. Здесь ДС выступает как индивидуальное приобретение моторики.
В основе рефлекторной деятельности лежит восприятие раздражения и перенос его на реагирующий орган.
Рефлекторная дуга состоит чаще из цепи двух или трех нейронов, по которым передается нервный импульс. Та часть дуги, которая воспринимает раздражение и проводит его в пределы ЦНС, называется афферентной; та, которая отводит ответный импульс к рабочему органу, именуется эфферентной.
По месту приложения раздражения рефлексы могут быть разделены на:
* поверхностные (кожные, со слизистых оболочек): корнеальный, глоточный, рефлекс с мягкого неба, брюшные рефлексы (верхний, средний и нижний), кремастерный, подошвенный и анальный рефлексы;
* глубокие (сухожильные, периостальные): надбровный, нижнечелюстной, сгибательно-локтевой, разгибательно-локтевой, пястно-лучевой, коленный, ахиллов рефлексы.
Рефлексы могут изменяться в сторону:
* понижения или утраты их;
* в патологических условиях могут появляться новые рефлексы, в норме не вызываемые - патологические рефлексы. К числу патологических рефлексов относятся следующие рефлексы орального автоматизма:
* назолабиальный рефлекс (М.И.Аствацатуров);
* дистант-оральный рефлекс (С.И.Карчикян);
* ладонно-подбородочный рефлекс (Маринеско-Родовичи).
На руках исследуются верхний симптом Россолимо, симптом Якобсона - Ласка.
* разгибательные рефлексы - рефлекс Бабинского, Оппенгейма, Гордона, Шефера, Чеддока;
* сгибательные рефлексы - рефлекс Россолимо, Бехтерева -Менделя, Жуковского-Корнилова.
Спинной мозг обеспечивает протекание многих элементарных двигательных рефлексов, включение которых в сложные двигательные акты и регуляция по мощности, пространственной ориентации и моменту включения осуществляется вышележащими отделами головного мозга под контролем коры больших полушарий.
Роль спинного мозга и подкорковых отделов ЦНС в регуляции движений
Спинной мозг осуществляет ряд элементарных двигательных рефлексов: рефлексы на растяжение (миотатические и сухожильные рефлексы, например, коленный рефлекс), кожные сгибательные рефлексы (например, защитный рефлекс отдергивания конечности при уколах, ожогах), разгибательные рефлексы (рефлекс отталкивания от опоры, лежащий в основе стояния, ходьбы, бега), перекрестные рефлексы и др.
Элементарные двигательные рефлексы включаются в более сложные двигательные акты — регуляцию деятельности мышц-антагонистов, ритмических и шагательных рефлексов, лежащих в основе локомоций и других движений.
Для сгибательного движения в суставе необходимо не только сокращение мышц-сгибателей, по и одновременное расслабление мышц-разгибателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей — торможение. При разгибании сустава, наоборот, тормозятся центры сгибателей и возбуждаются центры разгибателей. Такие координационные взаимоотношения между спинальными моторными центрам и названы реципрокной (взаимосочетанной) иннервацией мышц-антагонистов. Однако реципторные отношения между центрами мы�?ц-антагонистов в необходимых ситуациях (например, при фиксации суставов, при точностных движениях) могут сменяться одновременным их возбуждением.
Нейроны промежуточной продольной зоны коры мозжечка согласуют позные реакции с движениями. Они выполняют также точные расчеты по ходу движений, необходимые для коррекции ошибок и адаптации моторных программ к текущей ситуации. Программирование каждого последующего шага осуществляется ими на основе анализа предыдущего. Кроме того производится согласование движений рук и ног, и особенно — регуляция активности мышц-разгибателей, обеспечивающих опорную фазу движения. Значение мозжечка в четком поддержании темпа ритмических движений объясняют геометрически правильным чередованием рядов эфферентных клеток Пуркинье и походящих к ним афферентных волокон.
К управлению ритмическими движениями непосредственное отношение имеют активирующие и угнетающие отделы ретикулярной формации, влияющие на силу и темп сокращения мышц, а также подкорковые ядра, которые организуют автоматическое их протекание и содружественные движения конечностей. Включение древних форм ритмических движений (циклоидных) в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи. То же самое происходит при освоении акта ходьбы — с переходом от отдельных шагов к ритмической походке. Плавность ритмических движений, четкое чередование реципрокных сокращений мышц обеспечивают премоторные отделы коры.
1.Роль спинного мозга и подкорковых отделов ЦНС в регуляции движений
Спинной мозг обеспечивает протекание многих элементарных дви-гательных рефлексов, включение которых в сложные двигательные акты и регуляция осуществляется вышележащими отделами головного мозга под контролем коры больших полушарий.
Спинной мозг осуществляет ряд элементарных двигательных реф-лексов: рефлексы на растяжение (миотатические и сухожильные рефлек-сы, например, коленный рефлекс), кожные сгибательные рефлексы (на-пример, защитный рефлекс отдергивания конечности при уколах, ожогах), разгибателъные рефлексы (рефлекс отталкивания от опоры, лежащий в основе стояния, ходьбы, бега), перекрестные рефлексы и др.
Элементарные двигательные рефлексы включаются в более сложные двигательные акты – регуляцию деятельности мышц-антаго-нистов, ритмических и шагательных рефлексов, лежащих в основе локо-моций и других движений.
Для сгибательного движения в суставе необходимо не только сокра-щение мышц-сгибателей, но и одновременное расслабление мышц-разги-бателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей – торможения. При разгибании сустава, наоборот, тормозятся центры сгибателей и возбужда-ются центры разгибателей. Такие координационные взаимоотношения ме-жду спинальными моторными центрами названы реципрокной (взаимосо-четанной) иннервацией мышц-антагонистов. Однако реципрокные отно-шения между центрами мышц-антагонистов в необходимых ситуациях (например, при фиксации суставов, при точностных движениях) могут сменяться одновременным их возбуждением.
Нейроны промежуточной продольной зоны коры мозжечка согла-суют позные реакции с движениями. Они выполняют также точные расчеты по ходу движений, необходимые для коррекции ошибок и адаптации мо-торных программ к текущей ситуации. Программирование каждого после-дующего шага осуществляется ими на основе анализа предыдущего. Кроме того, производится согласование движений рук и ног и, особенно, регуляция активности мышц-разгибателей, обеспечивающих опорную фазу движения. Значение мозжечка в четком поддержании темпа ритмических движений объясняют геометрически правильным чередованием рядов эфферентных клеток Пуркинье и подходящих к ним афферентных волокон.
К управлению ритмическими движениями непосредственное отно-шение имеют активирующие и угнетающие отделы ретикулярной фор-мации,влияющие на силу и темп сокращения мышц, а также подкорковые ядра, которые организуют автоматическое их протекание и содружествен-ные движения конечностей. Включение древних форм ритмических дви-жений в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи. То же самое происходит при ос-воении акта ходьбы – с переходом от отдельных шагов к ритмической по-ходке. Плавность ритмических движений, четкое чередование реципрок-ных сокращений мышц обеспечивают премоторные зоны коры.
2.Роль различных отделов коры больших полушарий в регуляции движений
В организации двигательных актов участвуют практически все отделы коры больших полушарий.
Моторная область коры (прецентральная извилина) посылает им-пульсы к отдельным мышцам, преимущественно к дистальным мышцам
Постцентральная извилина коры представляет собой общечувстви-тельное поле, которое обеспечивает субъективное ощущение движений. Нижнетеменные области коры (задние третичные поля) формируют пред-ставления о взаимном расположении различных частей тела и положении тела в пространстве, обеспечивают точную адресацию моторных команд к отдельным мышцам и пространственную ориентацию движений. Лимбиче-ская система (нижние и внутренние части коры) ответственна за эмоцио-нальную окраску движений и управление вегетативными их компонентами.
В высшей регуляции произвольных движений важнейшая роль принадлежит передним третичным полям.Здесь помимо обычных вер-тикальных колонок нейронов существует принципиально новый тип функ-циональной единицы – в форме замкнутого нейронного кольца. Циркуля-ция импульсов в этой замкнутой системе обеспечивает кратковременную память. Она сохраняет в коре возбуждение между временем прихода сен-сорных сигналов и формированием ответной эфферентной команды. Такой механизм служит основой сенсомоторной интеграции при программирова-нии движений, при осуществлении зрительно-двигательных реакций.
Функцией передней третичной области коры является сознатель-ная оценка текущей ситуации и предвидение возможного будущего, выра-ботка цели и задачи поведения, программирование произвольных дви-жений, их контроль и коррекция. Соответствие выполняемых действий поставленным задачам придает движениям человека определенную целе-сообразность и осмысленность. При поражении лобных долей движения человека становятся бессмысленными.
3.Речевая регуляция движений
Спецификой регуляции движений у человека является то,что они подчинены речевым воздействиям,т.е. могут программироваться лобными долями в ответ на поступающие из внешней среды словесные сигналы, а также благодаря участию внешней или внутренней речи (мыш-ления) самого человека. В этой функции принимают участие расположен-ные в левом полушарии человека сенсорный центр речи Вернике и мо-торный центр речи – центр Брока. Считают, что афферентная импульса-ция от речевой мускулатуры является важным ориентиром, дополняющим проприоцептивные сигналы от работающих мышц, а формирующиеся на
Речевой основе избирательные связи в коре облегчают составление моторных программ.
Эта управляющая система еще не развита у ребенка 2 - 3 лет. Она появляется лишь к 3 - 4 годам. Внешняя речь, сменяясь постепенно шепотом и переходя затем во внутреннюю речь, становится важным регулятором моторных действий взрослого человека.
Нисходящие моторные системы
Высшие отделы головного мозга осуществляют свои влиянияна деятельность нижележащих отделов, в т.ч. спинного мозга, через нисходящие пути, которые группируют обычно в две основные нисходящие системы - пирамидную и экстрапирамидную.
Пирамидная система выполняет 3 основные функции:
■ посылает мотонейронам спинного мозга импульсы - команды к движениям (пусковые влияния);
■ изменяет проведение нервных импульсов во вставочных спи-нальных нейронах, облегчая протекание нужных в данный момент спинномозговых рефлексов;
■ осуществляет контроль за потоками афферентных сигналовв нервные центры, выключая постороннюю информацию и обеспечивая обратные связи от работающих мышц.
Волокна пирамидной системы вызывают преимущественно возбуждение мотонейронов мышц-сгибателей, особенно влияя на отдельные мышцы и даже части мышц верхних конечностей, в частности, на мышцы пальцев рук.
обеспечивают повышение возбудимости мотонейронов мышц-сгибателей (корково-спинномозговая, корково-красноядерно-спинномозговая и корко-во-ретикуло-спинномозговая), а одна система (вестибуло-спинномозговая) – тормозит эти мотонейроны.
Материалы для самостоятельной подготовки
Вопросы к коллоквиуму и для самоконтроля
1. Назовите основные принципы организации движений.
2. Что называют функциональной системой?
3. Какие процессы включает функциональная система?
4. Назовите физиологические механизмы, лежащие в основе управления про-извольными движениями человека.
5. Опишите механизм рефлекторного кольцевого регулирования; программно-го управления движениями.
6. Перечислите три основных функциональных блока мозга и выполняемые ими функции.
7. Какова роль различных отделов ЦНС в регуляции позно-тонических реакций?
8. Какие рефлексы относятся к установочным? Какие отделы головного мозга участвуют в их осуществлении?
9. В чем заключается роль различных отделов ЦНС в регуляции движений?
10. Какое значение имеет речь в регуляции движений?
11. Назовите основные нисходящие моторные системы.
12. Перечислите функции пирамидной, экстрапирамидной системы.
Модуль 7 ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ
Лекция 19ОБЩАЯ ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ
1.Основные принципы строения и общий план организации сенсорных систем
Организм и окружающая его среда представляют собой единство (И. М. Сеченов). Человеку, для которого характерны сложные акты поведения во внешней среде, необходимы постоянное получение информации о состоянии и изменениях внешней среды и внутренних органов, переработка этой информации и на основе ее составление программ предстоящей деятельности. Постоянный анализ окружающего мира обеспечивается работой ряда механизмов и систем,которые называют сенсорными,или анализаторами.Учение об анализаторах было создано И. П. Павловым. Анализатором Павлов считал совокупность нейронов, участвующих в восприятии раздражений, проведении возбуждения, а также анализе его свойств клетками коры большого мозга. Работа любого анализатора начинается с восприятия рецепторами внешней для мозга физической или химической энергии, трансформации ее в нервные сигналы и передача их в мозг через цепи нейронов, образующих ряд уровней. Процесс передачи сенсорных сигналов сопровождается многократными их преобразованиями и завершается высшим анализом и синтезом (опознанием образа), после чего происходит выбор или разработка программы ответной реакции организма. Современное представление об анализаторах как сложных многоуровневых системах, передающих информацию от рецепторов к коре и включающих регулирующие влияния коры на рецепторы и нижележащие центры, привело к появлению более общего понятия сенсорные системы.
Всем сенсорным системам свойственны следующие основные принципы строения:
1. многослойность,т.е. наличие нескольких слоев нервных клеток, первый из которых связан с рецептором, а последний - с нейронами ассоциативных отделов коры полушарий;
2. многоканальность- наличие в каждом из слоев множества (иногда до миллиона) нервных элементов, связанных со множеством элементов следующего слоя и т.д. Наличие множества каналов обеспечивает сенсорным системам животных и человека большую надежность и тонкость анализа;
3. неодинаковое число элементов в соседних слоях,например, в зрительной сенсорной системе;
4. дифференциация сенсорных системзаключается в образовании отделов, состоящих из того или иного числа слоев нервных элементов.
Общий план организации сенсорных систем.В составе сенсорной системы различают 3 отдела:
■ периферический, или рецепторный, состоящий из рецепторов, воспринимающих определенные сигналы, и специальных образований, представляющих собой органы чувств - глаз, ухо и др.;
■ проводниковый,включающий проводящие пути и подкорковые нервные центры;
■ корковый- области коры больших полушарий, которым адресуется данная информация.
Нервный путь, связывающий рецептор с корковыми клетками, обычно состоит из четырех нейронов: первый,чувствительный, нейронрасположен вне ЦНС - в спинномозговых узлах или узлах черепно-мозговых нервов (спиральном узле улитки, вестибулярном узле и др.); второй нейроннаходится в спинном, продолговатом или среднем мозге; третий нейрон -в релейных (переключательных) ядрах таламуса; четвертый нейронпредставляет собой корковую клетку проекционной зоны коры больших полушарий.
Последнее изменение этой страницы: 2016-08-06; Нарушение авторского права страницы
Мы рассмотрели, как мышцы, развивая усилие, воздействуют на кости, к которым они прикреплены, производя, таким образом, движение. Этот процесс невозможен без участия нервной системы. Как скелет остается неподвижным до тех пор, пока мышцы не приложат усилие, так и мышцы не могут сокращаться до тех пор, пока их не возбуждает нервная система. Нервная система планирует, начинает и координирует все движения человека. Эта роль нервной системы — контроль движений человека
Нервная система влияет на всю физиологическую деятельность организма человека. Нервы образуют своеобразную сеть, по которой электрические импульсы передаются практически во все участки тела, а также принимаются из них. Головной мозг действует как компьютер, интегрируя поступающую информацию, подыскивая нужный ответ и затем инструктируя соответствующие части тела, как поступить. Таким образом, нервная система обеспечивает коммуникацию и координацию взаимодействий между всеми тканями организма, а также с внешним миром.
Основная единица нервной системы — нейрон. Обычный нейрон состоит из тела, или сомы, дендритов и аксона. Тело содержит ядро. От тела отходят отростки — дендриты и аксон. Сбоку по направлению к аксону клетка сужается, образуя аксонный холмик, который играет важную роль в передаче импульсов.
Нейроны содержат множество дендритов. Это — рецепторы нейронов. Большинство импульсов, поступающих в нерв из соседних нейронов, как правило, поступают в нейрон через лендриты и затем передаются телу нейрона. Наряду с этим нейроны, как правило, имеют лишь один аксон. Это—нейромедиатор нейрона, передающий импульсы из тела клетки. Аксон разветвляется, образуя окончания, или терминальные фибриллы. Окончания аксона расширяются, образуя крошечные выпуклости, так называемые синаптические холмики. В них содержится множество пузырьков (мешочков), наполненных химическими соединениями — нейромедиаторами, обеспечивающими связь нейронов друг с другом (речь об этом пойдет далее).
Дендриты - ветвящиеся короткие отростки, воспринимающие сигналы от других нейронов, рецепторных клеток или непосредственно от внешних раздражителей. Дендрит проводит нервные импульсы к телу нейрона.
Аксоны – длинный отросток, для проведения возбуждения от тела нейрона.
Уникальными способностями нейрона являются:
- способность генерировать электрические заряды
- передавать информацию с помощью специализированных окончаний – синапсов.
По функциям нейроны подразделяются на:
- эфферентные (рецепторные, чувствительные),передающие информацию от органов чувств в центральные отделы нервной системы;
- эфферентные (двигательные, моторные; лат. efferens - выносящий), посылающие импульсы к различным органам и тканям. Они находятся главным образом в передних рогах спинного мозга и в специализированных центрах головного мозга;
- вставочные (замыкательные, кондуктивные, промежуточные), служащие для переработки и переключения импульсов. ЦНС на 90% состоит из вставочных нейронов.
Тела эфферентных нейронов находятся в ЦНС (или в симпатических и парасимпатических узлах). Их аксоны идут к рабочим органам (мышцам или железам). Различают два вида рабочих, или исполнительных, органов: анимальные - поперечнополосатые (скелетные) мышцы и вегетативные - гладкие мышцы и железы. Соответственно этому имеются нервные окончания аксонов эфферентных нейронов двух типов: двигательные и секреторные. Первые ( мотонейроны ) оканчиваются на мышечных волокнах, образуя бляшки, которые в поперечнополосатых мышцах представляют аксомышечные синапсы. Нервные окончания неисчерченной (гладкой) мышечной ткани образуют вздутия, в которых также содержатся синаптические пузырьки. Секреторные окончания контактируют с железистыми клетками.
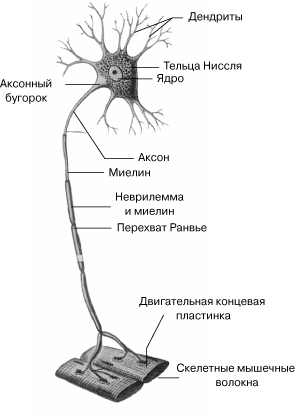
Рис. 26. Мотонейрон (ДВИГАТЕЛЬНЫЙ НЕЙРОН)
Управление сократительной активностью мышцы осуществляется с помощью большого числа мотонейронов– нервных клеток, тела которых лежат в спинном мозге, а длинные ответвления – аксоны в составе двигательного нерва подходят к мышце (рис.26). Войдя в мышцу, аксон разветвляется на множество веточек, каждая из которых подведена к отдельному волокну, подобно электрическим проводам присоединенным к домам.. Мотонейроны (нейроны эфферентные, двигательные, моторные)
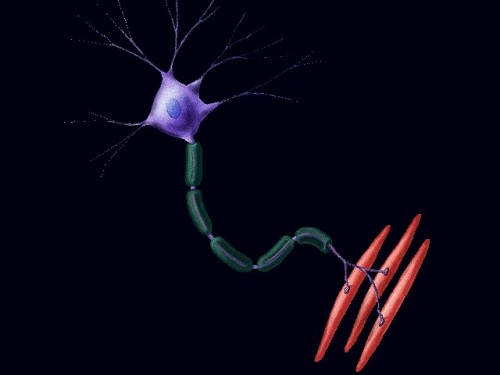
Рис. 27. Двигательная единица
Мотонейрон и мышечные волокна, которые он иннервирует, составляют двигательную единицу (рис. 27) Таким образом, один мотонейрон управляет целой группой волокон, которая работает как единое целое.
Двигательная единица (моторная единица):
группа мышечных волокон, иннервируемых одним двигательным нейроном,
сочетание моторного нейрона и возбуждаемых им волокон мышцы.
Когда нейрон посылает сигнал, все мышечные волокна в такой единице отвечают одновременно. В крупных мышцах в одной единице может быть до 2 тыс. волокон, тогда как в очень маленьких мышцах их всего 10—20. Сила мышечного сокращения зависит от того, сколько моторных единиц сокращается в ответ на импульсы от головного мозга, какие типы клеток вовлечены в сокращение и какое напряжение развивает каждая клетка. Изменяя число моторных единиц, вовлекаемых в работу данной мышцы, наше тело осуществляет тонкую настройку мышечной силы, требующейся ему в разных ситуациях постоянно меняющейся повседневной жизни.
При подходе аксона к поверхности мышечного волокна миелиновая оболочка заканчивается, и он образует терминальную часть (нервное окончание) в виде нескольких коротких отростков, располагающихся в желобках на поверхности мышечного волокна (аксон мотонейрона разделяется на множество ветвей, каждая из которых образует одно соединение с мышечным волокном) . Таким образом, один мотонейрон иннервирует много мышечных волокон, но каждым мышечным волокном управляет ветвь только от одного мотонейрона. Мышечные волокна одной двигательной единицы находятся в одной и той же мышце, но не в виде компактной группы, а рассеяны по ней . Когда в мотонейроне возникает потенциал действия, все они получают стимул к сокращению.
Мышцы, контролирующие выполнение тонких движений (например, движения глаз), имеют не-большое количество мышечных волокон на один двигательный нейрон. В мышцах, имеющих более общие функции, содержится много волокон на двигательный нейрон. Мышечные волокна определенной двигательной единицы гомогенны относительно типа волокна. Следовательно, вы не найдете двигательную единицу, содержащую как быстро-, так и медленносокращающиеся волокна. считается, что характеристики двигательного нейрона определяют тип волокна в данной двигательной единице.
Что такое двигательная единица?
Двигательная единица состоит из а-мотонейрона и всех иннервируемых им мышечных волокон. Все волокна одной двигательной единицы относятся к одному типу (БОГВ, МОВ или БГВ).
Дайте определение понятия "коэффициент иннервации".
Коэффициент иннервации показывает, сколько мышечных волокон иннервируется одним а-мотонейроном. Низкий коэффициент означает, что один нейрон иннервирует лишь несколько волокон. Такие мотонейроны контролируют тонкую моторику, поскольку активация двигательных единиц в различных сочетаниях обеспечивает плавно изменяющиеся сокращения. Более высокий коэффициент иннервации означает, что один нейрон иннервирует большое количество мышечных волокон. В соответствии с законом "все или ничего" надпороговая стимуляция такого нейрона вызывает сокращение всех иннервируемых им волокон. Высокий коэффициент иннервации поддерживает сильные сокращения ценой потери точности. Стимулируя различные — по виду и по количеству — двигательные единицы, ЦНС может вызывать разнообразные мышечные движения.
Мышцы, контролирующие движения глаз (экстраокулярные мышцы) имеют коэффициент иннервации 1:15, что означает, что один нейрон обслуживает всего 15 мышечных волокон. Наоборот, икроножная и передняя бояъшеберцовая мышцы имеют коэффициент иннервации порядка 1:2 000
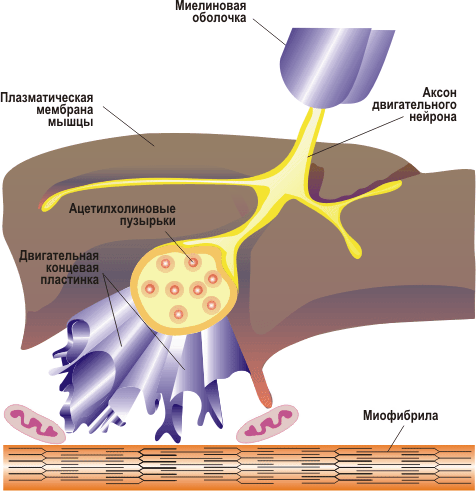
Рис. 28. Нервно-мышечное соединение, нервно-мышечный синапс
Если связь между нейронами осуществляется в помощью синапсов, то их связь с мышечными волокнами происходит в нервно-мышечном соединении. Нервно-мышечное соединение выполняет ту же функцию, что и синапс, включает пресинаптическое окончание аксона, синаптическую щель и рецепторы на сарколемме мышечного волокна. В нервно-мышечном соединении импульс принимает мышечное волокно.
При подходе аксона к поверхности мышечного волокна миелиновая оболочка заканчивается, и он образует терминальную часть (нервное окончание) в виде нескольких коротких отростков, располагающихся в желобках на поверхности мышечного волокна. Область плазматической мембраны мышечного волокна, лежащая непосредственно под нервным окончанием, обладает особыми свойствами и называется двигательной концевой пластинкой. Структура, состоящая из нервного окончания и двигательной концевой пластинки, - это нервно-мышечное соединение (нервно-мышечный синапс) (рис. 28). Синаптическая мембрана аксона и постсинаптическая мембрана мышечного волокна разделены синаптической щелью. В этой области мышечное волокно не имеет поперечной исчерченности, характерно скопление митохондрий и ядер. Терминали аксонов содержат большое количество митохондрий и синаптических пузырьков с медиатором ацетилхолином.
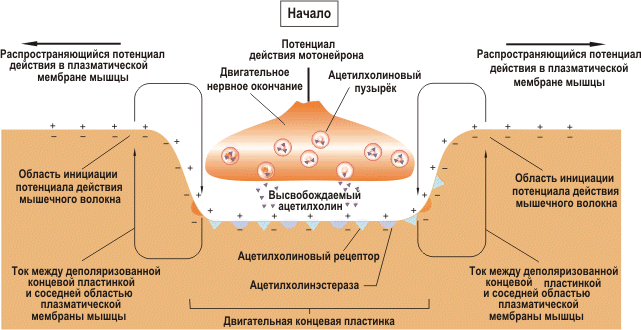
Рис.29. Генерирование потенциала действия в плазматической мембране мышечного волокна.
События в нервно-мышечном соединении, приводящие к генерированию потенциала действия в плазматической мембране мышечного волокна представлены на рис.29.
Когда моторный нейрон активизирован, он выпускает медиатор ацетилхолин в нейромышечном соединении. Ацетилхолин распространяется по нему и достигает рецепторов на мышечной мембране. Это заставляет натрий быстро проникать в мышечную мембрану, а калий – немедленно покидать ее - процесс деполяризации мышечной мембраны. Когда эта мембрана деполяризована, открываются каналы кальция в мышце. Так кальций проникает во внутриклеточное пространство мышцы. Кальций связывается с белком тропонином на актиновой нити (представьте себе тропонин контрольно-пропускным пунктом между миозиновыми и актиновыми нитями. Тропонин блокирует движение миозина и актина, когда находится в покое). И тогда с помощью АТФ миозиновые и актиновые нити могут скользить мимо друг друга. Это вызывает сокращение мышцы – удивительно сложный процесс, начинающийся с сигнала моторного нейрона. Важно, что большинство нервно- мышечных соединений расположены в срединной части мышечного волокна, откуда возникший потенциал действия распространяется к обоим его концам. Таким образом, каждый потенциал действия мотонейрона, как правило, вызывает потенциал действия в каждом мышечном волокне своей двигательной единицы.
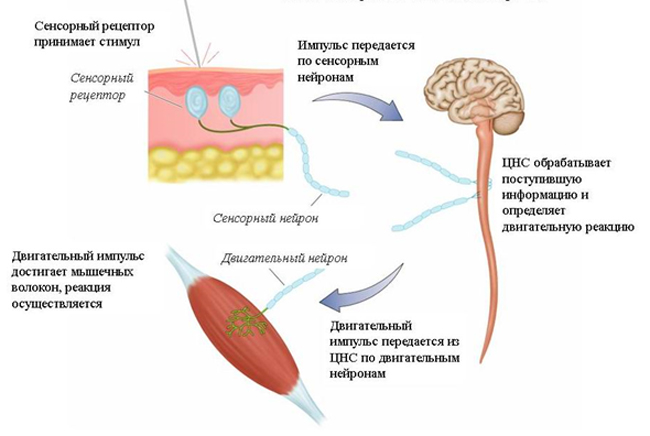
Рис. 30. Сенсорно-двигательная интеграция
Как происходит Управление движениями?
Сокращая мышцы, мы изменяем, положение частей тела, т.е. производим движение. В двигательной деятельности человека различают непроизвольные движения, происходящие без участия сознания и представляющие собой безусловные рефлексы, либо автоматизированные двигательные навыки, а также произвольные движения – сознательно управляемые целенаправленные действия. Произвольные движение человека сознательно управляются (контролируются) нервной системой посредством механизма сенсорно-двигательной интеграции. Любое движение можно рассматривать как двигательную реакцию на определенный внешний или внутренний стимул.
Сенсорно-двигательная интеграция –
механизм, посредством которого, нервная система управляет (контролируют) произвольные движения человека
взаимодействие сенсорного и двигательного отделов нервной системы, в результате чего осуществляется выполнение двигательных реакций
процесс последовательного функционирования сенсорного и двигательного отделов нервной системы, чтобы тело прореагировало на сенсорный стимул
процесс последовательного взаимодействия сенсорного и двигательного отделов нервной системы, в результате чего осуществляется управление (контроль) движениями человека
Сенсорно-двигательная интеграция представляет собой процесс, посредством которого ПНС передает сенсорный импульс ЦНС, где он интерпретируется, после чего посылается в соответствующий двигательный сигнал, направленный на то, чтобы вызвать необходимую двигательную реакцию.
Процесс, который называется сенсорно-двигательной интеграцией, показан на рис. 30. Чтобы тело прореагировало на сенсорный стимул, сенсорный и двигательный отделы нервной системы должны функционировать вместе в такой последовательности событий:
1) сенсорные рецепторы принимают сенсорный стимул ;
2). сенсорный импульс передается по сенсорным нейронам в ЦНС ;
3) ЦНС обрабатывает поступившую сенсорную информацию и определяет наиболее подходящую реакцию на нее;
4) сигналы реакции передаются из ЦНС по двигательным нейронам ;
5) двигательный импульс передается мышце и реакция осуществляется.
Сенсорный импульс
Ощущения и физиологический статус организма определяют сенсорные рецепторы. Импульсы вследствие сенсорного стимулирования передаются через сенсорные нервы в спинной мозг. Достигнув его, они "включают" локальный рефлекс на данном уровне или идут в более высокие участки спинного мозга или в головной мозг. Сенсорные пути к головному мозгу могут прерываться в сенсорных участках ствола мозга, в мозжечке, таламусе или коре головного мозга. На рис. 31 показаны сенсорные рецепторы, а также их нервные пути к спинному мозгу и различным участкам головного мозга.
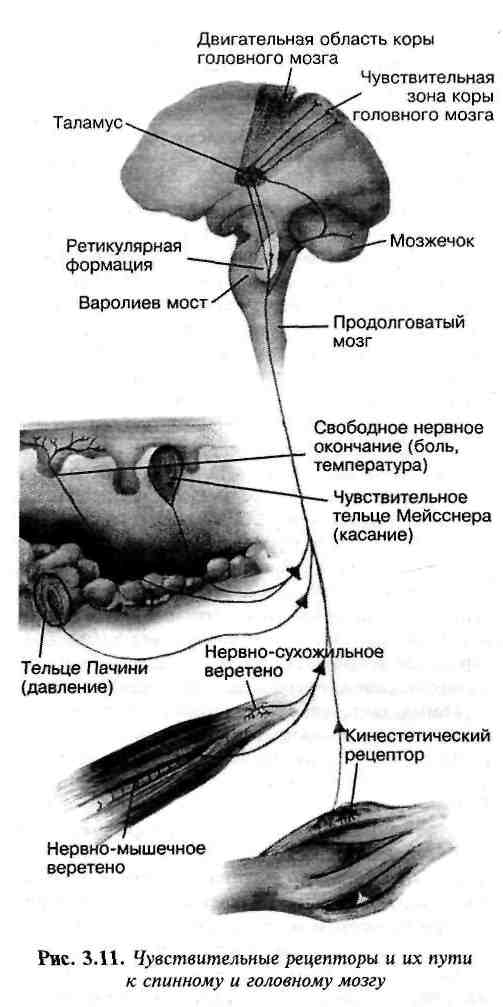
Рис. 31. Чувствительные рецепторы и пути к спинному и головному мозгу
Функции интеграционных центров различны:
· сенсорные импульсы, которые прерываются (заканчиваются) в спинном мозгу, здесь и интегрируются. Реакцией обычно является простой двигательный рефлекс, представляющий собой наипростейший тип интеграции;
· сенсорные импульсы, заканчивающиеся в нижней части ствола мозга, вызывают подсознательные двигательные реакции более высокого уровня и более сложные, чем рефлексы спинного мозга, контроль при передвижении, пребывании в положении сидя или стоя;
· сенсорные импульсы, которые заканчиваются в мозжечке, также участвуют в реализации подсознательного контроля движения. Это центр координации, делающий наши движения более плавными, координируя действия различных сокращающихся мышечных групп. Мозжечок вместе с базальными ядрами головного мозга координирует все тонкие и грубые движения тела. Без контроля со стороны мозжечка все выполняемые движения были бы нескоординированными и неконтролируемыми;
· сенсорные сигналы, заканчивающиеся в таламусе, достигают уровня сознания, и человек начинает различать все возможные ощущения;
· когда сенсорные сигналы попадают в кору головного мозга, человек может дискретно локализовать сигнал. Основная чувствительная зона коры головного мозга, расположенная в постцентральной извилине (в теменной доле), принимает общие сенсорные импульсы от рецепторов кожи, а также проприорецепторов мышц, сухожилий и суставов. В этом участке имеется “карта” всего тела. Стимулирование в определенном участке распознается, и его точное нахождение сразу же становится известным. Таким образом, эта часть головного мозга постоянно обеспечивает нас информацией обо всем, что нас окружает, и о нашей взаимосвязи с окружающей средой.
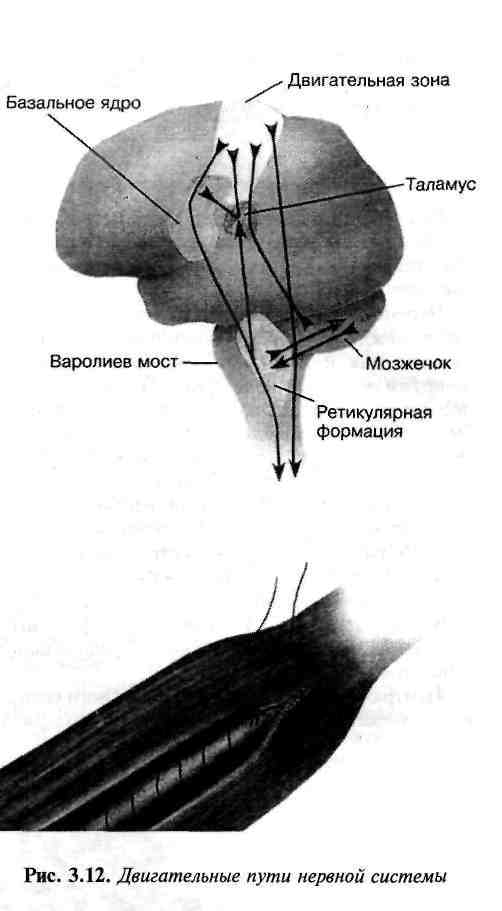
Рис. 32. Двигательные пути нервной системы
После поступления в ЦНС сенсорного импульса сразу же возникает реакция двигательного нейрона, независимо от уровня, на котором "остановился" импульс. Контроль скелетных мышц осуществляют импульсы, проводимые двигательными (эфферентными) нейронами, которые берут начало в одном из трех уровней: 1) спинном мозгу; 2) нижних участках головного мозга; 3) двигательном участке коры головного мозга.
По мере перемещения уровня осуществления контроля от спинного мозга к двигательной области коры головного мозга увеличивается сложность движений от простых рефлексов к усложненным движениям, выполнение которых требует участия мыслительных процессов. Двигательные реакции более сложных движений, как правило, берут свое начало в двигательной зоне коры головного мозга. Некоторые двигательные пути показаны на рис. 32.
Теперь мы можем связать воедино две системы на основании сенсомоторной интеграции. Простейшей ее формой является рефлекс, с него мы и начнем.
Рефлекторная деятельность. Рефлексы представляют собой простейшую форму нервной интеграции. Рефлекс представляет собой заранее запрограммированную реакцию. В любой момент при передаче сенсорными нервами особых импульсов ваше тело реагирует мгновенно и одинаково. К тому времени, когда вы осознанно ощутите особый стимул, после того как сенсорные импульсы будут переданы в чувствительную зону коры головного мозга, рефлекторная деятельность, скорее всего, завершится. Вся нервная деятельность протекает очень быстро, однако рефлекс — наиболее быстрый режим реакции, поскольку не требует принятия решения. Возможна лишь одна реакция — нет необходимости рассматривать варианты. Рассмотрим два рефлекса, которые помогают контролировать мышечную функцию:
Рассмотрим два рефлекса, которые помогают контролировать мышечную функцию. Первый из них включает особую структуру — нервно-мышечное веретено.
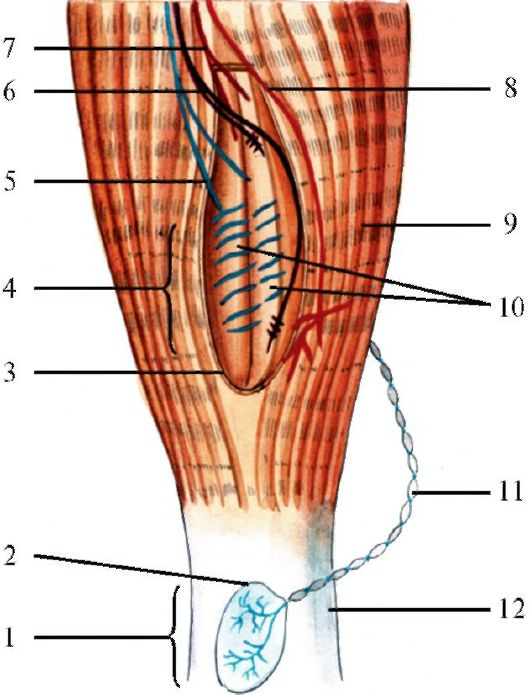
Рис. 33. Схема:Нервно-мышечное веретено Нервно-сухожильное веретено.
1 - сухожильный орган Гольджи; 2 - капсула; 3 - соединительная капсула; 4 - мышечное веретено; 5 - первичное чувствительное окончание (волокна Iа типа); 6 - вторичное чувствительное окончание (волокна II типа); 7 - гамма-эфферентное двигательное волокно к интрафузальным мышечным волокнам; 8 - альфа - эфферентное двигательное волокно к экстрафузальным мышечным волокнам; 9 - экстрафузальные мышечные волокна; 10 - интрафузальные мышечные волокна; 11 - чувствительное волокно; 12 – сухожилие
Дата добавления: 2016-04-02 ; просмотров: 3723 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Читайте также:
