
Дыхательный рефлекс отдел центральной нервной системы
Центральная нервная система (ЦНС) — это основная часть нервной системы у человека и животного, которая состоит из узла нейронов (нервных клеток) и их отростков. Она представлена у человека и позвоночного животного головным и спинным мозгами. У беспозвоночных — системой тесно связанных нервных узлов. Основной задачей и функцией ЦНС является осуществление сложных и простых рефлексов.
- Отделы центральной нервной системы
- Функции ЦНС
- Методы исследования ЦНС и ее функций
- Рефлексы
- Свойства центров нервной системы
- Принципы в координации центральной нервной системы
Отделы центральной нервной системы
Нервная система человека и высших животных состоит из следующих отделов:
- Спинной мозг;
- Продолговатый мозг;
- Средний мозг;
- Промежуточный мозг;
- Мозжечок.
Все эти отделы регулируют деятельность систем у высокоразвитого организма и отдельных органов. Также они связывают их и осуществляют их взаимодействие. Обеспечивают целостность деятельности и единство организма.
Высшими отделами ЦНС являются:
- Кора полушарий (больших) мозга головного;
- Подкорковые ближайшие образования.
Они регулируют взаимоотношения и связь между окружающей средой и организмом как единого целого.
Функции ЦНС
Среди основных функций центральной нервной системы выделяют следующие:
![]()
Координация. Это согласованная работа между различными органами и системами, которую обеспечивает ЦНС. Сюда входят все формы различных движений тела, перемещение организма в пространстве, сохранение определённого положения и позы, деятельность трудовая, а также некоторый ряд приспособительных общебиологических реакций.- Интеграция. Это объединение всех функций организма. Эта функция подразделяется на три вида. Нервная — объединение случается за счёт периферической и центральной нервных систем. Гуморальная — объединяются функции в организме преимущественно с помощью факторов гуморальных. Механическая — отвечает за выполнение функций в организме при наличии целостности органа (если наблюдаются повреждения или переломы в любом органе, то функция считается нарушенной).
- Корреляция. Эта функция обеспечивает взаимосвязь между различными отдельными функциями, органами и системами.
- Регуляция. Сюда входят саморегуляция, различные виды рефлексов, формирование систем функциональных, которые, в свою очередь, обеспечивают положительный приспособительный результат, связанный с изменениями условий внутренней и внешней среды в организме. Регулирующее влияние в центральной нервной системе может проявляться в виде запускающих, корригирующих и трофических обменных процессах.
- Установка и поддержка взаимосвязи окружающей среды и организма.
- Трудовые и познавательные процессы организма. Такие функции отвечают за адекватность организма в условиях окружающей среды.
Методы исследования ЦНС и ее функций
Все методы исследования связаны с интенсивным развитием физиологии центральной нервной системы. Они подразделяются на следующие типы:
- Разрушительный метод. Он связан с изучением того, какие типы функций сохраняются, а какие выпадают после вмешательства оперативного. Сопровождается значительными изменениями в организме и ЦНС.
- Перерезка. С помощью этого метода можно исследовать каково значение любого отдела ЦНС и влияние на него других отделов. Производится на любых уровнях ЦНС.
![]()
Раздражение. С помощью такого метода можно увидеть каково значение функциональное при различных образованиях центральной нервной системы.- Электрографический. Этот метод, в свою очередь, подразделяется на следующие подтипы: электроэнцефалография, локальное отведение потенциалов, вызванные потенциалы.
- Рефлекторные исследования.
- Биохимические исследования.
- Фармакологические.
Классифицируют рефлексы на следующие подтипы в зависимости от их типа:
- Происхождение: врождённые (безусловные) и приобретённые (условные);
- Рецепторы: экстероцептивные, интероцептивные, проприоцептивные;
- Биологические: оборонительные, половые, пищевые;
![]()
Эффекторы: сосудодвигательные, секреторные, двигательные;- Уровень замыкания: корковые, подкорковые, мезенцефалические, бульбарные, спинномозговые;
- Аксон-рефлекс: рефлекторная группа, которая без участия тела осуществляется по аксоновым разветвлениям;
- Функциональные: синергические и антагонистические;
- Сложность пути рефлекса: полисинаптические и моносинаптические;
- Вегетативные — участвуют в регулировании деятельности желез секреции (внутренней), сосудов, внутренних органов;
- Соматические — выявляют себя в виде сокращения мышц (фазного) и в тонусном изменении;
- Адаптационно-трофические: висцеро-моторные, висцеро-висцеральные, висцеро-кутальные.
Нервным центром называется объединение нейронов, которые будут принимать участие в работе одного конкретного рефлекса организма. Во всём организме для того чтобы сформировать адаптивный сложный процесс производится функциональное воссоединение нейронов, которые располагаются на разных уровнях ЦНС.
Нервные центры имеют ряд особенностей и свойств. К таким относятся:
- Возбуждение одностороннее — к органу рабочему от рецептора.
- В центрах нервных возбуждение проявляется медленнее, нежели по нервным волокнам.
- Происходит в нервных центрах и суммация возбуждений. Она может иметь последовательный, одновременный или временный характер.
- Трансформация в ритме возбуждения. Это изменение в количестве импульсов, которые выходят из нервных центров, в сравнении с тем числом, которое приводит к нему. Может проявляться в повышении или понижении количества импульсов.
- Последействие рефлексов — прекращение реакции чуть позже по сравнению с действием возбудителя.
- Повышенная чувствительность к веществам химического происхождения и кислородному недостатку.
- Нервные центры быстро утомляемы и имеют низкий уровень локальности, легко тормозятся.
- Нервные центры имеют пластичную структуру — могут изменять своё функциональное предназначение и восстанавливать частично функции, которые были утрачены.
Основой координационной деятельности нервной системы является взаимодействие процессов торможения и возбуждения. Существует ряд принципов, обеспечивающих координационное взаимодействие:
- Принцип доминанты. Он может быть охарактеризован такими свойствами: инертностью возбуждения, повышенным уровнем возбудимости, суммацией возбуждений, торможением субдоминантных очагов возбуждений, исходящих от других центров.
- Принцип окклюзии. Смысл этого принципа состоит в том, что пара афферентных входов вместе возбуждают более малую группу мотонейронов в сравнении с эффектом их отдельной активации.
- Принцип связи обратного порядка. Полноценно в организме процесс саморегуляции осуществляется только при полном функционировании обратного канала связи.
![]()
Принцип реципрокности (взаимообусловленности, сопряжения). Отображает отношения между теми центрами, которые несут ответственность за осуществление функций, которые являются противоположными.- Принцип конечного общего пути. Нейроны эффекторные ЦНС вовлекаются в осуществление разных реакций возбуждения в организме, которые приводят к ним большое количество промежуточных и афферентных нейронов, для которых они будут служить конечным путём.
- Явления конвергенции. Это процесс, при котором нервные импульсы сходятся на одни центральные нейроны.
- Явления дивергенции. Это процесс, при котором импульсы расходятся по соседним участкам.
- Взаимоотношения субординационные. Процесс, при котором верхние отделы ЦНС влияют на нижние отделы ЦНС.

Регуляция дыхания и другие функции организма
Дыхательный центр продолговатого мозга обеспечивает такую вентиляцию легких, которая необходима для поддержания на оптимальном уровне напряжения кислорода и углекислого газа. Напряжение этих газов, воздействуя на дыхательный центр через хеморецепторы, вызывает ответную реакцию дыхания, направленную на устранение отклонения в концентрации. Таким образом осуществляется регуляция дыхания по принципу отклонения регулируемого параметра от нормальных значений.
Вместе с тем, изменения вентиляции легких наблюдаются при самых разнообразных ситуациях, когда нет изменения напряжения кислорода и углекислого газа. Например, воздействия холода или тепла на кожу приводят к возбуждению дыхательного центра и учащению дыхания. Кроме того, на дыхание влияет изменение температуры тела: и понижение, и незначительное повышение вызывают увеличение вентиляции легких. Весьма существенно увеличивает частоту дыхания боль. Вызывают изменения дыхания и физическая, и эмоциональная нагрузки. Такое изменение паттерна дыхания, не обусловленное изменением концентрации газов в крови, является проявлением варианта регуляции по принципу возмущения. Это означает, что тем сигналом, который поступает в дыхательный центр и вызывает гипервентиляцию, служит не отклонение в газовом составе крови, а сигнал о происходящем в организме возмущении, способном привести к отклонению в газовом составе крови, регуляция осуществляется до того, как произойдут отклонения. Механизмы возмущающих влияний на дыхательный центр становятся понятны, если вспомнить о том, что дыхательный центр входит в состав ретикулярной формации ствола мозга.
Ретикулярная формация ствола мозга
Под ретикулярной формацией обычно понимают клеточную массу, лежащую в толще мозгового ствола от нижних отделов продолговатого до промежуточного мозга. Эта клеточная масса слабо структурирована и не имеет четких границ. Внутри ретикулярной формации расположены чувствительные и двигательные ядра продолговатого, среднего и промежуточного мозга. Нейроны ретикулярной формации характеризуются немногочисленными длинными и мало ветвящимися дендритами, их шипики слабо дифференцированы. В медиальной части ретикулярной формации расположены крупные и гигантские клетки, в продолговатом мозге они сконцентрированы в гигантоклеточном ядре. Именно от этих клеток отходят аксоны, которые формируют эфферентные пути. В частности, ретикулоспинальный тракт, пути к таламусу, мозжечку, базальным ганглиям и коре больших полушарий.
Сетевое строение ретикулярной формации обеспечивает высокую надежность ее функционирования и устойчивость к повреждающим воздействиям, потому что локальные повреждения всегда компенсируются за счет сохранившихся элементов сети. Такое сетевое строение обеспечивает и еще одну важную особенность функционирования ретикулярной формации: раздражение любой из ее частей за счет многочисленных связей охватывает всю данную структуру. Кроме того, эффекты стимуляции, как правило, оказываются весьма длительными за счет свойства нейронной сети пролонгировать возбуждение.
Гипоталамус
Существенную роль в регуляции дыхания играет гипоталамус. Основная роль гипоталамуса в изменении паттерна дыхания заключается в том, что ритм дыхания приводится в соответствие с уровнем обмена веществ. Одной из основных особенностей нейронов гипоталамуся является их чувствительность к составу омывающей их крови. Нейроны срединной группы ядер гипоталамуса обладают детектирующей функцией, они реагируют на изменение температуры крови, осмотическую концентрацию, концентрацию глюкозы и аминокислот. Таким образом, в гипоталамус поступает информация о состоянии внутренней среды организма. В гипоталамусе на основании этой информации происходит формирование мотиваций - побуждений к движению, к изменению поведения для того, чтобы восстановить изменившиеся параметры внутренней среды организма. Под влиянием мотивации и при участии коры головного мозга происходит формирование программы конкретного поведения, в которую включается и изменение дыхания.
Особенно велика роль дыхания в терморегуляции, потому что при изменении паттерна дыхания возможно или увеличить, или уменьшить теплоотдачу. При повышении температуры повышается частота дыхания, за счет этого происходит увеличение испарения воды и некоторое охлаждение организма. Умеренное охлаждение тоже приводит к увеличению частоты дыхания. Это связано с возбуждением задних областей гипоталамуса и общим повышением активности и возбудимости структур мозга, в том числе и дыхательного центра. При значительном охлаждении угнетается активность нейронов дыхательного центра, глубина и частота дыхания уменьшаются, что позволяет уменьшить потери тепла.
Из изложенного становится ясно, что в дыхательный центр, наряду с информацией от хеморецепторов и от рецепторов растяжения легких, поступает поток информации обо всем, что происходит в организме и окружающей среде. Разница заключается в том, что афферентные сигналы от разнообразных рецептивных полей и от анализаторов поступают не непосредственно в дыхательный центр, а в различные отделы головного мозга. Эти отделы, в свою очередь, оказывают возбуждающее или тормозное влияние и на дыхательный центр, и на другие функциональные системы (например, система кровообращения). В различных ситуациях центры головного мозга образуют с дыхательным центром временные функционально подвижные ассоциации (функциональные системы по А.П. Анохину), которые обеспечивают полноценное регулирование дыхания в соответствии с потребностями организма.
Кора больших полушарий
Роль коры больших полушарий в регуляции дыхания изучена в экспериментах на животных с электрическим раздражением различных зон коры и с их удалением. Оказалось, что у бескорковых животных с нормальным дыхательным ритмом в покое наступает резко выраженная и длительная одышка уже после нескольких шагов. Следовательно, для приспособления дыхания к мышечной деятельности требуется участие высших отделов центральной нервной системы. В коре мозга нет определенных участков, четко изменяющих деятельность дыхательного центра. Раздражение большинства участков коры приводит к изменению паттерна дыхания. Вместе с тем, наиболее существенные изменения дыхания наблюдаются при раздражении соматосенсорной области коры. Это и понятно, именно мышечная деятельность требует наиболее существенного изменения дыхания. Во время работы мышцам необходимо очень большое количество кислорода. Если в покое потребление кислорода составляет 250-300 мл/минуту, то при быстрой ходьбе оно возрастает до 2.5 литров, а при тяжелой мышечной работе до 4л/минуту. Обеспечение мышц кислородом достигается совместной деятельностью систем кровообращения и дыхательной.
Механизмы регуляции вентиляции при мышечной работе сложны. Казалось бы, увеличение МОД можно объяснить повышением частоты импульсов с хеморецепторов продолговатого мозга и каротидных синусов, которое возникает в результате повышения напряжения углекислого газа и снижения напряжения кислорода в крови. Однако вентиляция легких всегда увеличивается в начале работы, когда газовый состав крови еще не успел измениться. Следовательно, гиперпноэ возникает под влиянием нервных факторов. Кора больших полушарий, вызывая произвольные движения, активирует и деятельность дыхательного центра и непосредственно, и через гипоталамус. В дополнение к этой стимуляции возбуждение дыхательного центра увеличивается под влиянием импульсов от проприорецепторов работающих мышц. Эти импульсы возникают даже в тех случаях, когда работа мышц осуществляется пассивно, без затрат кислорода и выделения углекислого газа (например, экспериментатор сгибает конечности испытуемого).
Позднее, во время продолжающейся физической работы, происходит более медленное увеличение вентиляции легких. Это увеличение вентиляции уже обусловлено раздражением артериальных и центральных хеморецепторов. Однако и здесь не все так просто. Выяснилось, что даже при физической работе высокой интенсивности напряжение кислорода и углекислого газа в крови может не измениться (а углекислого газа даже снизиться в результате возросшей вентиляции легких). Даже удаление каротидных телец не устраняет увеличения легочной вентиляции во время физической работы, и тем не менее, сигналы от хеморецепторов имеют существенное значение в увеличении МОД во время физической работы. Оказывается, что во время работы увеличивается чувствительность дыхательного центра к гиперкапнии и гипоксии, возрастает и возбудимость хеморецепторов. Имеет значение и повышение температуры тела: этот фактор через центры гипоталамуса увеличивает частоту дыхания.
Кортикальные влияния на дыхание отчетливо проявляются при тренировке к выполнению одной и той же работы. Постепенно колебания дыхания становятся меньше, дыхание делается более ровным. Если много раз выполняется работа в одинаковом темпе, но с различной интенсивностью, то изменения вентиляции при переходе на новый уровень происходят быстрее, вырабатывается динамический стереотип, в котором вентиляция легких имеет волнообразный характер. Из этого примера видно, что важные приспособительные изменения дыхания осуществляются посредством выработки условных рефлексов. Примером такого условно-рефлекторного изменения ритма дыхания может быть стартовый рефлекс у спортсменов.
Особенно отчетливо роль коры головного мозга проявляется в произвольном управлении дыханием. Своеобразие функции внешнего дыхания заключается в том, что она одновременно и обладает автоматизмом, и произвольно управляема. Человек прекрасно дышит и во сне, и под наркозом, однако любой человек может произвольно остановить дыхание, изменить его частоту и глубину. Произвольное управление дыханием возможно потому, что в коре есть представительство дыхательных мышц и корковомедуллярные нисходящие влияния на мотонейроны дыхательных мышц. Возможность произвольного управления дыханием ограничена определенными пределами изменений напряжения кислорода и углекислого газа, а так же рН крови. При чрезмерной задержке дыхания возникает стимул, который возвращает дыхание под контроль дыхательного центра. Значение возможности произвольного контроля дыхания для человека трудно переоценить - именно благодаря такой возможности человек приобрел одну из немногих, отличающих его от животных, функций - способность к членораздельной речи.
Заключение
Дыхательный центр получает большой объем информации и о газовом составе крови, и о состоянии дыхательной системы. В соответствии с этой информацией изменяется легочная вентиляция. Вместе с тем дыхательный центр способен обеспечивать смену дыхательных фаз за счет своих собственных механизмов возбуждения нейронов, т.е. обладает определенным автоматией. Автоматия дыхательного центра отличается рядом признаков от автоматии сердечного водителя ритма.
Периодическое возбуждение дыхательного центра обусловлено взаимодействием большого числа нервных клеток, объединенных в нейронные сети. Среди них особую роль играют тормозные нейроны. В дыхательном центре есть несколько популяций инспираторных и экспираторных нейронов соответственно фазам дыхательного цикла. Среди нейронов есть проприобульбарные нейроны, которые организуют паттерн дыхания, определяя уровнем своего возбуждения длительность фаз дыхательного цикла. Бульбоспинальные нейроны являются преимущественно премоторными по отношению к мотонейронам диафрагмального нерва и нервам межреберных мышц. Смена дыхательных фаз осуществляется путем постепенного возбуждения одних групп нейронов и торможения других. Для осуществления смены дыхательных фаз необходимо влияние пневмотаксического центра, расположенного в районе варолиевого моста среднего мозга.
Для автоматической деятельности дыхательного центра необходимо постоянное (тоническое) поступление к нему сигналов, повышающих возбудимость дыхательных нейронов. Сигналы, поступающие от хеморецепторов продолговатого мозга и каротидного синуса, а так же от рецепторов растяжения легких изменяют активность дыхательных нейронов в соответствии с газовым составом крови и состояние дыхательной системы. Информация от этих рецепторов лежит в основе регуляции по принципу отклонения. Афферентные сигналы, поступающие от всех рецептивных полей через ретикулярную формацию, гипоталамус и кору головного мозга изменяют активность дыхательных нейронов в соответствии с состоянием внутренней среды организма и окружающей среды, поведением. Таким образом, осуществляется регуляция дыхания по принципу возмущения.
Автоматическая деятельность дыхательного центра человека находится под значительным произвольным контролем.Человек может в широких пределах произвольно изменять частоту и глубину дыхания. Вместе с тем, этот произвольный контроль ограничен влияниями от центральных и периферических хеморецепторов, рецепторов растяжения легких, ирритантных и других механорецепторов.
Объединим на рисунке 25 основные структуры, принимающие участие в регуляции дыхания и по принципу отклонения, и по принципу возмущения, и суммируем их роль в регуляции дыхания.
Рисунок 25. Отделы головного мозга,
Теория по нормальной физиологии. Тема: Регуляция дыхания. Дыхательный центр, опыты Фредерика и Холдена, действие карбогена, хеморецепторы, механорецепторы..
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Регуляция дыхания осуществляется ЦНС.
Спокойное дыхание взрослого человека характеризуется последовательной сменой актов вдоха и выдоха с частотой 14-16 дыханий в минуту.
Чтобы произошел вдох, необходимо сокращение дыхательных мышц.
Импульсы к ним поступают от мотонейронов передних рогов спинного мозга.
К диафрагме – от III – IV шейных сегментов, к межреберным мышцам – от грудных сегментов спинного мозга.
Мотонейроны получают импульсы от дыхательного центра , расположенного в продолговатом мозге.
Если перерезать спинной мозг под продолговатым мозгом , то дыхание – прекращается .
Дыхательный центр
Был открыт в 1812 г. Легаллуа и в 1842 г. Флурансом, которые своими опытами доказали его локализацию в продолговатом мозге .
Н.А.Миславский в 1885 г. уточнил местоположение дыхательного центра – в области РФ (ретикулярной формации) продолговатого мозга , т.к. перерезка между грудными и шейными сегментами спинного мозга – сохраняет диафрагмальное дыхание:
- продолговатым и спинным – полностью прекращает дыхание ,
- выше продолговатого мозга – дыхание сохраняется .
Н.А.Миславским также было показано, что дыхательный центр состоит из двух отделов:
- инспираторного (отвечает на вдох),
- экспираторного (отвечает за выдох).
Они находятся в реципрокных (то есть в противоположных) отношениях.
В настоящее время установлено:
- в РФ продолговатого мозга находятся инспираторные и экспираторные нейроны ,
- эти нейроны рассеяны диффузно , среди других нейронов РФ и относительно друг друга,
- инспираторных нейронов примерно вдвое больше , чем экспираторных.
В продолговатом мозге есть еще два скопления дыхательных нейронов – дорсальные и вентральные дыхательные ядра.
Дорсальное ядро содержит, в основном, инспираторные нейроны, контролирующие сокращение диафрагмы .
В вентральном ядре содержатся как инспираторные , так и экспираторные нейроны, которые включаются при форсированном дыхании.
Все дыхательные нейроны делят на 6 групп:
- ранние инспираторные – активны в начале фазы вдоха (инспирации),
- поздние инспираторные – активны в конце вдоха,
- полные инспираторные – активны в течение всего вдоха,
- постинспираторные – максимальный разряд в начале выдоха,
- экспираторные – активны во вторую фазу выдоха,
- преинспираторные – активны перед вдохом, они включают активную экспирацию (выдох.
Значение дыхательных нейронов:
- Преинспираторные и ранние инспираторные влияют на момент начала вдоха.
- Инспираторные нейроны ДЦ генерируют дыхательный ритм (частоту и глубину дыхания), иннервируют мотонейроны.
- Постинспираторные нейроны контролируют процесс пассивного выдоха.
- Экспираторные нейроны отвечают за активный выдох, так как иннервируют мотонейроны внутренних межреберных мышц и мышц передней брюшной стенки.
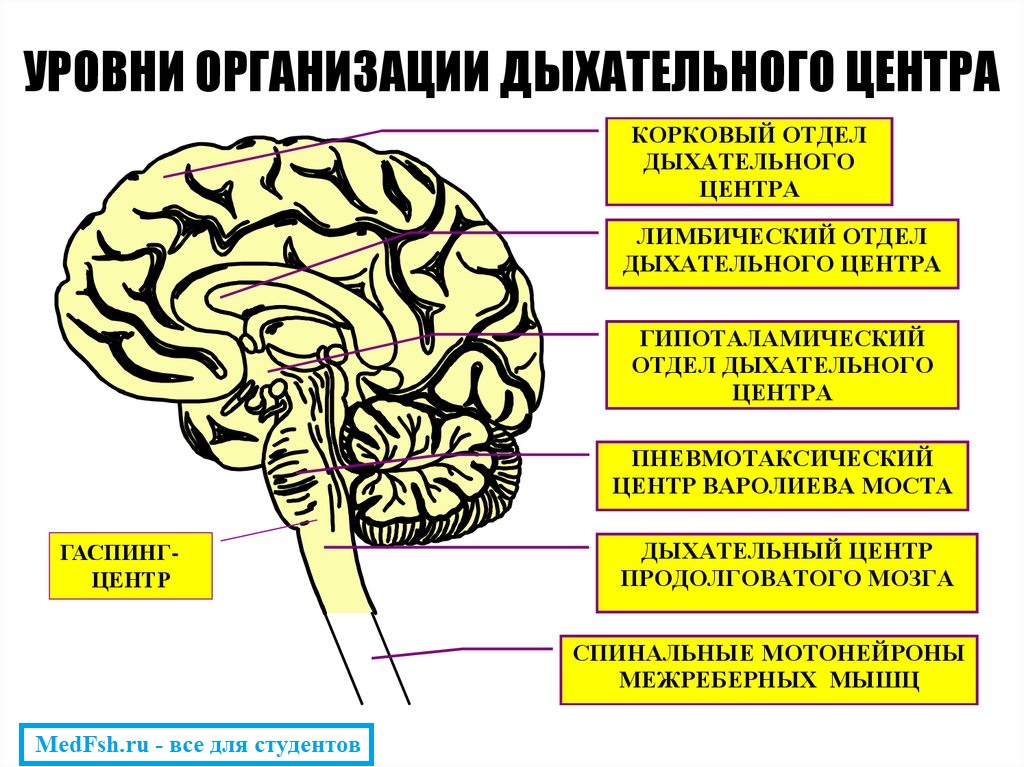
Дыхательный центр – это совокупность нервных клеток, расположенных в различных отделах ЦНС и принимающих участие в регуляции дыхания .
У двух собак, находящихся под наркозом, перекрестно соединяли сонные артерии и яремные вены, латерально же расположенные сосуды пережимали. При этом голова 1-й собаки снабжались кровью из туловища 2-й и наоборот.
У первой собаки кратковременно пережимали трахею, и у нее в крови уменьшалось содержание кислорода (гипоксемия) и увеличивалось содержание углекислого газа (гиперкапния). Эта кровь поступала в голову 2-й собаки, и у нее наступала одышка (диспноэ).
В результате у нее в крови увеличивалось содержание кислорода (гипероксимия) и уменьшалось содержание углекислого газа (гипокапния), и эта кровь поступала в голову 1-й собаки, и у нее наступало апноэ – остановка дыхания. (Нормальное дыхание — эйпноэ).
На состояние ДЦ влияет газовый состав крови:
При увеличении в крови напряжения углекислого газа и уменьшении кислорода, ДЦ – возбуждается и, наоборот, уменьшается , если в крови уменьшается напряжение углекислого газа и увеличивается напряжение кислорода.
При дыхании в герметичной камере в воздухе увеличивалось содержание CO2 и уменьшалось O2 – наступала одышка . Когда CO2 поглощается натронной известью, одышка наступала намного позже, хотя содержание O2 в воздухе значительно снижалось .
Одышка наступает:
- При снижении O2 в атмосферном воздухе с 20,94% до 12%, т.е. на 9%.
- При повышении содержания CO2 в альвеолярном воздухе на 0,17% вентиляция удваивается.
Главный активатор ДЦ – CO2
Действие карбогена
Карбоген – газовая смесь, состоящая из 96% — O2, 4% — CO2.
В сравнении с воздухом в карбогене в 4,8 раза больше O2 и в 130 раз – CO2.
Карбоген применяют при расстройствах дыхания.
Эффект карбогена связан с эффектами содержащегося в нем CO2:
- стимуляция ДЦ,
- расширение бронхов и кровеносных сосудов,
- сдвиг кривой диссоциации HbO2 вправо –> увеличение диффузии O2 из крови в ткани.
Карбоген применяется в медицинских целях для лечения горной болезни, отравления угарным газом, глаукомы, стресса, при восстановлении слуха после воздействия шума и в ряде случаев для улучшения кровоснабжения опухолей при химио- и лекарственной терапии.
В 1911 г. Винтерштейн доказал, что возбудителем ДЦ также являются ионы Н .
Ацидоз – усиливает легочную вентиляцию.
Он показал, что возбуждают ДЦ нелетучие кислоты – молочная, никотиновая и другие.
Самым сильным стимулятором дыхания являются:
- pCO2 (гиперкапния),
- pH (ацидоз),
- pO2 (гипоксемия).
Механизм действия гуморальных факторов (CO2, O2, H) на ДЦ :
- Через хеморецепторы (периферические) сосудистых рефлексогенных зон.
- Через хеморецепторы, находящиеся в продолговатом мозге (медуллярные).
- расположены в каротидных и в аортальных тельцах,
- реагируют на (в артериальной крови):
- увеличение pCO2,
- уменьшение pO2,
- увеличение H (от есть уменьшение pH).
Хеморецепторы возбуждаются постоянно CO2 и O2 , растворенными в крови, а также H, т.к. порог для pCO2 равен 20-30 мм.рт.ст. В норме pCO2 = 40 мм.рт.ст.
Порог для pO2 равен 130-140 мм.рт.ст. В норме pO2 = 100 мм.рт.ст. Одышка же наступает при pO2 ниже 50-60 мм.рт.ст.
Таким образом, хеморецепторы постоянно посылают импульсы в ДЦ, возбуждая инспираторные нейроны, причем большую роль играют хеморецепторы каротидного синуса.
- Находятся на вентролатеральной поверхности продолговатого мозга .
- Реагируют только на H и изменение напряжения CO2.
- Эти рецепторы возбуждаются позднее, поскольку требуется время для проникновения CO2 через гематоэнцефалический барьер.
- Импульсы, поступающие с медуллярных хеморецепторов в ДЦ, увеличивают прирост вентиляции на 60-80% .
Находятся в:
- легких,
- дыхательных путях,
- дыхательных мышцах (проприорецепторы)
Это барорецепторы рефлексогенных зон.
Механорецепторы легких (РРЛ)
Рецепторы растяжения легких (РРЛ) – являются наиболее значимым среди всех механорецепторов.
В 1868 г. Геринг и Брейер доказали наличие в легких рецепторов, которые возбуждаются при их растяжении, то есть при вдохе. Они являются чувствительными окончаниями блуждающих нервов, которые направляют свои импульсы в ДЦ.
Геринг и Брейтер раздували легкие и наблюдали прекражение вдоха (инспираторно-тормозной рефлекс).
Таким образом, этот рефлекс способствует смене вдоха на выдох. Он называется рефлексом Геринга-Брейера и является рефлексом саморегуляции дыхания.
При перерезке блуждающих нервов, дыхание становится редким и глубоким, альвеолы расширяются до максимального предела, т.к. вдох не тормозится. В этом случае, смене вдоха на выдох будет способствовать пневмотаксический центр (ПТЦ).
В настоящее время известно, что в легких существует 3 разновидности механорецепторов :
- РРЛ медленноадаптирующиеся,
- РРЛ быстроадаптирующиеся или ирритантные,
- Юкстаальвеолярные рецепторы капилляров.
РРЛ медленноадаптирующиеся:
- Они расположены в ГМК (гладкомышечных клетках) дыхательных путей.
- Возбуждаются при вдохе.
- С них осуществляется рефлекс Геринга-Брейера.
Ирритантные рецепторы (быстроадаптирующиеся):
- Расположены в слизистой дыхательных путей.
- Реагируют на механические и химические стимулы.
- Быстро адаптирующиеся.
- Длительное раздражение этих рецепторов приводит к хроническому бронхиту.
Физиологическое значение при вдыхании токсических веществ: Сужение бронхов -> вентиляция альвеол -> поступление этих веществ в альвеолы и кровь.
- расположены в паренхиме легких, в альвеолярных перегородках, прилегающих к капиллярам,
- стимулируются, главным образом, растяжением легочных сосудов,
- быстро реагируют на введение химических веществ в легочные сосуды,
- стимуляция может вызвать апноэ, затем учащение дыхания, уменьшение давления, брадикардию и бронхоспазм.
Возбуждение механорецепторов верхних ДП вызывает возникновение дыхательных рефлексов.
Защитные дыхательные рефлексы:
- Чихание – с рецепторов слизистой носа.
- Кашель – с ирритантных рецепторов слизистой гортани, трахеи, бронхов.
- Рефлекс ныряльщиков – остановка дыхания при действии воды на носовые ходы.
- Остановка дыхания во время акта глотания .
- Рефлексогенная задержка дыхания – сужение голосовой щели, бронхоконстрикция при вдыхании дыма, газов, едких веществ.
Механорецепторы межреберных мышц и диафрагмы (проприорецепторы) – они осуществляют обратную связь дыхательных мышц с ДЦ.
Барорецепторы рефлексогенных зон – возбуждаются они при увеличении АД и дыхание при этом угнетается. При уменьшении АД – дыхание усиливается.
Ритмическая смена дыхательных фаз
Хеморецепторы возбуждаются постоянно и постоянно посылают импульсы в продолговатый мозг, возбуждая α-инспираторные нейроны. Они возбуждаются и посылают импульсы к мотонейронам спинного мозга.
Мотонейроны возбуждаются и посылают импульсы к мышцам. Они сокращаются и наступает вдох .
При вдохе объем легких увеличивается и возбуждаются РРЛ, которые посылают возбуждающие импульсы по чувствительным волокнам блуждающего нерва к β-инспираторным нейронам. В результате суммации импульсов от α-инспираторных нейронов и рецепторов растяжения легких достигается порог и возбуждаются и β-инспираторные нейроны благодаря влиянию вышележащих отделов ЦНС.
Инспираторно-тормозные нейроны посылают тормозные импульсы к α-инспираторным нейронам. В результате α-инспираторные нейроны тормозятся и не посылают импульсы к мотонейронам. Мышцы расслабляются, происходит выдох .
К β-инспираторным нейронам не поступают импульсы, и они тормозятся (не возбуждаются).
β-инспираторные нейроны не возбуждают инспираторно-тормозные нейроны и поэтому они не посылают импульсы к α-инспираторным нейронам. α-инспираторные нейроны вновь возбуждаются импульсами от хеморецепторов и наступает вдох .
Читайте также:
