
Эфферентные нервы дыхательной системы

Дыхательный центр (его компоненты) и эфферентные нервы:
К - кора; Гт — гипоталамус; Пм — продолговатый мозг; См - спинной мозг; Th, -Th,; - грудной отдел; Сд-Сд - отдел спинного мозга
Дыхательный центр управляет двумя основными функциями: двигательной, которая проявляется в виде сокращения дыхательных мышц, и гомеостатической, связанной с поддержанием постоянства внутренней среды организма при сдвигах в ней содержания 02 и СО2.
Понятие о дыхательном центре появилось еще в XVIII в., когда было обнаружено прекращение дыхания у животных при разрушении ограниченной области продолговатого мозга. Однако современное представление о структуре и функции центрального дыхательного механизма сложилось лишь в последние десятилетия в результате использования электрофизиологических методов исследования,позволивших выявить определенные группы так называемых дыхательных нейронов. К дыхательным нейронам относят нервные клетки, импульсная активность которых меняется в соответствии с фазами дыхательного цикла. Различают инспираторные нейроны, которые разряжаются в фазу вдоха, экспираторные, активные во время выдоха, и целый ряд нейронных популяций, активность которых или занимает часть определенной фазы дыхательного цикла (ранние, поздние), или включается в моменты перехода инспираторной фазы в экспираторную либо экспираторной в инспираторную.
Центральный дыхательный механизм входит в состав ретикулярной формации ствола мозга. Подавляющая масса дыхательных нейронов сосредоточена в двух группах ядер: дорсальной и вентральной. Большая часть нейронов дорсальной группы - инспираторные, бульбоспинальные. Их аксоны направляются в шейные сегменты спинного мозга и образуют синапсы с мотонейронами диафрагмального ядра. Эти нейроны непосредственно управляют сокращением диафрагмы.Ядра вентральной дыхательной группы содержат инспираторные и экспираторные нейроны. Последние связаны преимущественно с мотонейронами межреберных и брюшных мышц, расположенных в грудных и поясничных сегментах спинного мозга, частично с мотонейронами диафрагмы, обеспечивая дыхательную активность указанных мышц.Активность центрального дыхательного механизма, в свою очередь, управляется стимулами, исходящими от хеморецепторов и механорецепторов дыхательной системы. Главная особенность работы этого механизма - линейное нарастание активности инспираторных нейроновна протяжении вдоха и резкий обрыв инспираторной активности, знаменующий окончание вдоха и переход в выдоху. Чем сильнее импульсация от хеморецепторов, тем круче нарастает инспираторная активность и быстрее развивается вдох, но так как при этом резче растягиваются легкие, то вдох быстрее сменяется выдохом. В итоге увеличивается и глубина, и частота дыхания. Полному расслаблению инспираторных мышц предшествует плавное снижение их активности, обусловленное, как полагают, включением особой группы нейронов, которые оказывают тормозящее ("запирающее") влияние на инспираторную и экспираторную активность. Эту фазу назвали постинспираторной. По-видимому, постинспираторная фаза обеспечивает интервалы, необходимые для опорожнения легких после очередного вдоха.
Таким образом, центральный паттерн дыхания включает три фазы:
Нейроны, связанные с регуляцией дыхания, имеются и в варолиевом мосту. Здесь выделяют так называемый пневмотаксический центр, который участвует в переключении фаз дыхательного цикла; при разрушении этого центра вдохи становятся затянутыми, необычно глубокими. Центральный дыхательный механизм продолговатого мозга обладает автоматией, то есть постоянной ритмической активностью. Однако это его свойство у высших позвоночных полностью отлично от автоматии, свойственной, например, узлам проводящей системы сердца, возбуждение которых происходит в силу их внутренних свойств. Дыхательные нейроны функционируют нормально лишь при двух условиях. Первым условием является сохранность связей между их различными группами (хотя пока не установлено, какие именно нейроны являются водителями ритма, пейсмекерами, и существуют ли среди них такие пейсмекеры вообще). Вторым условием - наличие афферентной стимуляции. В этом плане важнейшую роль играет импульсация, поступающая от хеморецепторов.
ХЕМОРЕЦЕПТОРЫ. ХЕМОРЕЦЕПТОРНЫЕ СТИМУЛЫ ДЫХАНИЯ.
1.Бульбарные хемочувствительные зоны. На вентролатериальной поверхности продолговатого мозга расположены нейрональные структуры, чувствительные к напряжению СО2 и концентрации ионов Н + во внеклеточной жидкости мозга. Локальное воздействие этих факторов вызывает увеличение дыхательного объема и легочной вентиляции. Напротив, снижение PСО2 и подщелачивание внеклеточной жидкости мозга, а также блокирование холодом или разрушение этих структур ведет к полному или частичному исчезновению реакции дыхания на избыток CO2 (гиперкапнию) и ацидоз, а также к резкому угнетению инспираторной активности вплоть до остановки дыхания.
2. Артериальные хеморецепторы. В области бифуркации сонной артерии расположено так называемое каротидное тело (каротидный клубочек, гломус). Оно обильно снабжается кровью и содержит сложно устроенный рецепторный аппарат, реагирующий на изменения газового состава артериальной крови:
•снижение напряжения 02 (гипоксемию),
•повышения напряжения CO2 (гиперкапнию),
•увеличение концентрации ионов Н + (ацидоз).
Все три фактора, вызывая возбуждение каротидных хеморецепторов, усиливают активность центрального дыхательного механизма. Особенно важна чувствительность этих рецепторов к гипоксемии, ибо они являются единственными в организме сигнализаторами о недостатке кислорода. Афферентные пути от каротидного тела идут через синусный нерв (ветвь языкоглоточного) и достигают дорсальной дыхательной группы продолговатого мозга.
3.Хеморецепторные стимулы дыхания. В настоящее время установлено, что нейроны центрального дыхательного механизма прямой чувствительностью к химизму среды не обладают, а их активность определяется главным образом импульсами от хеморецепторов, прежде всего бульбарных. Главным стимулом, управляющим дыханием, служит гиперкапнический:
чем выше напряжение СО2 (а с этим параметром связана и концентрация ионов Н + в артериальной крови и внеклеточной жидкости мозга), тем сильнее возбуждение бульбарных хемочувствительных структур и артериальных хеморецепторов и тем выше вентиляция.
Так, если человек дышит из того же мешка, куда выдыхает (возвратное дыхание), то по мере роста PСО2 в дыхательной среде увеличивается легочная вентиляция. Напротив, если усиленно провентилировать легкие животного, резко снизив таким образом артериальное PСО2, т.е. вызвав гипокапнию, дыхательные движения прекращаются, пока в крови не восстановится нормальный уровень напряжения CO2.
Меньшее значение в регуляции дыхания имеет гипоксический стимул. Его выключение с помощью дыхания кислородом, либо денервации каротидных тел лишь немного (и то не всегда) снижает легочную вентиляцию. Во время дыхания газовыми смесями с пониженным содержанием О2 благодаря рост активности артериальных хеморецепторов вентиляция нарастает, однако зависимость ее от PО2 нелинейна. Эта особенность связана с тем, что в соответствии с формой кривой диссоциации оксигемоглобина крутое падение содержания О2 в крови наступает лишь тогда, когда PО2 опускается ниже 80-70 мм рт. ст. – в этот момент и происходит значительный подъем легочной вентиляции.
Особенно сильным стимулом для центрального дыхательного механизма является сочетание гиперкапнии с гипоксемией (и связанным с ней ацидозом).
Функция центральных и артериальных хеморецепторов заключается в поддержании газового и кислотно-основного гомеостаза организма и прежде всего наиболее требовательной к постоянству химизма среды ткани мозга. Недаром рецепторы каротидного тела контролируют химизм крови, снабжающей головной мозг, а бульбарные хемочувствительные структуры - химизм внеклеточной жидкости самого мозга.
МЕХАНОРЕЦЕПТОРЫ ДЫХАТЕЛЬНОЙ СИСТЕМЫ
Механорецепторы дыхательной системы выполняют двоякую роль:
во-первых, они участвуют в регуляции параметров дыхательного цикла – глубины вдоха и его длительности,
во-вторых, эти рецепторы служат источником ряда рефлексов защитного характера - кашля.
Рецепторы растяжения легких. Эти рецепторы локализованы главным образом в гладкомышечном слое стенок трахеобронхиального дерева и чувствительны к трансмуральному давлению, т.е. к разности давлений внутри и снаружи просвета воздухоносных путей. Тем самым эти механорецепторы оказываются источником сигнализации о растяжении дыхательных путей и легких. Афферентные волокна рассматриваемых рецепторов идут в составе блуждающего нерва, центральный путь их импульсации идет до области дорсальной группы ядер продолговатого мозга. Возбуждение рецепторов растяжения легких, нарастая в ходе вдоха, вызывает в конечном счете торможение активности инспираторных нейронов центрального дыхательного механизма, способствуя прерыванию вдоха и смене его выдоха (рефлекс Геренга - Брейера). Так как активность одной части этих рецепторов зависит от достигнутого легочного объема, а другой части ("динамических") - от скорости вдоха, то прекращение вдоха наступает тем скорее,чем глубже данный вдох и чем быстрее он развивается.
Так регулируется паттерн дыхания - соотношение между глубиной и частотой дыхания.
Роль рецепторов растяжения легких наглядно выявляется при их выключении с помощью блокады или перерезки блуждающих нервов (ваготомии): вдохи становятся углубленными, затянутыми, как это происходит при разрушении пневмотоксического центра. Если же разрушение названного центра сочетать с ваготомией, наступает апноэ: дыхательные движения останавливаются на вдохе, который лишь изредка прерывается короткими экспирациями.
Ирритантные рецепторы расположены в эпителиальном и субэпителиальном слоях стенок воздухоносных путей. Их афферентные волокна тоже идут в стволе блуждающего нерва. Эти рецепторы реагируют на резкие изменения объема легких, в частности на их спадение, которое вызывает рост инспираторной активности центрального механизма, прерывая таким путем выдох. Чувствительны они и к частицам пыли, скоплению слизи, некоторым химическим раздражителям.
Возбуждение ирритантных рецепторов характерно для многих патологических изменений в легких и воздухоносных путях.
Юкстаальвеолярные (юкстакапилярные), или J - рецепторы.
Эти рецепторы чувствительны к ряду биологически активных веществ (никотину, гистамину, простагландинам и др.), проникающим либо из воздухоносных путей, либо с кровью малого круга. Они локализованы в интерстиции легких вблизи капилляров альвеол и дают начало немиелинизированным С-волокнам, проходящим в составе блуждающего нерва. Стимуляция как ирритантных, так и J-рецепторов проявляется в частом, поверхностном дыхании (тахипноэ), которое наблюдается, например, при ограничении (рестрикции) газообменной поверхности легких.
Рецепторы верхних дыхательных путей. Они служат главным образом источником рефлексов защитного характера, возникающих при скоплении в воздухоносных путях слизи, попадании туда инородных тел и химических раздражителей. Эти реакции опосредованы афферентными волокнами целого ряда черепно-мозговых нервов: верхне- гортанного, языкоглоточного, тройничного и т.д.
К рефлексам защитного характера относится прежде всего кашель. Чихание вызывается обычно изолированным раздражением рецепторов носа и отличается от кашля тем, что голосовая щель с самого начала остается открытой. Глотаниесопровождается рефлекторным закрытием голосовой щели и торможением дыхательной активности диафрагмы. Остановка дыхательных движений (апноэ) может происходить и за счет рефлексов, возникающих при попадании в воздухоносные пути воды (или при погружении головы в воду), воздействие струи воздуха, особенно холодного, и т.п.
Проприоцепторы дыхательных мышц. Эти рецепторы контролируют соответствие сокращений "заданию", поступающему по эфферентным путям от центрального дыхательного механизма. Если укорочение мышцы оказывается меньше заданного (например, вследствие повышенного сопротивления дыханию), импульсация от интрафузальных волокон усиливается и через посредство гамма-петли повышается активность соответствующих спинальных альфа-мотонейронов. Сокращение мышцы усиливается, что способствует преодолению возникшего препятствия. Основной инспиратор – диафрагма – беден мышечными веретенами, здесь преобладают сухожильные рецепторы Гольджи. Функцию афферентной системы, контролирующей ее сокращения по принципу обратной связи, выполняют механорецепторы легких и воздухоносных путей.
ДЫХАНИЕ В ИЗМЕНЕННЫХ УСЛОВИЯХ
В различных условиях среды обитания системы нейрогуморальной регуляции дыхания и кровообращения функционируют в тесном взаимодействии как единая кардиореспираторная система. Особенно четко это проявляется при интенсивной физической нагрузке и в условиях гипоксии — недостаточном снабжении организма кислородом. В процессе жизнедеятельности в организме возникают различные виды гипоксии, имеющие эндогенную и экзогенную природу.
Дыхание при физической нагрузке.
Во время выполнения физической работы мышцам необходимо большое количество кислорода. Потребление 02 и продукции СО2 возрастают при физической нагрузке в среднем в 15—20 раз Обеспечение организма кислородом достигается сочетанным усилением функции дыхания и кровообращения. Уже в начале мышечной работы вентиляция легких быстро увеличивается. В возникновении гиперпноэ в начале физической работы периферические и центральные хеморецепторы как важнейшие чувствительные структуры дыхательного центра еще не участвуют. Уровень вентиляции в этот период регулируется сигналами, поступающими к дыхательному центру главным образом из гипоталамуса, лимбической системы и двигательной зоны коры большого мозга, а также раздражением проприорецепторов работающих мышц. По мере продолжения работы к нейрогенным стимулам присоединяются гуморальные воздействия, вызывающие дополнительный прирост вентиляции. При тяжелой физической работе на уровень вентиляции оказывают влияние также повышение температуры, артериальная двигательная гипоксия и другие лимитирующие факторы.
Таким образом, наблюдаемые при физической работе изменения дыхания обеспечиваются сложным комплексом нервных и гуморальных механизмов. Однако из-за индивидуально лимитирующих факторов биомеханики дыхания, особенностей экопортрета человека, не всегда удается при выполнении одной и той же нагрузки полностью объяснить точное соответствие вентиляции легких уровню метаболизма в мышцах.
Дыхание при гипоксии.
Гипоксией (кислородной недостаточностью) называется состояние, наступающее в организме при неадекватном снабжении тканей и органов кислородом или при нарушении утилизации в них кислорода в процессе биологического окисления. Исходя из этого достаточно точного определения гипоксии, все гипоксические состояния целесообразно разделить на экзогенные и эндогенные.
Экзогенная гипоксия развивается в результате действия измененных (в сравнении с обычными) факторов внешней среды.
Эндогенная гипоксия возникает при различных физиологических и патологических изменениях в различных функциональных системах организма.
Реакция внешнего дыхания на гипоксию зависит от продолжительности и скорости нарастания гипоксического воздействия, степени потребления кислорода (покой и физическая нагрузка), индивидуальных особенностей организма и совокупности генетически обусловленных свойств и наследственных морфофункциональных признаков (экопортрет коренных жителей высокогорья и популяции различных этнических групп).
Реакция на гипоксию у коренных жителей высокогорья и у горных животных практически отсутствует, и, по мнению многих авторов, у жителей равнин гипоксическая реакция также исчезает после продолжительной (не менее 3—5 лет) их адаптации к условиям высокогорья.
Основными факторами долговременной акклиматизации к условиям высокогорья являются: повышение содержания углекислоты и понижение содержания кислорода в крови на фоне снижения чувствительности периферических хеморецепторов к гипоксии, увеличения плотности капиллляров и относительно высокого уровня утилизации тканями 02 из крови. У горцев также возрастают диффузионная способность легких и кислородная емкость крови за счет роста концентрации гемоглобина. Одним из механизмов, позволяющих горцам в условиях гипоксии повысить отдачу кислорода тканям и сохранить углекислоту, является способность повышенного образования у них метаболита глюкозы — 2,З - дифосфоглицерата. Этот метаболит снижает сродство гемоглобина к кислороду.

Плотность капилляров сердца:
а — сердце жителя равнины, б — сердце горца
Предметом интенсивных физиологических исследований как в эксперименте, так и в различных природно-климатических и производственных условиях является изучение функционального взаимодействия систем регуляции дыхания и кровообращения. Обе системы имеют общие рефлексогенные зоны в сосудах, которые посылают афферентные сигналы к специализированным нейронам основного чувствительного ядра продолговатого мозга — ядра одиночного пучка. Здесь же в непосредственной близости находятся дорсальное ядро дыхательного центра и сосудодвигательный центр. Особо следует отметить, что легкие являются единственным органом, куда поступает весь минутный объем крови. Это обеспечивает не только газотранспортную функцию, но и роль своеобразного фильтра, который определяет состав биологически активных веществ в крови и их метаболизм.
Дыханиепри высоком атмосферном давлении.
Вероятность возникновения кессонной болезни может быть значительно снижена при дыхании специальными газовыми смесями, например гелиево-кислородной. Гелий почти нерастворим в крови, он быстрее диффундирует из тканей.
Теория по нормальной физиологии. Тема: Регуляция дыхания. Дыхательный центр, опыты Фредерика и Холдена, действие карбогена, хеморецепторы, механорецепторы..
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Регуляция дыхания осуществляется ЦНС.
Спокойное дыхание взрослого человека характеризуется последовательной сменой актов вдоха и выдоха с частотой 14-16 дыханий в минуту.
Чтобы произошел вдох, необходимо сокращение дыхательных мышц.
Импульсы к ним поступают от мотонейронов передних рогов спинного мозга.
К диафрагме – от III – IV шейных сегментов, к межреберным мышцам – от грудных сегментов спинного мозга.
Мотонейроны получают импульсы от дыхательного центра , расположенного в продолговатом мозге.
Если перерезать спинной мозг под продолговатым мозгом , то дыхание – прекращается .
Дыхательный центр
Был открыт в 1812 г. Легаллуа и в 1842 г. Флурансом, которые своими опытами доказали его локализацию в продолговатом мозге .
Н.А.Миславский в 1885 г. уточнил местоположение дыхательного центра – в области РФ (ретикулярной формации) продолговатого мозга , т.к. перерезка между грудными и шейными сегментами спинного мозга – сохраняет диафрагмальное дыхание:
- продолговатым и спинным – полностью прекращает дыхание ,
- выше продолговатого мозга – дыхание сохраняется .
Н.А.Миславским также было показано, что дыхательный центр состоит из двух отделов:
- инспираторного (отвечает на вдох),
- экспираторного (отвечает за выдох).
Они находятся в реципрокных (то есть в противоположных) отношениях.
В настоящее время установлено:
- в РФ продолговатого мозга находятся инспираторные и экспираторные нейроны ,
- эти нейроны рассеяны диффузно , среди других нейронов РФ и относительно друг друга,
- инспираторных нейронов примерно вдвое больше , чем экспираторных.
В продолговатом мозге есть еще два скопления дыхательных нейронов – дорсальные и вентральные дыхательные ядра.
Дорсальное ядро содержит, в основном, инспираторные нейроны, контролирующие сокращение диафрагмы .
В вентральном ядре содержатся как инспираторные , так и экспираторные нейроны, которые включаются при форсированном дыхании.
Все дыхательные нейроны делят на 6 групп:
- ранние инспираторные – активны в начале фазы вдоха (инспирации),
- поздние инспираторные – активны в конце вдоха,
- полные инспираторные – активны в течение всего вдоха,
- постинспираторные – максимальный разряд в начале выдоха,
- экспираторные – активны во вторую фазу выдоха,
- преинспираторные – активны перед вдохом, они включают активную экспирацию (выдох.
Значение дыхательных нейронов:
- Преинспираторные и ранние инспираторные влияют на момент начала вдоха.
- Инспираторные нейроны ДЦ генерируют дыхательный ритм (частоту и глубину дыхания), иннервируют мотонейроны.
- Постинспираторные нейроны контролируют процесс пассивного выдоха.
- Экспираторные нейроны отвечают за активный выдох, так как иннервируют мотонейроны внутренних межреберных мышц и мышц передней брюшной стенки.
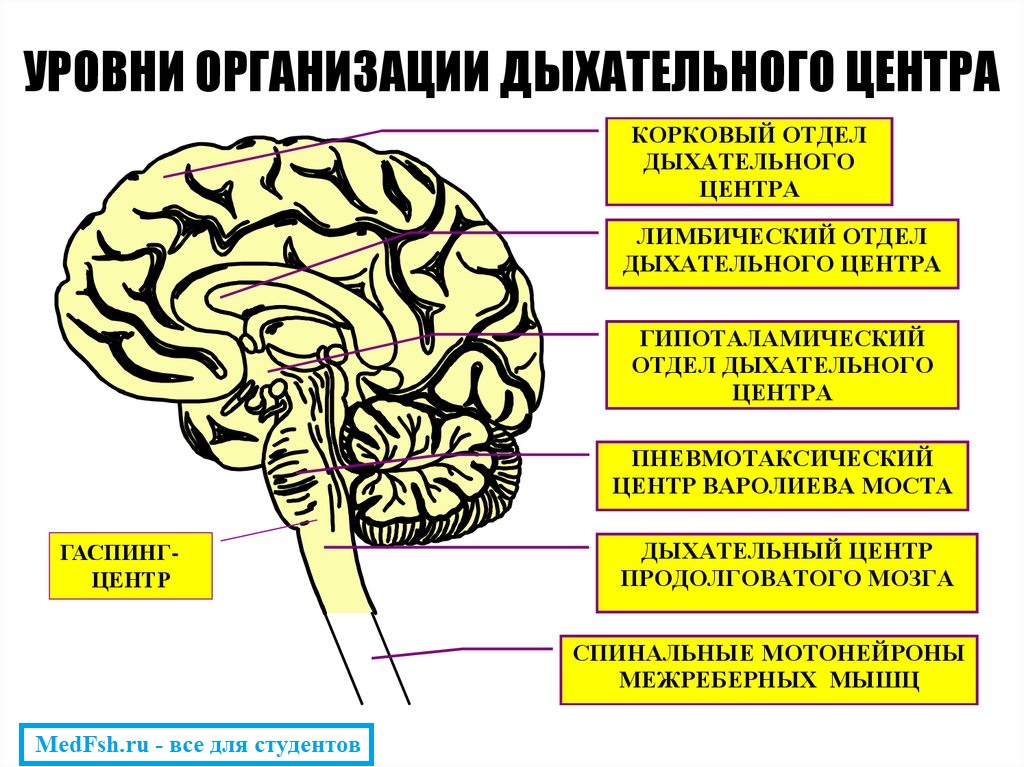
Дыхательный центр – это совокупность нервных клеток, расположенных в различных отделах ЦНС и принимающих участие в регуляции дыхания .
У двух собак, находящихся под наркозом, перекрестно соединяли сонные артерии и яремные вены, латерально же расположенные сосуды пережимали. При этом голова 1-й собаки снабжались кровью из туловища 2-й и наоборот.
У первой собаки кратковременно пережимали трахею, и у нее в крови уменьшалось содержание кислорода (гипоксемия) и увеличивалось содержание углекислого газа (гиперкапния). Эта кровь поступала в голову 2-й собаки, и у нее наступала одышка (диспноэ).
В результате у нее в крови увеличивалось содержание кислорода (гипероксимия) и уменьшалось содержание углекислого газа (гипокапния), и эта кровь поступала в голову 1-й собаки, и у нее наступало апноэ – остановка дыхания. (Нормальное дыхание — эйпноэ).
На состояние ДЦ влияет газовый состав крови:
При увеличении в крови напряжения углекислого газа и уменьшении кислорода, ДЦ – возбуждается и, наоборот, уменьшается , если в крови уменьшается напряжение углекислого газа и увеличивается напряжение кислорода.
При дыхании в герметичной камере в воздухе увеличивалось содержание CO2 и уменьшалось O2 – наступала одышка . Когда CO2 поглощается натронной известью, одышка наступала намного позже, хотя содержание O2 в воздухе значительно снижалось .
Одышка наступает:
- При снижении O2 в атмосферном воздухе с 20,94% до 12%, т.е. на 9%.
- При повышении содержания CO2 в альвеолярном воздухе на 0,17% вентиляция удваивается.
Главный активатор ДЦ – CO2
Действие карбогена
Карбоген – газовая смесь, состоящая из 96% — O2, 4% — CO2.
В сравнении с воздухом в карбогене в 4,8 раза больше O2 и в 130 раз – CO2.
Карбоген применяют при расстройствах дыхания.
Эффект карбогена связан с эффектами содержащегося в нем CO2:
- стимуляция ДЦ,
- расширение бронхов и кровеносных сосудов,
- сдвиг кривой диссоциации HbO2 вправо –> увеличение диффузии O2 из крови в ткани.
Карбоген применяется в медицинских целях для лечения горной болезни, отравления угарным газом, глаукомы, стресса, при восстановлении слуха после воздействия шума и в ряде случаев для улучшения кровоснабжения опухолей при химио- и лекарственной терапии.
В 1911 г. Винтерштейн доказал, что возбудителем ДЦ также являются ионы Н .
Ацидоз – усиливает легочную вентиляцию.
Он показал, что возбуждают ДЦ нелетучие кислоты – молочная, никотиновая и другие.
Самым сильным стимулятором дыхания являются:
- pCO2 (гиперкапния),
- pH (ацидоз),
- pO2 (гипоксемия).
Механизм действия гуморальных факторов (CO2, O2, H) на ДЦ :
- Через хеморецепторы (периферические) сосудистых рефлексогенных зон.
- Через хеморецепторы, находящиеся в продолговатом мозге (медуллярные).
- расположены в каротидных и в аортальных тельцах,
- реагируют на (в артериальной крови):
- увеличение pCO2,
- уменьшение pO2,
- увеличение H (от есть уменьшение pH).
Хеморецепторы возбуждаются постоянно CO2 и O2 , растворенными в крови, а также H, т.к. порог для pCO2 равен 20-30 мм.рт.ст. В норме pCO2 = 40 мм.рт.ст.
Порог для pO2 равен 130-140 мм.рт.ст. В норме pO2 = 100 мм.рт.ст. Одышка же наступает при pO2 ниже 50-60 мм.рт.ст.
Таким образом, хеморецепторы постоянно посылают импульсы в ДЦ, возбуждая инспираторные нейроны, причем большую роль играют хеморецепторы каротидного синуса.
- Находятся на вентролатеральной поверхности продолговатого мозга .
- Реагируют только на H и изменение напряжения CO2.
- Эти рецепторы возбуждаются позднее, поскольку требуется время для проникновения CO2 через гематоэнцефалический барьер.
- Импульсы, поступающие с медуллярных хеморецепторов в ДЦ, увеличивают прирост вентиляции на 60-80% .
Находятся в:
- легких,
- дыхательных путях,
- дыхательных мышцах (проприорецепторы)
Это барорецепторы рефлексогенных зон.
Механорецепторы легких (РРЛ)
Рецепторы растяжения легких (РРЛ) – являются наиболее значимым среди всех механорецепторов.
В 1868 г. Геринг и Брейер доказали наличие в легких рецепторов, которые возбуждаются при их растяжении, то есть при вдохе. Они являются чувствительными окончаниями блуждающих нервов, которые направляют свои импульсы в ДЦ.
Геринг и Брейтер раздували легкие и наблюдали прекражение вдоха (инспираторно-тормозной рефлекс).
Таким образом, этот рефлекс способствует смене вдоха на выдох. Он называется рефлексом Геринга-Брейера и является рефлексом саморегуляции дыхания.
При перерезке блуждающих нервов, дыхание становится редким и глубоким, альвеолы расширяются до максимального предела, т.к. вдох не тормозится. В этом случае, смене вдоха на выдох будет способствовать пневмотаксический центр (ПТЦ).
В настоящее время известно, что в легких существует 3 разновидности механорецепторов :
- РРЛ медленноадаптирующиеся,
- РРЛ быстроадаптирующиеся или ирритантные,
- Юкстаальвеолярные рецепторы капилляров.
РРЛ медленноадаптирующиеся:
- Они расположены в ГМК (гладкомышечных клетках) дыхательных путей.
- Возбуждаются при вдохе.
- С них осуществляется рефлекс Геринга-Брейера.
Ирритантные рецепторы (быстроадаптирующиеся):
- Расположены в слизистой дыхательных путей.
- Реагируют на механические и химические стимулы.
- Быстро адаптирующиеся.
- Длительное раздражение этих рецепторов приводит к хроническому бронхиту.
Физиологическое значение при вдыхании токсических веществ: Сужение бронхов -> вентиляция альвеол -> поступление этих веществ в альвеолы и кровь.
- расположены в паренхиме легких, в альвеолярных перегородках, прилегающих к капиллярам,
- стимулируются, главным образом, растяжением легочных сосудов,
- быстро реагируют на введение химических веществ в легочные сосуды,
- стимуляция может вызвать апноэ, затем учащение дыхания, уменьшение давления, брадикардию и бронхоспазм.
Возбуждение механорецепторов верхних ДП вызывает возникновение дыхательных рефлексов.
Защитные дыхательные рефлексы:
- Чихание – с рецепторов слизистой носа.
- Кашель – с ирритантных рецепторов слизистой гортани, трахеи, бронхов.
- Рефлекс ныряльщиков – остановка дыхания при действии воды на носовые ходы.
- Остановка дыхания во время акта глотания .
- Рефлексогенная задержка дыхания – сужение голосовой щели, бронхоконстрикция при вдыхании дыма, газов, едких веществ.
Механорецепторы межреберных мышц и диафрагмы (проприорецепторы) – они осуществляют обратную связь дыхательных мышц с ДЦ.
Барорецепторы рефлексогенных зон – возбуждаются они при увеличении АД и дыхание при этом угнетается. При уменьшении АД – дыхание усиливается.
Ритмическая смена дыхательных фаз
Хеморецепторы возбуждаются постоянно и постоянно посылают импульсы в продолговатый мозг, возбуждая α-инспираторные нейроны. Они возбуждаются и посылают импульсы к мотонейронам спинного мозга.
Мотонейроны возбуждаются и посылают импульсы к мышцам. Они сокращаются и наступает вдох .
При вдохе объем легких увеличивается и возбуждаются РРЛ, которые посылают возбуждающие импульсы по чувствительным волокнам блуждающего нерва к β-инспираторным нейронам. В результате суммации импульсов от α-инспираторных нейронов и рецепторов растяжения легких достигается порог и возбуждаются и β-инспираторные нейроны благодаря влиянию вышележащих отделов ЦНС.
Инспираторно-тормозные нейроны посылают тормозные импульсы к α-инспираторным нейронам. В результате α-инспираторные нейроны тормозятся и не посылают импульсы к мотонейронам. Мышцы расслабляются, происходит выдох .
К β-инспираторным нейронам не поступают импульсы, и они тормозятся (не возбуждаются).
β-инспираторные нейроны не возбуждают инспираторно-тормозные нейроны и поэтому они не посылают импульсы к α-инспираторным нейронам. α-инспираторные нейроны вновь возбуждаются импульсами от хеморецепторов и наступает вдох .
Читайте также:
