
Электрические явления в нервной ткани
При определенных условиях в нервах и мышцах возникают электрические токи. Различают: 1) токи, или потенциалы, покоя и повреждения и 2) токи, или потенциал, действия – акционные точки.
Токи покоя
При отсутствии раздражения и, следовательно, возбуждения, существует разность потенциалов между наружной поверхностью цитоплазмы живой клетки (мембраной) и её внутренним содержимым.
Поверхностный слой цитоплазмы живой клетки, находиться в состоянии покоя, имеет положительный электрический заряд, а цитоплазма внутри клетки заряжена электроотрицательно. Поэтому, введя в клетку микроэлектрод и приложив другой электрод к поверхности клетки, можно отвести ток к регистрирующему прибору и измерить потенциал покоя, мембранный или клеточный потенциал.
Токи покоя, или клеточные потенциалы, обнаруживаются в неповрежденных живых клетках. Современными методами отведения электрического тока и его регистрации установлено, что токи покоя отдельных клеток животных, например гигантских нервных волокон кальмара, — 50-60 мв (милливольт), миелиновых нервных волокон амфибий – 70 мв, теплокровных – 90 мв, большинства тел нейронов – 80-100 мв, поперечнополосатых мышц амфибий – 80 мв, эпителия амфибий – 20 мв, волокон сердечной мышцы животных с постоянной температурой тела – 95 мв.
Токи повреждения
При повреждении живой ткани открывается доступ к внутреннему содержимому клеток, которое заряжено электроотрицательно. Поэтому если приложить электрод к неповрежденному участку ткани, находящемуся в состоянии покоя и, следовательно, заряженному электроположительно, а другой электрод приложить к поврежденному или перерожденному участку, заряженному электроотрицательно, то при соединении обоих электродов проводником тока с регистрирующим прибором можно измерить разность потенциалов.
Токи повреждения можно обнаружить на нервно-мышечном препарате. Если разрезать мышцу или любым способом повредить ее в одном участке (прижечь, раздавить и т. д.) и затем, приложив неполяризующиеся электроды, соединить срез мышцы или участок повреждения с целой поверхностью мышцы, не подвергающейся раздражению, то от целой поверхности к участку разреза или повреждения направится электрический ток. Сильно поврежденный участок всегда электроотрицателен по отношению к электроположительному неповрежденному участку. Следовательно, биоэлектрический ток повреждения, или альтерационный ток, возникает на границе целой и поврежденной частей ткани. Он обнаруживается и на некотором расстоянии от места повреждения, уменьшаясь с увеличением расстояния от места повреждения. Поэтому по месту происхождения его называют также демаркационным током. Однако ток повреждения появляется только тогда, когда отрицательно заряженный поврежденный участок соединяется проводником тока с положительно заряженным целым участком.
Величина тока повреждения меньше, чем клеточного потенциала, так как межклеточная и внеклеточная жидкости производят шунтирование (укорочение) при его измерении.
Разность потенциалов между целым и поврежденным участками скелетных мышц лягушки сразу же после повреждения равна 25-50 мв, наибольшая разность доходит до 80. В скелетных мышцах кролика токи повреждения равны 40 мв. В гладких мышцах токи повреждения меньше и быстрее уменьшаются до нуля. Разность потенциалов между целым и поврежденным участками седалищного нерва лягушки достигает 20-30 мв, седалищного нерва лошади — 6-16 мв, а седалищного нерва человека и обезьяны — 6-7 мв. Следовательно, токи повреждения нервов меньше, чем токи повреждения мышц. Особенно малы токи повреждения нервов человека и высших животных. Постепенно разность потенциалов уменьшается.
Когда нерв второго нервно-мышечного препарата набрасывается на первый нервно-мышечный препарат таким образом, чтобы он одновременно коснулся неповрежденного поврежденного участков мышцы, электрическая цепь замыкается. При этом мышца второго препарата сокращается в результате раздражения нерва током, возникающим в момент его замыкания между положительно заряженным нормальным участком мышцы первого препарата и отрицательно заряженным поврежденным ее участком.
Токи действия
Акционные токи, или токи (потенциалы) действия, возникают в раздражаемых тканях. Их появление характеризует возбуждение.
Для обнаружения тока действия нужно приложить один электрод к неповрежденному участку живой ткани, который находится в состоящих покоя и поэтому заряжен электроположительно, а другой — к раздражаемому участку и присоединить оба электрода к регистрирующему прибору. Раздражаемый участок при приложении к нему раздражителя немедленно или через несколько сотых миллисекунды становится электроотрицательным. Это падение потенциала’ доходит до максимума, а затем данный участок снова становится электроположительным. Различают однофазные и двухфазные токи действия.
При прикладывании одного из неполяризующихся электродов к неповрежденному участку нерва или мышцы, а другого — к поврежденному участку возникает электрический ток. Этот ток регистрируется при включении в цепь гальванометра или другого более чувствительною прибора.
Стрелка гальванометра отклонится в одну сторону вследствие электроотрицательного заряда поврежденного участка и электроположительного заряда нормального участка. Но если нанести раздражение по соседству с нормальным участком, то волна возбуждения, которая возникнет в месте раздражения, на некоторое время превратит заряд под нормальным участком в электроотрицательный. В этот момент тока не будет, так как под обоими электродами заряд станет электроотрицательным и стрелка гальванометра станет на нулевую линию. Когда волна возбуждения продвинется дальше и будет находиться между электродами, нормальный участок снова станет электроположительным и стрелка гальванометра снова отклонится в ту же сторону.
Когда волна возбуждения дойдет до поврежденного участка, положение стрелки гальванометра не изменится, так как под вторым электродом в месте повреждения уже имеется электроотрицательный заряд. Таким образом, стрелка гальванометра движется только в одну сторону, а запись этого движения регистрирует однофазный, или монофазный ток действия.
Это явление Дюбуа-Реймон назвал отрицательным колебанием тока покоя, которое возникает каждый раз, когда волна возбуждения проходит под электродом, расположенным на неповрежденном участке ткани.
Однофазный ток действия можно зарегистрировать также при раздражении ткани во время отведения клеточного потенциала. При внутриклеточном введении одного электрода и приложении другого электрода к поверхности клетки установлено, что клеточный потенциал не только исчезает в течение тысячных долей секунды, но что в течение этого промежутка времени амплитуда тока действия на 30-50 мв больше клеточного потенциала. Это объясняется тем, что в пункте прохождения волны возбуждения наружная поверхность мембраны становится электроотрицательной, а внутренняя — электроположительной. Продолжительность тока действия в нервных волокнах и клетках скелетных мышц составляет 0,1-5 мс.
Однофазный ток действия представляет собой местный электроотрицательный потенциал, мгновенно возникающий при очень слабых, подпороговых одиночных раздражениях и ограниченный несколькими миллиметрами от пункта раздражения. Величина местного низковольтного электроотрицательного заряда убывает по мере удаления от пункта раздражения. Чем больше подпороговое раздражение, тем больше возникающее возбуждение и амплитуда или отклонение однофазного тока действия.
Следовательно, местное возбуждение или местный сдвиг обмена веществ отличается градуальностью — возрастанием возбуждения по мере увеличения силы раздражения. Местное возбуждение наступает сразу после раздражения, т. е. без латентного периода. В участке слабого местного нераспространяющегося возбуждения возбудимость сохраняется, рефрактерность отсутствует. Если бы при местном возбуждении наблюдалась рефрактерность, то оно не могло бы перейти в пороговое, распространяющееся возбуждение.
Местный однофазный ток действия предшествует распространяющейся волне возбуждения и подготавливает ее. При одиночном пороговом раздражении волна возбуждения возрастает, достигает критического уровня и превращается из местной в распространяющуюся. Поэтому распространяющееся возбуждение наступает после латентного периода, продолжительность которого равна длительности местного нераспространяющегося возбуждения. В отличие от местного возбуждения распространяющееся возбуждение не градуально и сопровождается фазами изменения возбудимости. Потенциал действия, возникший в раздражаемом участке мембраны, вызывает возбуждение соседних участков мембраны и таким образом возбуждение распространяется по нервным и мышечным волокнам.
О прохождении волны возбуждения по возбудимой ткани судят прежде всего по токам действия. Однофазное колебание тока действия распространяющейся волны возбуждения в отдельных нервных и мышечных клетках состоит из нескольких составляющих: 1) высоковольтного потенциала, пика, 2) отрицательного следового потенциала и 3) положительного следового потенциала.
По сравнению со следовыми потенциалами пик имеет наибольшую высоту и наименьшую продолжительность. Пик имеет приблизительно одинаковую высоту на всем протяжении возбудимых тканей животных с постоянной температурой тела. Это указывает на то, что по мере распространения возбуждения вольтаж потенциала не падает. Этот факт свидетельствует, что химические вещества, необходимые для возникновения возбуждения, равномерно распределены по возбудимой ткани и что распространение возбуждения происходит самостоятельно, автоматически. Раздражение при достаточной интенсивности вызывает возбуждение только в том участке, который раздражается, а дальше возбуждение движется по ткани, вовлекая в сдвиг обмена веществ вещества, имеющиеся в возбудимой ткани.
Пик играет ведущую роль в распространении возбуждения. Высота пика различна у нервных волокон разных групп, имеющих неодинаковый диаметр, и изменяется в зависимости от физиологического состояния волокна и скорости проведения возбуждения. Чем толще нервное миелиновое волокно, тем меньше длительность пика. По мышечному волокну человека возбуждение проходит за 4-5 м/с.
В волокнах скелетных мышц однофазные токи действия различаются по амплитуде и по продолжительности. В волокнах сердечной мышцы они отличаются большей продолжительностью пика, который после начального крутого снижения начинает снижаться постепенно, а затем снова круто падает.
Токи действия в гигантских нервных волокнах кальмара достигают 90-110 мв, в миелиновых нервных волокнах амфибий 110 мв, в волокнах поперечнополосатых мышц амфибий — 120 мв, а в волокнах сердечной мышцы животных с постоянной темпера турой тела — 135 мв. Чем больше частота тока действия, тем меньше его вольтаж. Гладкие мышечные волокна отличаются тем, что их токи действия не превосходят мембранных потенциалов (не больше 70-80 мв).
Условия, улучшающие обмен веществ (повышение темпера туры, отдых), уменьшают продолжительность волны возбуждения, а условия, ухудшающие обмен веществ (утомление), увеличивают ее. С понижением температуры амплитуда пика уменьшается, а продолжительность его возрастает.
При температуре тела продолжительность подъема высоковольтного потенциала нерва составляет примерно 1/3 а продолжительность снижения — 2/3 его общей продолжительности. Предполагается, что крутой подъем пика обусловлен быстрым движением ионов натрия внутрь клетки.
Отрицательный следовой потенциал соответствует остаточным сдвигам распределения ионов калия и натрия. В отличие от пика он неустойчив и изменяется от условий среды. Он достигает 0,05 потенциала пика. Положительный низковольтный следовой потенциал также связан с перемещением ионов и равен примерно 0,002 потенциала пика.
Тепло и отдых уменьшают, а охлаждение и истощение увеличивают продолжительность следовой электроотрицательности. В нерве, лишенном кислорода, при действии наркоза и солей калия следовая низковольтная электроотрицательность отсутствует, а начальная высоковольтная электроотрицательность сохраняется, и при известных концентрациях наркоза даже не наступает заметных изменений её величины. Это указывает на то, что пик может возникать и при отсутствии кислорода, а следовая электроотрицательность связана с восстановительными процессами, которые совершаются в ткани вслед за возбуждением, и для её появления необходим кислород. Нарушение обмена веществ, утомление увеличивают продолжительность следовых потенциалов. Чем больше лабильность, тем меньше выражены следовые потенциалы. В высоколабильных мякотных волокнах человека и высших животных они почти не проявляются.
Двухфазный ток действия
В естественных условиях одиночные волны возбуждения встречаются чрезвычайно редко; В организме в нервах и мышцах передаются серии волн возбуждения, взаимно влияющих друг на друга. Возбуждение, которое достигает порога или несколько превышает его, носит ритмический характер. В других случаях возбуждение в естественных условиях является градуальным, длительным, стойким, переходящим в торможение. В последнем случае при чрезмерно большой силе или частоте раздражения, превышающей меру лабильности, полностью угнетается высоковольтная электрическая активность.
Волновое распространяющееся возбуждение возникает из неволнового, градуального. Такая же эволюция возбуждения происходила в филогенезе. Представление о возникновении волнового возбуждения дает следующий простой опыт.
После приложения обоих неполяризующихся электродов к нормальным целым участкам возбудимой ткани и включения этой ткани в цепь гальванометра его стрелка останется на нуле, так как все участки нормальной ткани, находящейся в состоянии покоя, имеют одинаковый заряд. Но когда ткань раздражают у одного из электродов, то возникает возбуждение, и заряд ткани под этим электродом становится отрицательным. При этом стрелка гальванометра отклоняется в одну сторону. Передвигаясь далее, волна возбуждения оказывается между электродами, и в это время заряды под электродами вновь становятся одинаковыми и стрелка гальванометра возвращается к нулю. Но когда волна возбуждения доходит до второго электрода, то гальванометр опять отмечает разность потенциалов, так как под вторым электродом заряд становится отрицательным, а в то же самое время под первым электродом заряд положительный, так как под ним нег возбуждения. Теперь стрелка гальванометра отклоняется в противоположную сторону, а затем, после прекращения возбуждения, возвращается к нулю. Следовательно, когда по ткани проходит волна возбуждения, то стрелка гальванометра отклоняется сначала в одну сторону, потом в другую. Запись этого движения стрелки гальванометра регистрирует двухфазный ток действия.
Форма регистрации токов, или потенциалов, действия зависит от места возникновения возбуждения, от используемых приборов и от расположения электродов. В настоящее время их записывают посредством катодных осциллографов, не имеющих инерции, в отличие от гальванометров. Современные катодные осциллографы регистрируют биопотенциалы в миллионные доли вольта, продолжающиеся одну стомиллионную или даже миллиардную долю секунды. Эти токи отличаются сложностью, и их нередко расшифровывают при помощи электронных устройств.
Биопотенциалы каждой возбудимой ткани, например сердца, имеют характерный вид и изменяются в зависимости от ее функционального состояния. Записываются биопотенциалы кожи, эпителия, соединительной ткани, рецепторов, сетчатки глаза, скелетных мышц, пищеварительных желез, гладкой мускулатуры пищеварительного канала и других органов, разных отделов центральной нервной системы и даже одного нейрона. Первые исследования биопотенциалов больших полушарий головного мозга провели В. Я. Данилевский, Р. Кетон (1875), продолговатого мозга — И. М. Сеченов (1882).
Н. В. Введенский (1883, 1884) соединил мышцу проводами с телефонной трубкой и установил важнейшие законы, по которым протекает в ней возбуждение. Он же впервые применил телефон для изучения явлений возбуждения в нерве.
Выше мы уже показывали, что проведение возбуждения в нервных и мышечных волокнах осуществляется с помощью электрических импульсов, распространяющихся по поверхностной мембране. Передача же возбуждения с нерва на мышцу основана на другом механизме. Она осуществляется в результате выделения нервными окончаниями высокоактивных химических соединений - медиаторов нервного импульса. В синапсах скелетных мышц таким медиатором является ацетилхолин (АХ).
В нервно-мышечном синапсе выделяют три основных структурных элемента - пресинаптическая мембрана на нерве,постсинаптическая мембрана на мышце, между ними - синаптическая щель. Форма синапса может быть разнообразной. В состоянии покоя АХ содержится в так называемых синаптических пузырьках внутри концевой пластинки нервного волокна. От синаптической щели цитоплазма волокна с плавающими в ней синаптическими пузырьками отделена пресинаптической мембраной. При деполяризации пресинаптической мембраны меняется ее заряд и проницаемость, пузырьки подходят близко к мембране и изливаются в синаптическую щель, ширина которой достигает 200-1000 ангстрем. Медиатор начинает диффундировать через щель к постсинаптической мембране.
Постсинаптическая мембрана не электрогенна, но обладает высокой чувствительностью к медиатору за счет наличия в ней так называемых холинорецепторов - биохимических групп, способных избирательно реагировать с АХ. Последний достигает постсинаптической мембраны через 0,2-0,5 мсек. (так называемая "синаптическая задержка") и, взаимодействуя с холинорецепторами, вызывает изменение проницаемости мембраны для Na, что приводит к деполяризации постсинаптической мембраны и генерации на ней волну деполяризации, которая носит название возбуждающего постсинаптического потенциала, (ВПСП), величина которого превышает Ек соседних, электрогенных участков мембраны мышечного волокна. В результате в них возникает ПД (потенциал действия), который распространяется по всей поверхности мышечного волокна, вызывая затем его сокращение, инициируя процесс т.н. электромеханического сопряжения (Каплинг). Медиатор в синаптической щели и на постсинаптической мембране работает очень короткое время, так как разрушается ферментом холинэстеразой, которая готовит синапс к восприятию новой порции медиатора. Показано также, что часть не прореагировавшего АХ может возвращаться в нервное волокно.
При очень частых ритмах раздражения постсинаптические потенциалы могут суммироваться, так как холинэстераза не успевает полностью расщепить выделяющийся в нервных окончаниях АХ. В результате такой суммации постсинаптическая мембрана все более и более деполяризуется. При этом соседние электрогенные участки мышечного волокна приходят в состояние угнетения, сходное с тем, которое развивается при продолжительном действии катода постоянного тока(катодическая депрессия Вериго).
Возбуждение в ткани проявляется в появлении специфической для нее функции (проведение возбуждения нервной тканью, сокращение мышцы, секреция железы) и неспецифических реакциях (генерация потенциала действия, метаболические изменения).
Ток действия (ПД и ПКП) - электрический ток, возникающий в нервных, мышечных и некоторых растительных клетках между их возбуждёнными и соседними покоящимися участками. Обусловлен изменениями ионной проницаемости мембраны и потенциала, которые развиваются в возбуждённом участке. Играет важную роль в распространении потенциала действия вдоль клетки (волокна). Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны.
При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400–500 раз, и градиент нарастает быстро, для ионов К – в 10–15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный. Точные измерения, показали, что амплитуда потенциала действия на 30-50 мВ превышает величину потенциала покоя.
Фазы ПД. ПД состоит из 2 фаз:
1. Фаза деполяризации. Соответствует быстрому изменению мембранного потенциала (деполяризации мембраны) примерно на 110 мВ. Мембранный потенциал изменяется от уровня покоя (около -70мВ) до значения, близкого к равновесному потенциалу – потенциал при котором входящий ток принимает нулевое значение (ЕNa+ (примерно 40 мВ)).
2. Фаза реполяризации. Мембранный потенциал вновь достигает уровня покоя (мембрана реполяризуется), после чего наступает гиперполяризация до значения примерно на 10 мВ меньшего (более отрицательного), чем потенциал покоя, т.е. примерно -80 мВ.
Продолжительность потенциала действия в нервных и в скелетных мышечных волокнах варьирует в пределах 0,1 - 5 мсек., при этом фаза реполяризации всегда продолжительнее фазы деполяризации.
Соотношение фаз потенциала действия и возбудимости. Уровень возбудимости клетки зависит от фазы ПД. В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением. В фазу реполяризации ПД, когда открываются все натриевые каналы и ионы натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза абсолютной рефрактерности. В фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Этому соответствует фаза относительной рефрактерности. Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно в этот момент ее возбудимость повышена. Эта фаза называется фазой супернормальной возбудимости. В момент следовой гиперполяризации МП выше исходного уровня. Она находится в фазе субнормальной возбудимости.
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим.


Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.
Прежде чем приступить к изложению современных представлений о возбуждении в нервной и мышечной тканях, кратко остановимся на исторических аспектах. Как уже говорилось в начале главы, о наличии процесса возбуждения в различных клетках и тканях можно судить по ряду признаков — критериев. Физиологами, изучавшими проблему возбуждения в XIX в., в нервной и мышечной тканях были найдены два весьма показательных и удобных критерия, используемых в клинической и лабораторной практике: появление электрического сигнала в нерве и сокращение мышцы, иннервируемой эти нервом. Классическими опытами, в которых впервые была продемонстрирована связь возбуждения в нервах и мышцах с их электрической активностью, явились эксперименты, проведенные в конце XVIII в. итальянским ученым Л. Гальвани, а затем в начале XIX в. его учеником К. Мат-теучи. Гальвани приготовил из лапки лягушки нервно-мышечный препарат, состоящий из седалищного нерва, включающего в себя большое количество отдельных нервных волокон, и икроножной мышцы, также состоящей из многих отдельных мышечных клеток — мышечных волокон. В середине мышцы делался надрез, в результате чего часть мышечных волокон рассекалась. На поврежденный участок мышцы с помощью стеклянных крючков (стекло было выбрано в качестве изолятора к электрическому току) помещался нерв таким образом, что одна часть касалась поврежденного, а другая — интактного участка мышцы (рис. 2.3, А). В момент накладывания нерва мышца сокращалась. Благодаря результатам
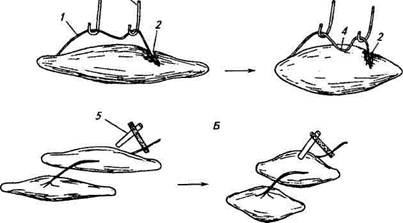 |
| А |
Рис. 2.3. Опыты Гальвани (А) и Маттеучи (Б), доказывающие наличие электрических потенциалов в нервно-мышечном препарате:
1 — нервный ствол; 2—место повреждения мышцы; J-стеклянные крючки; 4— место контакта нерва с интактным участком мышцы; 5 — биметаллический элемент из пластинок меди и цинка для раздражения нерва и мышцы
Результаты экспериментов Маттеучи со вторичным сокращением мышцы оказались в пользу положений Гальвани. Маттеучи в своем опыте использовал два нервно-мышечных препарата (рис. 2.3, Б). Нерв первого препарата соединялся с источником электрического тока, а нерв другого накладывался на мышцу первого. При раздражении нерва электрическим током от внешнего источника тока сокращались мышцы не только первого препарата, но и второго. Таким образом, электрический ток как от внешнего источника, так и возникающий во время возбуждения является эффективным раздражителем для этих тканей. Процесс возбуждения, происходящий в результате раздражения нервов и мышц, может распространяться на значительные расстояния от места возникновения, причем он сопровождается изменением
электрической поляризации ткани. Для объективности следует указать, что прямых измерений электрических токов в нервно-мышечном препарате в описываемых опытах не производилось, тем не менее результаты исследований Гальвани и Маттеучи явились основой для возникновения активно развивающейся по настоящее время науки о механизмах электрических явлений в организмах — электрофизиологии.
Результаты работ физикохимиков Аррениуса, Оствальда, Нернста позволили ученику Дюбуа-Реймона Ю. Бернштейну развить взгляды своего учителя и в начале XX в. сформулировать мембранную теорию биоэлектрических явлений. Согласно концепции Бернштейна, живая клетка окружена мембраной, которая, пропуская одни ионы и задерживая другие, может поляризоваться, т. е. между внутренней и наружной средой клетки возникает электрический потенциал. Необходимо отметить, что к тому времени было известно о существовании мембраны только у растительных клеток. Предположение, что мембрана может быть и у клеток животных, возникло после проведения экспериментов по измерению ионного состава внутри клетки и окружающей среды. Было обнаружено, что внутри клеток преобладают ионы калия, а во внеклеточной среде — ионы натрия. В соответствии с этими данными Бернштейн полагал, что в состоянии покоя клеточная мембрана избирательно проницаема только для ионов калия и эта избирательность утрачивается на время возбуждения, когда мембрана становится в равной мере проницаемой и для других ионов, например для ионов натрия и хлора, при этом потенциал покоя исчезает. Мембранная теория Бернштейна внесла существенный вклад в выяснение механизмов электрических явлений в живых клетках. В частности, она хорошо объясняла, как клетка поддерживает чрезвычайно экономно градиент концентраций ионов. В соответствии с этим получили свое объяснение существование (предсуществование) потенциала покоя, его знак и примерная величина. Вместе с тем в дальнейшем стали накапливаться экспериментальные факты, которые было трудно объяснить с позиций теории Бернштейна. Концепция Бернштейна была существенно видоизменена и переработана в 50-х годах XX в. А. Ходжкиным, А. Хаксли и Б. Катцем. Нужно отметить, что возникновение новой мембранной теории было подготовлено благодаря усовершенствованию методов регистрации электрических потенциалов в живых клетках и экспериментальных методов с одиночными нервными и мышечными клетками, а также нервными волокнами. Кроме того, значительный вклад внесли исследования структуры самой клеточной мембраны. Видимые доказательства существования клеточной мембраны у клеток животных стали возможны только в середине XX в. в связи с появлением электронного микроскопа.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
При подвешивании нервно - мышечного препарата на железную решетку с помощью медного крючка, проходящего через спинной мозг препарата, имело место сокращение мышц лапки каждый раз, когда эта лапка соприкасалась с железной решеткой балкона.
Гальвани считал, что причиной сокращения мышцы в данном случае является электричество, причем, источник этого электричества ученый видел именно в нервно - мышечном препарате лягушки.
Позднее открытия Гальвани были подтверждены в работах Маттеучи (1837).
Однако Маттеучи обнаружил явление вторичного или индуцированного сокращения: при помещении нерва одного нервно-мышечного препарата на мышцу другого препарата и раздражителя нерва этого препарата, Маттеучи наблюдал сокращение мышцы обоих препаратов
На основании этого явления Маттеучи выдвинул предположение об изменении электрических зарядов нервной ткани при ее возбуждении.
Современные представления о природе биоэлектрических явлений в тканях базируются на результатах работ Алана Ходжкина, Эндрью Хаксли, Бернарда Катца. Эти ученые в 40-50 годах нашего века модифицировали и экспериментально обосновали мембранно-ионную теорию Ю. Бернштейна. В настоящее время их взгляды о природе биоэлектрических явлений пользуются всеобщим признанием. Согласно их представлениям, наличие электрических потенциалов в живых клетках обусловлено различной концентрацией ионов Na+, K+, Ca2+ и Cl- внутри и вне клетки, а также различной проницаемостью для них клеточной мембраны. За разработку теории ионного механизма возбуждения эти авторы были удостоены звания лауреатов Нобелевской премии.
Здесь надо рассказывать про потенциал покоя и действия
Понятие о потенциале покоя. Роль ионов К+, Na+, Ca+2, Cl- в происхождении мембранного потенциала. Калий-натриевый насос, его значение. Уравнения Нернста и Гольдмана, расчет величины мембранного потенциала.
Потенциал покоя (ПП) - это разность потенциалов между наружной и внутренней поверхности мембраны в состоянии покоя, т.е. в покое мембрана поляризована.
Происхождение ПП обусловлено:
1. Неравномерным распределением ионов калия и натрия между цитоплазмой и межклеточной жидкостью.
В клетке - калия порядка 400 мкмоль/литр, вне клетки 10, соответственно, натрия в клетке - 50 и 460 - вне клетки - в состоянии покоя.
2. Избирательная проницаемость клеточной мембраны в покое для натрия и калия.
В покое - высокая проницаемость для калия, а для натрия в покое она практически отсутствует небольшая.
В покое за счет процесса облегченной диффузии через неуправляемые медленные калиевые каналы за счет градиента концентрации - калий постоянно выходит из клетки во внеклеточное пространство, это формирует постоянный выходящий калиевый ток. Он является причиной разности потенциалов в покое и обуславливает ПП.
Постоянному выходящему калиевому току противодействует работа калиевая часть калий-натриего насоса, которая обеспечивает постоянное возвратное поступление 2 молекул калия из внешней среды в клетку. В покое скорости этих двух процессов невелики. В реальных условиях в клетке возникает некое равновесное состояние между выходящим калиевым током и входящим калиевым током. Это формирует некий равновесный потенциал /ЕК/, который формирует по существу ту реальную разность потенциалов, которая существует между наружной и внутренней поверхностью клетки, если бы ее создавал один вид ионов.
Его величина, описывается уравнением Нернста:
ie k KK nF RT E ln
где: R- газовая постоянная, Т- абсолютная температура, F- число Фарадея, Ке - концентрация свободных ионов калия в наружном растворе, Кi - их концентрация в цитоплазме, n - валентность, ln - натуральный логарифм.
По этой формуле Нернста можно подсчитать вклад калия в формирование потенциала мембраны в состоянии покоя, а так же - возбуждения. Реально равновесный потенциал для калия в покое равен минус 90 милливольт. Натрий, его равновесный потенциал в покое - плюс 60 милливольт. Хлор, для него равновесный потенциал равен - минус 70.
Гольдман - американский ученый - создал объединительное уравнение, в котором суммировал эти показатели потенциалов для натрия, калия, хлора, только в эти уравнения введено одно добавление - коэффициент проницаемости мембраны для иона, который характеризует скорость диффузии каждого из ионов. Расчет с использованием уравнения Гольдмана показывает, что в состоянии покоя потенциал мембраны составляет минус 70 милливольт. Реальные измерения ПП 70-80 мВ. Таково электрическое состояние мембраны клетки в покое - она поляризована.
Общая физиология. Физиологические основы поведения. Высшая нервная деятельность. Физиологические основы психических функций человека. Физиология целенаправленной деятельности. Приспособление организма к различным условиям существования. Физиологическая кибернетика. Частная физиология. Кровь, лимфа, тканевая жидкость. Кровообращение. Дыхание. Пищеварение. Обмен веществ и энергии. Питание. Центральная нервная система. Методы исследования физиологических функций. Физиология и биофизика возбудимых тканей.
У нас самая большая информационная база в рунете, поэтому Вы всегда можете найти походите запросы
Эта тема принадлежит разделу:
Физиология
Общая физиология. Физиологические основы поведения. Высшая нервная деятельность. Физиологические основы психических функций человека. Физиология целенаправленной деятельности. Приспособление организма к различным условиям существования. Физиологическая кибернетика. Частная физиология. Кровь, лимфа, тканевая жидкость. Кровообращение. Дыхание. Пищеварение. Обмен веществ и энергии. Питание. Центральная нервная система. Методы исследования физиологических функций. Физиология и биофизика возбудимых тканей.
Читайте также:
