
Эволюция нервной системы позвоночных животных достижения этологии
Эволюция нервной системы тесно связана с развитием органов чувств. У всех хордовых животных она построена по типу нервной трубки эктодермального происхождения.
Направления эволюции нервной системы
1. Дифференцировка нервной трубки на головной и спинной мозг.
2. Эволюция головного мозга шла по следующим направлениям:
- увеличение объема и усложнение строения переднего мозга;
- появление и дифференцировка коры переднего мозга;
- увеличение поверхности коры за счет борозд и извилин;
- компактная укладка головного мозга за счет изгибов мозга;
- совершенствование органов чувств.
У ланцетника центральная нервная система очень примитивна и представлена нервной трубкой, которая внешне не делится на головной и спинной мозг, но во внутреннем строении и функциях различия существуют. Расширение передней части нервной трубки представляет собой первый этап в развитии головного мозга хордовых. Повреждение этого отдела ведет к нарушению координации движений. На передней части нервной трубки находится обонятельная ямка Келликера. Весь мозг является светочувствительным, так как по всей нервной трубке расположены многочисленные светочувствительные клетки – глазки Гессе.
У свободноплавающих личинок асцидий, в отличие от живущего в грунте ланцетника, передняя часть нервной трубки содержит хорошо развитый глаз и своеобразно дифференцированный орган равновесия – сложный сенсорный центр. Однако при переходе во взрослое состояние нервная система и органы чувств асцидий подвергаются большей редукции, чем у ланцетника.
В противоположность бесчерепным и оболочникам у позвоночных, как свободноживущих и активно питающихся животных, сформировался сложнейший головной мозг со сложными органами чувств.
Головной мозг всех классов позвоночных состоит из пяти отделов и в процессе эмбриогенеза проходит одинаковые стадии. На переднем конце нервной трубки возникают три расположенные друг за другом вздутия (пузыря) – передний, средний и задний. Передний и задний разделяются поперечным перехватом на два, а самая передняя часть переднего пузыря – продольной бороздой – на правую и левую половины. Так образуются пять вторичных мозговых пузырей, дающих начало пяти отделам головного мозга: конечному (или большим полушариям), промежуточному (вместе с конечным он составляет передний мозг), среднему, заднему (включающему мозжечок) и продолговатому. У млекопитающих в состав заднего мозга входит мост и мозжечок, у всех остальных позвоночных мост в самостоятельный отдел головного мозга не выделяется, поскольку связи между спинным мозгом и головным мозгом у них еще малочисленны. Невроцель (полость нервной трубки) тоже дифференцируется на отдельные сообщающиеся между собой полости – желудочки головного мозга, заполненные спинно-мозговой жидкостью. У высших позвоночных I и II желудочки расположены в переднем мозге, у низших – это одна общая полость, III желудочек расположен в полости промежуточного мозга. Полость среднего мозга называется сильвиевым водопроводом. IV желудочек расположен в продолговатом мозге и называется ромбовидной ямкой, которая продолжается в полость спинного мозга. Передние части переднего мозга образуют парный выступ – обонятельные доли.
У круглоротых головной мозг очень примитивного строения, отделы головного мозга выражены слабо, лежат в одной плоскости. Передний мозг мал, в нем хорошо развиты обонятельные доли. Обонятельные доли, стенки полостей желудочков и дно переднего мозга состоят из серого мозгового вещества. Крыша мозга не содержит нервных клеток и образована эпителиальной тканью. На крыше промежуточного мозга расположены теменной орган (париетальный) и эпифиз (пинеальный), которые выполняют функцию дополнительных светочувствительных органов (рис. 1).
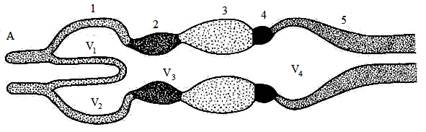
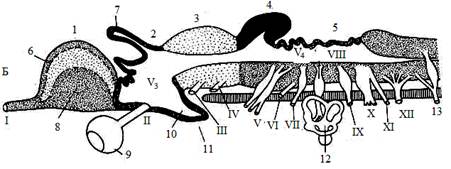
Рис. 1. Схема строения головного мозга позвоночных
А–фронтальный разрез, Б–сагиттальный разрез. 1–передний мозг; продольный срез проходит между парными передними мозговыми пузырями (полушариями), показано внутреннее строение правого полушария: плащ (6) и базальный ганглий (8); 2–промежуточный мозг: эпифиз (7), воронка (10), гипофиз (11); глазные яблоки (9); 3–средний мозг; 4–мозжечок; 5–продолговатый мозг.
V1, V2–боковые желудочки полушарий переднего мозга; V3–третий желудочек; V4–четвертый желудочек. Головные нервы: I–обонятельный,
II–зрительный, III–глазодвигательный, IV–блоковый, V–тройничный,
VI–отводящий, VII–лицевой, VIII–равновесно-слуховой,
IX–языкоглоточный, X–блуждающий, XI–добавочный; XII–подъязычный; 12–слуховой лабиринт, 13–первый спинномозговой нерв.
У рыб головной мозг небольших размеров. У костных рыб он устроен во многом примитивнее, чем у хрящевых: конечный мозг развит слабо и не разделен на полушария, поэтому его полость едина и не образует боковых желудочков. Нервная ткань имеется только в дне конечного мозга – полосатых телах, и в обонятельных долях. Крыша эпителиальная. Основной функцией конечного мозга рыб является обработка обонятельных импульсов, кроме того, он участвует в регуляции двигательной активности и поведения, поскольку связан с промежуточным и средним мозгом. В состав промежуточного мозга входят зрительные бугры (таламусы) и гипоталамус. В таламусы поступает информация от всех органов чувств, там находится центр координации движений. Гипоталамус вместе с гипофизом (нижний придаток промежуточного мозга) образуют гипоталамо-гипофизарную систему, которая регулирует деятельность внутренних органов и эндокринных желез. Теменной орган сохраняется у некоторых рыб как светочувствительный орган. Эпифиз функционирует как железа внутренней секреции. Средний мозг – самый крупный отдел. Основная часть его образует зрительные доли. Сюда поступает информация от органов чувств, происходит ее анализ и синтез, а затем по эфферентным нейронам импульсы направляются в другие отделы головного мозга и в спинной мозг. Поэтому средний мозг (точнее, его крыша) у всех рыб является интегрирующим центром нервной системы. Такой тип головного мозга называют ихтиопсидным. У всех классов позвоночных животных (за исключением млекопитающих) крыша среднего мозга разделена продольной бороздой на два бугра (двухолмие). Мозжечок хорошо развит, он координирует движения, поэтому у быстрых пловцов (акул) достигает больших размеров. В продолговатом мозге сосредоточены центры, управляющие деятельностью кровеносной, пищеварительной, дыхательной систем, опорно-двигательного аппарата. Через продолговатый мозг проходят связи между спинным и головным мозгом, однако, у рыб таких волокон немного. В области дна продолговатого мозга имеется пара гигантских маутнеровских клеток, толстые аксоны которых направляются в спинной мозг и проходят его до конца. Функция маутнеровских клеток – общий контроль за спинномозговыми рефлексами, управляющими последовательными изгибами тела в процессе движения рыбы в толще воды. В области среднего мозга появляется изгиб. От головного мозга рыб отходят 10 пар черепно-мозговых нервов.
Ихтиопсидного типа головной мозг земноводных, по сравнению с рыбами, имеет ряд прогрессивных особенностей. Передний мозг значительно крупнее и более четко дифференцирован. Относительно крупный конечный мозг разделен полной перегородкой, при этом образуются отдельные латеральные желудочки. Крыша и боковые стенки переднего мозга состоят из нервной ткани, однако на поверхности находятся лишь отростки нейронов, а тела клеток лежат ниже. Таким образом, у амфибий формируется первичный мозговой свод – архипаллиум (среди рыб свод мозга имеют лишь двоякодышащие). Развитие архипаллиума, сопровождающееся усилением связей с промежуточным и особенно средним мозгом, приводит к тому, что ассоциативная деятельность, регулирующая поведение, осуществляется не только продолговатым и средним мозгом, но и полушариями переднего мозга. Обонятельные доли слабо отделены от полушарий. Промежуточный мозг имеет небольшие размеры. Сверху на нем расположен эпифиз. От дна промежуточного мозга отходит воронка, к которой прилегает хорошо развитый гипофиз. Средний мозг крупный, особенно его крыша. Средний мозг, как и у рыб, является интегрирующим центром центральной нервной системы. Мозжечок из-за простоты движений амфибий развит слабо и имеет вид небольшого поперечного валика. Продолговатый мозг развит также как у рыб. Изгибы мозга выражены слабо. От головного мозга отходят 10 пар черепно-мозговых нервов.
У рептилий передний мозг является самым крупным отделом. Основная масса серого вещества полушарий сосредоточена в полосатых телах, расположенных в основании конечного мозга. У рептилий они становятся главными координаторами нервной деятельности. Головной мозг, в котором интегрирующим центром являются полосатые тела конечного мозга, относят к зауропсидному типу. Его имеют рептилии и птицы. В полушариях переднего мозга четко дифференцированы обонятельные доли. Прогрессивной особенностью мозга рептилий является появление на боковых поверхностях полушарий отдельных участков серого вещества – коры, которые представляют собой зачаток вторичного мозгового свода, или неопаллиума. Однако строение коры очень примитивно, поэтому ее называют древней корой (архикортекс). На крыше промежуточного мозга сидит теменной орган, который имеет глазоподобное строение (у гаттерии и ящериц). Эпифиз и гипофиз функционируют как железы внутренней секреции. Размеры среднего мозга относительно невелики, поскольку этот отдел не является ведущим. Мозжечок значительно увеличен за счет разрастания серого вещества коры мозжечка. Продолговатый мозг образует резкий изгиб в вертикальной плоскости, который характерен для всех амниот. От головного мозга отходят 11 пар черепно-мозговых нервов.
У птиц, несмотря на большие размеры полушарий, сохраняются черты строения мозга рептилий. Мозговые центры, которые ведают высшей нервной деятельностью, развиваются не в коре полушарий переднего мозга (как у млекопитающих), а в основании полушарий, образуя систему центров полосатых тел, которые являются основным интегрирующим центром нервной системы. Поэтому головной мозг птиц, как и рептилий, относится к зауропсидному типу. Обонятельные доли малы, что связано с плохим обонянием птиц. В среднем мозге выделяются очень крупные зрительные доли. В этом отделе происходит высший анализ сигналов, поступающих от органов зрения (у млекопитающих эту функцию выполняет кора полушарий конечного мозга). Мозжечок развит значительно лучше, чем у рептилий, поэтому птицы способны совершать сложные высоко координированные движения, что особенно важно в полете. Имеется 12 пар черепно-мозговых нервов.
У млекопитающих значительно развиваются все отделы головного мозга. Наибольшего развития достигает конечный мозг, в котором большая часть серого мозгового вещества сосредоточена в коре полушарий, тогда как полосатые тела относительно невелики. Кора больших полушарий мозга формируется путем разрастания нервной ткани стенок боковых желудочков. Образующийся таким образом мозговой свод носит название вторичного свода или неопаллиума; его зачатки появляются у амфибий и более заметны у рептилий и птиц. В коре больших полушарий располагаются центры высшей нервной деятельности, они обеспечивают наиболее сложные поведенческие реакции, которые развиваются в течение жизни, максимально эффективно приспосабливая животных к быстро меняющимся условиям внешней среды. Кроме того, в коре располагаются высшие нервные центры анализаторов. Кора конечного мозга становится ведущим отделом головного мозга млекопитающих, который интегрирует деятельность ЦНС. Мозг такого типа называют маммальным. Усложнение поведения и развитие многочисленных условных рефлексов требует значительной площади коры. У низших млекопитающих кора гладкая, а у высших – образуются борозды и извилины, что существенно увеличивает общую поверхность коры. Положение и количество извилин и борозд является характерным признаком для того, или иного вида. Обонятельные доли развиты хорошо, особенно у тех видов, для которых обоняние является ведущим органом чувств. Полосатые тела выполняют функцию регуляции инстинктивных реакций под контролем коры полушарий. Промежуточный мозг включает зрительные бугры (таламусы), через которые проходит зрительный тракт и где происходит первичная обработка зрительной информации. В крыше расположен эпифиз (эндокринная железа). В дне промежуточного мозга (гипоталамусе) расположены вегетативные центры, участвующие в регуляции процессов метаболизма и терморегуляции. Там же находится воронка, тесно соединенная с эндокринной железой – гипофизом. Средний мозг характеризуется развитием четверохолмия с передними зрительными и задними слуховыми долям, они обеспечивают ориентировочные реакции на световые и слуховые раздражители. У всех прочих классов позвоночных крыша среднего мозга разделена продольной бороздой на два бугра (двухолмие). Задний мозг представлен мозжечком и хорошо выраженным мостом, что связано с более высокой организацией проводящих путей у млекопитающих, значительная их часть проходит через мост. Мозжечок увеличен в размерах, дифференцируется на два полушария и среднюю часть – червь. Имеются три изгиба мозга: теменной – на уровне среднего мозга, затылочный – в месте перехода продолговатого в спинной и мостовой, который направлен дорсально. От головного мозга отходят 12 пар черепно-мозговых нервов.
Нервная система возникла у многоклеточных в связи с необходимостью быстро реагировать на изменения внешней среды. Нервная система хордовых эктодермального происхождения, производной нервной трубки с полостью внутри. Передний отдел нервной трубки увеличивается в размерах, дифференцируется на отделы, преобразуются в головной мозг, который включает регулирующие центры.
Головной мозг позвоночных закладывается из 3-х мозговых пузырей: переднего (передний и промежуточный мозг), среднего (средний) и заднего (задний и продолговатый). Внутри головного и спинного мозга располагается общая полость соответствующая нейроцелям, где циркулирует спинномозговая жидкость. В спинном мозге имеется спинномозговой канал, в головном – желудочки мозга.
Направление эволюции – усиливает роль передних отделов мозга и крыши по сравнению с задним основанием. Наблюдается смена 3-х типов мозга: 1) ихтиопсидный – у рыб и земноводных, высший интегральный центр находится в среднем мозге со стороны дна, изгиб мозга один – теменной. 2) зауропсидный – у птиц и рептилий, высший интегральный центр – полосатые тела в области переднего мозга со стороны дна. Изгибов 2 – теменной и затылочный. 3) маммалийный – у млекопитающих, высший интегральный центр – передний мозг со стороны крыши, изгибов 3 – теменной, затылочный, мозговой.
Головной мозг рыб имеет примитивное строение. В продолговатом мозге находятся центры дыхания, кровообращения, пищеварения. Задний мозг - хорошо развит мозжечок (компактный, цельный). Средний мозг наиболее развит, разделен на 2-е зрительные доли, имеет в крыше слой серого вещества, здесь теменной изгиб. Промежуточный – в нем гипоталамус, синтезирующий нейрогормоны, здесь же расположены эпифиз и гипофиз. Передний мозг развит слабо, не разделен на полушария, крыша тонкая, состоящая из эпителия; хорошо развиты обонятельные доли.
Земноводные – продолговатый как у рыб. Мозжечок редуцирован в связи с примитивным характером движений. Средний мозг – достигает больших размеров, является высшим интегрирующим центром, центр зрения, теменной изгиб. Передний мозг – разделен на полушария, серое в-во дифференцируется на: древнюю кору и зачаток старой коры.
Пресмыкающиеся – продолговатый мозг хорошо развит, образует изгиб в вертикальной полости (затылочный). Мозжечок хорошо развит, компактный, цельная структура. Средний мозг теряет значение ведущего отдела, теменной изгиб. Промежуточный мозг отвечает за вегетативные функции и нейрогуморальные регуляции. Передний мозг разделен на полушария, высший интегральный центр – полосатые тела. Крыша переднего мозга – архикортекс, в виде 2-х групп островков.
Млекопитающие – продолговатый мозг хорошо развит. Мозжечок сильно увеличивается в размерах, имеет 2 полушария. Средний мозг состоит из четыреххолмия. Промежуточный мозг – эпифиз, гипофиз. Передний мозг полностью разделен на 2 полушария, является ведущим отделом. Кора имеет очень сложное строение.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Нервная система возникла у многоклеточных в связи с необходимостью быстро реагировать на изменения внешней среды. Нервная система хордовых эктодермального происхождения, производной нервной трубки с полостью внутри. Передний отдел нервной трубки увеличивается в размерах, дифференцируется на отделы, преобразуются в головной мозг, который включает регулирующие центры.
Головной мозг позвоночных закладывается из 3-х мозговых пузырей: переднего (передний и промежуточный мозг), среднего (средний) и заднего (задний и продолговатый). Внутри головного и спинного мозга располагается общая полость соответствующая нейроцелям, где циркулирует спинномозговая жидкость. В спинном мозге имеется спинномозговой канал, в головном – желудочки мозга.
Направление эволюции – усиливает роль передних отделов мозга и крыши по сравнению с задним основанием. Наблюдается смена 3-х типов мозга: 1) ихтиопсидный – у рыб и земноводных, высший интегральный центр находится в среднем мозге со стороны дна, изгиб мозга один – теменной. 2) зауропсидный – у птиц и рептилий, высший интегральный центр – полосатые тела в области переднего мозга со стороны дна. Изгибов 2 – теменной и затылочный. 3) маммалийный – у млекопитающих, высший интегральный центр – передний мозг со стороны крыши, изгибов 3 – теменной, затылочный, мозговой.
Головной мозг рыб имеет примитивное строение. В продолговатом мозге находятся центры дыхания, кровообращения, пищеварения. Задний мозг - хорошо развит мозжечок (компактный, цельный). Средний мозг наиболее развит, разделен на 2-е зрительные доли, имеет в крыше слой серого вещества, здесь теменной изгиб. Промежуточный – в нем гипоталамус, синтезирующий нейрогормоны, здесь же расположены эпифиз и гипофиз. Передний мозг развит слабо, не разделен на полушария, крыша тонкая, состоящая из эпителия; хорошо развиты обонятельные доли.
Земноводные – продолговатый как у рыб. Мозжечок редуцирован в связи с примитивным характером движений. Средний мозг – достигает больших размеров, является высшим интегрирующим центром, центр зрения, теменной изгиб. Передний мозг – разделен на полушария, серое в-во дифференцируется на: древнюю кору и зачаток старой коры.
Пресмыкающиеся – продолговатый мозг хорошо развит, образует изгиб в вертикальной полости (затылочный). Мозжечок хорошо развит, компактный, цельная стр-ра. Средний мозг теряет значение ведущего отдела, теменной изгиб. Промежуточный мозг отвечает за вегетативные функции и нейрогуморальные регуляции. Передний мозг разделен на полушария, высший интегральный центр – полосатые тела. Крыша переднего мозга – архикортекс, в виде 2-х групп островков.
Млекопитающие – продолговатый мозг хорошо развит. Мозжечок сильно увеличивается в размерах, имеет 2 полушария. Средний мозг состоит из четыреххолмия. Промежуточный мозг – эпифиз, гипофиз. Передний мозг полностью разделен на 2 полушария, является ведущим отделом. Кора имеет очень сложное строение.
Эволюция мочеполовой системы позвоночных.
Выделительная и половая система выполняют разные функции, но рассматриваются в едином комплексе в связи с единством эмбрионального развития и связью со второй полостью тела. В эмбриогенезе закладка мочеполовой системы формируется в области ножки сомита (мезодерма) в непосредственном контакте.
Половые железы располагаются в целоме, продукты диссимиляции тоже выводятся в целом. Наиболее простой способ выведения половых клеток и продуктов диссимиляции во внешнею среду ч/з общий канал, к-ый начинается воронкой в целоме и открывается на покровах.
Принцип выведения продуктов диссимиляции сходен, осуществляется благодаря двум основным принципам: 1) ультрафильтрация жидкости, ч/з полупроницаемую мембрану организмов, выделяя непроходимые белки и другие крупные молекулы; 2) активный транспорт веществ происходит в 2-х противоположных направлениях (с помощью секреции продукты обмена переносятся из внутренней среды в просвет экскреторного органа; при реабсорбции в обратном направлении происходит транспорт глюкозы, воды, аминокислот, анионов и катионов).
Состав мочевыделительной системы: почки, мочеточники, мочевой пузырь, мочеиспускательный канал.
Эволюция почки. В филогенезе почка прошла 3 этапа: предпочка, туловищная, тазовая.
Предпочка закладывается у зародышей, как самостоятельный орган. Функционирует у круглоротых, личинок рыб, земноводных. Находится на переднем конце тела, состоящий из 6-12 нефронов. Нефрон – воронка, открывающаяся в целом и соединяется прямым выделительным канальцем с общим выводным протоком (пронефротический каналец), в к-ый открывается клапан.
Туловищная (первичная почка) функционирует у взрослых рыб и земноводных. Формируется в туловищных сегментах кзаде от предпочки, содержит несколько 100 нефронов. Число нефронов увеличивается за счет почкования. Нефроны вступают в связь с кровеносной системой формирует капсулы почечных клубочков. Капсулы имеют вид двустенных чаш, где располагаются сосудистые клубочки – капсула Шумлянского-Боуэна. Продукты диссимиляции поступают ч/з кровь в нефроны, некоторые нефроны сохраняют связь с целомом. Выделительные канальцы удлиняются, у земноводных появляется первая часть изветого канальца – проксимальный отдел, где осуществляется обратное всасывание в кровь воды, глюкозы и др в-тв.
Тазовая (вторичная почка) у пресмыкающихся, птиц и млекопитающих. Закладывается в тазовом отделе тела, содержит 100-и тысяч и млн нефронов. Нефроны образуются за счет многократного выделения развивающихся нефронов. Нефроны не имеют воронки и теряют связь с целомом, фильтрация только из крови. Каналец нефрона удлиняется, дифференцируется на проксимальный и дистальный участки, м/у которыми у птиц и млекопитающих появляется петля Генли, чем обеспечивается эффективное обратное всасывание в кровь воды, глюкозы, гормонов и т.д. Количество мочи мало, концентрация продуктов диссимиляции велика.
В эмбриогенезе при развития предпочки вдоль тела (от головы к клоаке) закладывается пронефрический канал, по которому продукты диссимиляции из нефронов поступает во внешнюю среду. При развитии первичной почки этот канал либо расщепляется на два, идущих параллельно, либо 2ой канал образуется в продольном утолщении стенки 1ого. Вольфов канал вступает в связь с нефронами первичной почки, Мюллеров – срастается передним концом с 1им из нефронов почки и образует яйцевод.
32. Эволюция иммунной и эндокринной систем. Онтофилогенетическая обусловленность пороков развития.
Эндокринная система – система желез внутренней секреции, топографически не является единым целым, но единофункционально, т.к. обеспечивают гуморальную регуляцию функций организмов. Регуляция осуществляется гормонами.
Гуморальная регуляция возникла раньше нервной, более проста, не требует развития сложных стр-ры.
Эволюция желез. Гипофиз – мозговой придаток, у млекопитающих состоящий из 3-х долей: передней (аденогипофиз), среднего (промежуточный), задней (нейрогипофиз). Гипофиз соединяется с гипоталамусом выростом – воронкой. Доли имеют разное происхождение: передний развивается из выпячивания эктодермального эпителия крыши ротовой полости; задний – из задней части воронки; промежуточная – производная от передней.
У рыб и личинок земноводных имеет передний и промежуточные доли. У взрослых амфибий, переходящих к наземному существованию, впервые появляется задняя доля, регулирующая водный обмен. У пресмыкающихся и млекопитающихся наиболее прогрессивно развивается задняя доля, что связано с интенсификацией воды и строением доли.
Щитовидная железа выделяет гормоном тироксин, регулирующий энергетический обмен, как компактный орган впервые проявляется у рыб. Закладывается м/у 1 и 2 жаберными щелями. У костных рыб состоит из 2-х частей, располагающихся под глодкой, у других позвоночных закладывается м/у 2 и 3 жаберными щелями. У земноводных перемещается в область подъязычной кости, состоит из 2-х долей. У пресмыкающихся – в шейную область. У птиц – у основания бронхов, у млекопитающих – в область шеи, имеет 2е ч. с добавочными дольками и перейшком.
Паращитовидная железа и ультимобранхиальные тельца – мелкие эндокринные образования, развиваются из эпителия глотки в области 3-5 жаберных карманов, регулирует кальциевый обмен. Паращитовидные железы, как самостоятельные появляются у наземных.
Тимус (зобная железа) – располагается под щитовидной только у позвоночных. Развивается как выступ эктодермы эпителия в спинной части жаберных мешков, затем туда врастают лимфоидные образования, происходящих из мезодермы. У рыб зачатки располагаются на всех жаберных мешках, вырабатывают антитела. У амфибий и рептилий – 2-3 пара жаберных мешков, вырабатывает антитела. У птиц – 3-4 пара жаберных мешков, отвечает за тканевую не совместимость. У млекопитающих – из 2 пары жаберных мешков, наиболее развит у молодых особей. У человека с 14 лет начинает обратное развитие.
Пороки: недоразвитие задней доли гипофиза; эктопия аденогипофиза; сохранение кармана Ратхе; эктопия щитовидной железы; крипторхизм.
Дата добавления: 2019-02-12 ; просмотров: 462 ;
Нервная система возникла у многоклеточных в связи с необходимостью быстро реагировать на изменения внешней среды. Нервная система хордовых эктодермального происхождения, производной нервной трубки с полостью внутри. Передний отдел нервной трубки увеличивается в размерах, дифференцируется на отделы, преобразуются в головной мозг, который включает регулирующие центры.
Головной мозг позвоночных закладывается из 3-х мозговых пузырей: переднего (передний и промежуточный мозг), среднего (средний) и заднего (задний и продолговатый). Внутри головного и спинного мозга располагается общая полость соответствующая нейроцелям, где циркулирует спинномозговая жидкость. В спинном мозге имеется спинномозговой канал, в головном – желудочки мозга.
Направление эволюции – усиливает роль передних отделов мозга и крыши по сравнению с задним основанием. Наблюдается смена 3-х типов мозга: 1) ихтиопсидный – у рыб и земноводных, высший интегральный центр находится в среднем мозге со стороны дна, изгиб мозга один – теменной. 2) зауропсидный – у птиц и рептилий, высший интегральный центр – полосатые тела в области переднего мозга со стороны дна. Изгибов 2 – теменной и затылочный. 3) маммалийный – у млекопитающих, высший интегральный центр – передний мозг со стороны крыши, изгибов 3 – теменной, затылочный, мозговой.
Головной мозг рыб имеет примитивное строение. В продолговатом мозге находятся центры дыхания, кровообращения, пищеварения. Задний мозг - хорошо развит мозжечок (компактный, цельный). Средний мозг наиболее развит, разделен на 2-е зрительные доли, имеет в крыше слой серого вещества, здесь теменной изгиб. Промежуточный – в нем гипоталамус, синтезирующий нейрогормоны, здесь же расположены эпифиз и гипофиз. Передний мозг развит слабо, не разделен на полушария, крыша тонкая, состоящая из эпителия; хорошо развиты обонятельные доли.
Земноводные – продолговатый как у рыб. Мозжечок редуцирован в связи с примитивным характером движений. Средний мозг – достигает больших размеров, является высшим интегрирующим центром, центр зрения, теменной изгиб. Передний мозг – разделен на полушария, серое в-во дифференцируется на: древнюю кору и зачаток старой коры.
Пресмыкающиеся – продолговатый мозг хорошо развит, образует изгиб в вертикальной полости (затылочный). Мозжечок хорошо развит, компактный, цельная стр-ра. Средний мозг теряет значение ведущего отдела, теменной изгиб. Промежуточный мозг отвечает за вегетативные функции и нейрогуморальные регуляции. Передний мозг разделен на полушария, высший интегральный центр – полосатые тела. Крыша переднего мозга – архикортекс, в виде 2-х групп островков.
Млекопитающие – продолговатый мозг хорошо развит. Мозжечок сильно увеличивается в размерах, имеет 2 полушария. Средний мозг состоит из четыреххолмия. Промежуточный мозг – эпифиз, гипофиз. Передний мозг полностью разделен на 2 полушария, является ведущим отделом. Кора имеет очень сложное строение.
31. Эволюция мочеполовой системы позвоночных.
Выделительная и половая система выполняют разные функции, но рассматриваются в едином комплексе в связи с единством эмбрионального развития и связью со второй полостью тела. В эмбриогенезе закладка мочеполовой системы формируется в области ножки сомита (мезодерма) в непосредственном контакте.
Половые железы располагаются в целоме, продукты диссимиляции тоже выводятся в целом. Наиболее простой способ выведения половых клеток и продуктов диссимиляции во внешнею среду ч/з общий канал, к-ый начинается воронкой в целоме и открывается на покровах.
Принцип выведения продуктов диссимиляции сходен, осуществляется благодаря двум основным принципам: 1) ультрафильтрация жидкости, ч/з полупроницаемую мембрану организмов, выделяя непроходимые белки и другие крупные молекулы; 2) активный транспорт веществ происходит в 2-х противоположных направлениях (с помощью секреции продукты обмена переносятся из внутренней среды в просвет экскреторного органа; при реабсорбции в обратном направлении происходит транспорт глюкозы, воды, аминокислот, анионов и катионов).
Состав мочевыделительной системы: почки, мочеточники, мочевой пузырь, мочеиспускательный канал.
Эволюция почки. В филогенезе почка прошла 3 этапа: предпочка, туловищная, тазовая.
Предпочка закладывается у зародышей, как самостоятельный орган. Функционирует у круглоротых, личинок рыб, земноводных. Находится на переднем конце тела, состоящий из 6-12 нефронов. Нефрон – воронка, открывающаяся в целом и соединяется прямым выделительным канальцем с общим выводным протоком (пронефротический каналец), в к-ый открывается клапан.
Туловищная (первичная почка) функционирует у взрослых рыб и земноводных. Формируется в туловищных сегментах кзаде от предпочки, содержит несколько 100 нефронов. Число нефронов увеличивается за счет почкования. Нефроны вступают в связь с кровеносной системой формирует капсулы почечных клубочков. Капсулы имеют вид двустенных чаш, где располагаются сосудистые клубочки – капсула Шумлянского-Боуэна. Продукты диссимиляции поступают ч/з кровь в нефроны, некоторые нефроны сохраняют связь с целомом. Выделительные канальцы удлиняются, у земноводных появляется первая часть изветого канальца – проксимальный отдел, где осуществляется обратное всасывание в кровь воды, глюкозы и др в-тв.
Тазовая (вторичная почка) у пресмыкающихся, птиц и млекопитающих. Закладывается в тазовом отделе тела, содержит 100-и тысяч и млн нефронов. Нефроны образуются за счет многократного выделения развивающихся нефронов. Нефроны не имеют воронки и теряют связь с целомом, фильтрация только из крови. Каналец нефрона удлиняется, дифференцируется на проксимальный и дистальный участки, м/у которыми у птиц и млекопитающих появляется петля Генли, чем обеспечивается эффективное обратное всасывание в кровь воды, глюкозы, гормонов и т.д. Количество мочи мало, концентрация продуктов диссимиляции велика.
В эмбриогенезе при развития предпочки вдоль тела (от головы к клоаке) закладывается пронефрический канал, по которому продукты диссимиляции из нефронов поступает во внешнюю среду. При развитии первичной почки этот канал либо расщепляется на два, идущих параллельно, либо 2ой канал образуется в продольном утолщении стенки 1ого. Вольфов канал вступает в связь с нефронами первичной почки, Мюллеров – срастается передним концом с 1им из нефронов почки и образует яйцевод.
Читайте также:
