
Факторы роста нервной ткани в центральной нервной системе
Фактор роста нервов (NGF) – это одна из групп небольших протеиноподобных молекул, которые называются нейротрофины (BDNF — другая). Они отвечают за развитие новых и здоровье старых нейронов.
Нейротрофины способствуют росту, поддержанию и выживанию нейронов и аксонов. Он также помогает восстановить миелиновую оболочку, покрывающую аксоны. Испытания на животных показали, что с уменьшением выработки NGF снижается способность формировать новые связи, сохранять воспоминания и получать доступ к ним. Поэтому нейротрофины могут спасти дегенерирующие нервы и восстановить когнитивную функцию. [R]
В 1986 году д-р Рита Леви-Монтальчини получила Нобелевскую премию за открытие фактора роста нервов. Итальянский нобелевский лауреат выделила NGF (вместе со своим коллегой Стэнли Коэном) в 1952 году благодаря своим исследованиям некоторых раковых опухолей, которые показали исключительно быстрый рост нервной ткани. Затем, она использовала глазные капли с NGF для увеличения продолжительности своей жизни и здоровья. Умерла она в возрасте 103 лет в 2012 году. [R]
- Полезные свойства фактора роста нервов
- Помогает справиться с депрессией
- Помогает при рассеянном склерозе
- Помогает при болезни Альцгеймера
- Поддерживает фертильность
- Регулирует когнитивные функции
- Негативные эффекты
- Воспаление и боль
- Рак
- Как повысить фактор роста нервов (NGF)
- Вести здоровый образ жизни
- Ноотропы для повышения фактора роста нервов
- Заключение
Полезные свойства фактора роста нервов
Уровни содержания NGF в крови были значительно ниже у людей с тяжелыми депрессивными расстройствами по сравнению со здоровыми людьми. [R]
Физические упражнения повышают уровень NGF, что улучшает выживаемость нейронов в гиппокампе и одновременно уменьшает депрессию. Происходит это за счет увеличения количества вырабатывающих серотонин клеток в стволе мозга. [R]
В животной модели демиелинизирующих заболеваний мозга человека (например, рассеянный склероз), NGF задерживает начало, снижает воспаление и уменьшает повреждение тканей. Он способствует росту и восстановлению повреждений миелина.
Фактор роста нервов также непосредственно контролирует некоторые из основных структурных белков миелиновой оболочки. Еще, он стимулирует производство BDNF, что также важно для миелинизации нервов.
Из-за способности NGF защитить мозг и сбалансировать иммунную систему, он изучается для ряда нарушений мозга, в том числе Рассеянный склероз, болезнь Альцгеймера, и болезнь Паркинсона. [R, R, R]
При болезни Альцгеймера и синдроме Дауна способность мозга преобразовывать proNGF в зрелый NGF нарушена, что приводит к дисфункциональным нейронам. Причинами могут быть образование амилоид-β бляшек и воспаление мозга.
Вещества, которые повышают уровень ацетилхолина, доказали свою эффективность при болезни Альцгеймера. Недавние исследования на животных показывают, что NGF избирательно защищает нейроны, активированные ацетилхолином, что положительно сказывается на лечении болезни Альцгеймера. [R, R]
Фактор роста нервов принимает активное участие в работе репродуктивной системы. Это так важно, что его также называют фактором, вызывающим овуляцию (OIF).
Недавние исследования подтвердили, что у некоторых млекопитающих он вызывает овуляцию (высвобождение яйцеклеток из яичников), что может способствовать фертильности.
Низкий уровень NGF (и BDNF) в фолликулярной жидкости яичника снижает способность яичника вырабатывать яйцеклетки и может также быть связан с повышенным риском эндометриоза. [R, R]
Поддерживает память. Фактор роста нервов присутствует в гиппокампе, который ассоциируется с памятью. Например, одно исследование показало, что NGF жизненно важен для пластичности и обучаемости. Кроме того, сохранение памяти было хуже после блокировки фактора роста нервов. Это еще одно доказательство того, что нейротрофины полезны для памяти. Другое исследование также подтверждает, что NGF помогает справляться с проблемами памяти, особенно у стареющих мужчин и женщин. [R]
Таким образом, можно сделать вывод, что фактор роста нервов является важным фактором в поддержании памяти.
Способствует восстановлению после стресса. Одно исследование показывает, что фактор роста нервов помогает омолаживать ткани после того, как они испытывают острые или хронические стрессовые события. Таким образом, можно предположить, что выработка NGF во время или после стрессовых ситуаций способствует снижению общего стресса. [R]
Влияет на внимание, мотивацию, возбуждение и сознание. Базальный передний мозг состоит в основном из холинергических нейронов. NGF поддерживает выживание и сохранение преимущественно холинергических нейронов. Таким образом, он также поддерживает структуры, состоящие из холинергических нейронов, таких как базальный передний мозг. [R]
Поскольку базальный передний мозг играет главную роль в внимании, памяти, мотивации, возбуждении и сознании, NGF косвенно поддерживает эти когнитивные процессы.
Негативные эффекты
Уровень фактора роста нервов часто высок у людей с хроническими болями [R]:
- Интерстициальный цистит/синдром болезненного мочевого пузыря.
- Хронический простатит/синдром хронической тазовой боли.
- Остеоартрит.
- Диабетическая периферическая невропатия.
Увеличение NGF может быть частью воспалительного ответа на травму головного мозга. [R]
Фактор роста нервов является одним из механизмов, с помощью которого стресс усиливает воспаление и аутоиммунитет (нейроиммунные взаимодействия). [R]
Согласно исследованиям, NGF (так же как и BDNF) стимулирует распространение и выживание опухолевых клеток и способствует выработке новых кровеносных сосудов в опухолях. [R]
Как повысить фактор роста нервов (NGF)
Социализация. У мышей, выращенных в более социальной среде, в отдельных участках мозга (гиппокампе и гипоталамусе) отмечается более высокий уровень NGF. [R]
Йога. Даже один 20 минутный сеанс увеличивает выработку фактора роста нервов. [R]
Меньше переживать. Хронический легкий стресс снижает концентрацию NGF в гипоталамусе. [R]
Силовые тренировки. Упражнения с отягощениями повышает уровень фактора роста нервов в гиппокампе и подавляет апоптоз (запрограммированная гибель клеток). [R]
Влюбиться. Определенные поведенческие и психологические особенности, связанные с влюбленностью, вызывают повышение уровня NGF. [R]
Семакс. Пожалуй самый сильный ноотроп в плане повышения выработки нейротрофинов. Он увеличивает выработку как BDNF, так и NGF. Подробнее про Семакс>>>
Гриб львиная грива. Это съедобный гриб, который на вкус напоминает морепродукты. Он содержит активные вещества, которые стимулируют синтез NGF. Подробнее про Львиная грива>>>
Никотин. При повреждении нейронов спинного мозга, никотин оказывает нейропротекторное действие в виде повышения NGF. Подробнее про Никотин>>> [R]
Пирролохинолинхинон (PQQ). Это антиоксидант, который также стимулирует синтез и секреции фактора роста нервов. Подробнее про PQQ>>>
Ацетил-L-Карнитин (ALCAR). Данная аминокислота предотвращает некоторые дефициты, связанные со старением в центральной нервной системе (ЦНС). А именно ухудшение когнитивных функций, которое связано со старением и снижением уровня фактора роста нервов. Подробнее про ALCAR>>>
Гиперзин А. Это алкалоид, который обладает прямой нейротрофической активностью. Такая активность может быть полезна при лечении нейродегенеративных расстройств, таких как болезнь Альцгеймера. Подробнее про Гиперзин А>>>
Идебенон (Нобен). Это стимулятор, который вызывает увеличение синтеза фактора роста нервов и мРНК. Он также ослабляет поведенческие нарушения, вызванные болезнью Альцгеймера. Подробнее про Идебенон>>>
Селегилин. Селегилин и его метаболит (десметилселегилин) защищают нейроны путем активации эндогенного (собственного) синтеза NGF, BDNF и GDNF. Подробнее про Селегилин>>>
Ноопепт. Долгосрочное применение (28 дней) Ноопепт показали значительное увеличение нейротрофических факторов в гиппокампе. Подробнее про Ноопепт>>
Заключение
Фактор роста нервов является жизненно важным компонентом для здоровья и развития мозга. Повышать NGF становится необходимым только в старости. Но сохранять его концентрацию на должном уровне лишнем никогда не будет. Лучше поддерживать, чем восстанавливать.
Мы рассмотрели много классов соединений, играющих важнейшую роль в деятельности мозга: ионные каналы, молекулярные насосы, рецепторы, медиаторы, вторичные посредники. Однако существует еще одна группа веществ, важных для ЦНС – это факторы роста нервов (ФРН).
ФРН являются белками и состоят из более чем 100 аминокислот. Наиболее известен из них ФРН, выделенный впервые из слюнной железы мыши. Факторы роста могут и должны синтезировать все ткани, к которым в ходе эмбриогенеза направляются нервные отростки, т.е. аксоны мотонейронов, аксоны клеток вегетативной нервной системы, отростки клеток сенсорных ганглиев и т.п.
Функцией ФРН является обеспечение выживания нервной клетки на ранних стадиях развития эмбриона и, частично, у взрослого организма. Они также направляют прорастание отростков нервных клеток и установление контактов с органом-мишенью на поздних стадиях развития эмбриона и при травмах. Иными словами, ФРН являются своеобразными указателями, привлекающими растущие отростки. При этом каждая периферическая ткань, видимо, секретирует свой ФРН, а соответствующая группа нейронов ЦНС врожденно настроена на этот фактор.
Выделяясь клетками-мишенями, ФРН захватывается нервными отростками и переносится в сому. В результате он попадает в ядро нейрона, воздействует на ДНК и регулирует синтез ферментов, что обеспечивает рост, развитие и выживание нервной клетки. Такой механизм действия можно классифицировать как гормоноподобный. Сходным образом влияют на работу клеток, например, гормон щитовидной железы тироксин и гормон гипофиза соматотропин.
В качестве примера работы ФРН рассмотрим процесс прорастания аксона мотонейрона и образование нервно-мышечного синапса.
Когда контакт еще не установлен, никотиновые рецепторы к ацетилхолину равномерно распределены по всей мембране мышечного волокна. Конус роста аксона направляется к мышечной клетке, которая выделяет много ФРН. На конусе роста уже сформированы все механизмы (Са 2+ -каналы и пр.), которые включают выброс ацетилхолина при возбуждении мотонейрона. При установлении контакта аксона и мышечной клетки образуется синапс. Ацетилхолин начинает активировать мышечное волокно, и никотиновые рецепторы собираются на постсинаптическую мембрану. В результате концентрация рецепторов на постсинаптической мембране оказывается в 1000 раз больше, чем на остальной мембране.
Мышечное волокно, получившее возможность сокращаться, резко уменьшает синтез ФРН. Фактор роста теперь выделяется только в зоне синапса, что ограничивает число формируемых на волокне нервно-мышечных контактов. В итоге первый проросший аксон "монополизирует" весь ФРН, а остальные волокна дегенерируют. Более того, примерно половина мотонейронов эмбриона вообще не успевает сформировать ни одного контакта с мышечными клетками и погибает.
В итоге одно мышечное волокно управляется одним аксоном, хотя один аксон благодаря своим коллатералям может иннервировать сразу много волокон. Интересно, что если электрически раздражать эмбриональное мышечное волокно, то ФРН тоже прекращает выделяться, и нервно-мышечный синапс вообще не образуется. Если, напротив, денервировать волокно (перерезка аксона, необратимые блокаторы никотиновых рецепторов), то синтез ФРН опять увеличивается, и к волокну начинают прорастать другие аксоны. То же происходит и при травмах периферических нервов.
Возможность роста нервной ткани не отрицается полностью тем фактом, что нейроны не способны к воспроизведению, как другие клетки тела. Нейроны отличаются друг от друга своими длинными волокнами, протяженностью разветвлений на теле клетки и теми амебовидными структурами на концах, названными конусами роста, которые можно наблюдать в культуре ткани и даже in situ (рис. П-2). Конус роста толкает и давит на находящуюся перед ним ткань и может открывать путь, по которому он затем сможет медленно течь, удлиняя конец нервного волокна. Таким образом, рост может происходить при условии наличия свободного пространства, когда же его нет, амебовидный конец сокращается только для того, чтобы снова и снова продвигаться вперед. В центральной нервной системе рост обычно исключен потому, что элементы ткани расположены очень близко друг к другу. Правда, возникли большие сомнения в этом, так как результаты электронной микроскопии показали, что внеклеточное пространство, которое было найдено во всех частях организма, существует и вокруг нейронов.
Поэтому исследователи мозга стали наблюдать за ростом нервного волокна, создавая в мозгу свободное пространство. До недавнего времени такие попытки не приводили к значительным результатам: обычная реакция мозговой ткани на инсульт состоит в разрушении поврежденных частей и разжижении их, в образовании полости или кисты, оболочки которых препятствуют любой попытке проникновения в них. Однако современная техника дала исследователям инструмент, позволяющий избежать этой обычной реакции. Циклотрон излучает радиацию, которая резко замедляет процессы, происходящие в мягкой ткани. Таким образом,
Рис 11-2 Конус роста в действии Фрагменты фильма с замедленной съемкой Обратите внимание на заметные изменения формы от фрагмента к фрагменту (Pomerat, 1964)
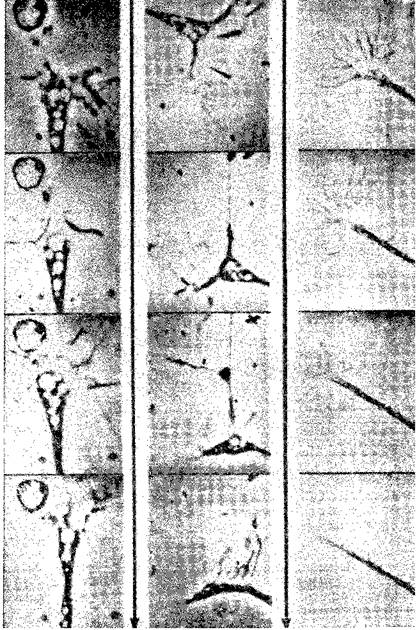
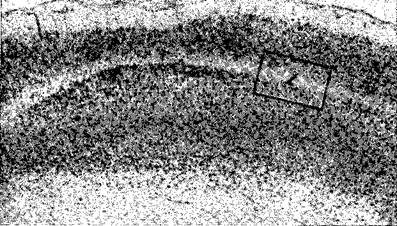
Рис П-3 Разрушение (I) в пятом слое постцентральной области коры кролика на 132-й день после воздействия пиковой дозы радиации в 48 000 рад Более тщательное исследование позволяет обнаружить многочисленные дендриты, достигающие области разрушения. Окрашено тионином (ХЗО) (Rose et al, 1961)
описанные разрушения можно произвести локально и только на том участке, на который направляется разрушающее воздействие. С помощью этого метода можно разрушать определенные слои коры взрослого кролика без видимого повреждения соседних. При этом методе повреждение ткани достаточно локально, так что обычно наблюдаемая реакция на большие разрушения нервной ткани не имеет места. Таким образом, создается пространство для активного роста нервных волокон (рис. И-3, П-4, П-5).
Срезы, сделанные спустя несколько недель или месяцев после такого повреждения, показывают, что сохранившиеся волокна становятся толще (Kruger, 1965). Это утолщение, по-видимому, является совершенно нормальным увеличением размера волокна. Так как диаметр волокна часто является индикатором его длины, то утолщение означает в этих случаях, что мог иметь место и рост нервного волокна. Более прямым доказательством роста является появление большого числа нормальных, четко ориентированных волокон, которые не были видны ни до, пи сразу же после воздействия циклотрона (Rose, Malis, Baker, 1961). Следовательно, рост волокон возможен и в зрелом мозгу, если для этого есть благоприятные условия.
Эти эксперименты, конечно, не могут показать, является ли рост действительно результатом жизненного опыта; для этого должен быть проведен другой тип исследований. Чтобы проверить-
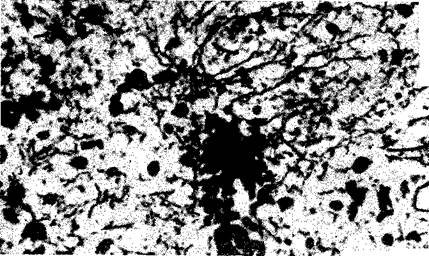
Рис. II-5. Разрастание аксонов в области разрушения в стриарной
коре крысы на 19-й день после облучения альфа частицами в 9000 рад
Окрашено по Водиану (Х390) (Kruger, 1965).
Таблица ll-l
Влияние различного живленного опыта н 1 затылочную кору крыс о возрасте от 25 до 105 дней, содержащихся в насыщенных или обедненных стимулами условиях
Процентное рчзличие НУ>ОУ

6,4
а —различие в весе животных в этих экзперлментах 1,0% " —различие в ве
е животных в этих эк°пер 1ментах Ь,&% НУ—условия эк перичента с насыщенной стимуляцией ОУ—условия эк периаента с обедненной стимуляцией НД—недостоверно Р —статистиче-ки вероятный уровень значимости
(ПоЕ L Bennett, M R Rosenzweig, 1970, р 183)
На процессы роста нейронов влияют многочисленные факторы, в том числе химические агенты – нейрональные индукторы. В раннем эмбриогенезе у позвоночных факторы индукции образуются при взаимодействии нейроэктодермы и хордомезодермы. Так, в самом начале онтогенеза нейрона клетками хордомезодермы секретируется низкомолекулярный протеин, направляющий развитие недетерминированной мультипотентной нейральной клетки-предшественницы в нейробласт. Есть данные о том, что одновременно (или чуть позже) секретируется белок, обеспечивающий различие между нейронами. Сама нервная пластина обособляется в конце второй недели эмбриогенеза. Её клетки делятся, превращаясь в узкие цилиндрические, резко отличающиеся от соседних эктодермальных эпителиоцитов. Именно они являются мультипотентными предшественниками нейро- и глиогенеза. На следующем этапе клетки, специализирующиеся как нейробласты, превращаются из аполярных в веретеновидные. Такой нейробласт устанавливает ранние контакты с соседними клетками, у него возникает конус роста, от которого идет формирование аксона, так что нейробласт превращается в растущую монополярную клетку. Основные процессы этого этапа развития нейронных систем связаны с интенсивным ростом аксонных, а потом и дендритных ветвлений с усложнением межнейронных связей. Именно в этот период на нейроны действуют множество индуцирующих факторов – как внешних, так и внутренних.
Исследования конца XX в. показали, что сохранение и дифференцировка нейронов, рост и ремоделирование аксонов и дендритов, состояние цитоскелета, экзоцитоз, формирование и пластичность синапсов регулируются факторами роста нервной ткани. Большинство полипептидных факторов роста действует одновременно по паракринному (стимулирующему соседние клетки) и аутокринному (когда, фактор роста, секретируемый клеткой, индуцирует синтез своего рецептора в той же самой клетке) механизмам. Отдельные факторы, такие как инсулиноподобные факторы роста, способны оказывать эндокринное действие (дистантное, осуществляемое через жидкие среды организма на многие клеточные мишени). Помимо этого, существует еще один способ действия факторов роста, который получил название интракринного. Факторы роста при этом не секретируются и не нуждаются в поверхностных рецепторах, опосредующих их активность. Они остаются внутри клетки и действуют в качестве посредников, регулируя ее функции. Ряд цитоплазматических факторов роста и цитокинов, действующих подобным образом, достаточно хорошо изучен. Это предшественники интерлейкинов, цилиарный нейротрофический фактор, и др. Эти факторы вызывают заметный биологический эффект до появления их на поверхности клетки-продуцента или в окружающем ее пространстве. В регуляторных белках, обладающих интракринным действием, имеются сигнальные последовательности, обеспечивающие внутриклеточную локализацию. До сих пор очень мало известно о внутриклеточной компартментализации факторов роста и их значении в рассматриваемых процессах. Полагают, что различные внутриклеточные пулы факторов роста могут использовать пара-, ауто- и интракринные механизмы для достижения специфического клеточного ответа.
В частности доказано, что ведущая роль в процессах дифференцировки аполярного нейробласта в монополярный принадлежит цитокинам. Цитокины – группа гормоноподобных пептидов – во взрослом организме синтезируются и секретируются клетками иммунной системы и другими типами клеток. Разнообразные биологические функции цитокинов подразделяются на три группы: они управляют развитием и гомеостазом иммунной системы, осуществляют контроль за ростом и дифференцировкой клеток крови (системой гемопоэза) и принимают участие в неспецифических защитных реакциях организма, оказывая влияние на воспалительные процессы, свертывание крови, кровяное давление. Вообще цитокины принимают участие в регуляции роста, дифференцировки и продолжительности жизни клеток, а также в управлении апоптозом. На сегодняшний день известен обширный перечень цитокинов, регулирующих развитие и функционирование центральной нервной системы. Наиболее известным является фактор роста нервов – NGF (nerve growth factor) – пептид, ускоряющий рост определенных типов нейронов. Изучение фактора роста нервов позволило значительно глубже понять мембранный и рецепторный компоненты клеточного метаболизма. Благодаря особенностям строения нейронов, рецепторопосредованный захват, ретроградный транспорт и разрушение фактора роста нервов в лизосомах нервных клеток были описаны за десять лет до начала детального изучения ретроградного транспорта и эндоцитоза в других системах организма. Исключительная важность первых экспериментов по изучению фактора роста нервов была оценена в 1986 г. присуждением Рите Леви-Монтальчини и Стэнли Коэну Нобелевской премии "в знак признания открытий, имеющих важнейшее значение для раскрытия механизмов регуляции роста клеток и органов".
Нервная пластинка и нервная трубка содержат ограниченное число клеток, во много крат уступающее числу дифференцированных нейронов взрослого мозга. Это связано с тем, что клетки нервной трубки интенсивно пролиферируют. Генетически запрограммированные процессы детерминации вносят коррективы в особенности самого пролиферативного периода, определяющегося числом митотических делений клеток предшественниц. Длительность клеточных циклов также начинает разниться – от нескольких часов до нескольких суток. Все это происходит на фоне миграции нейронов к определенным областям промежуточного или окончательного назначения. Начиная со стадии терминальной дифференцировки, пролиферация нейронов в мозгу млекопитающих становится невозможной.
Достигнув района окончательной локализации, нейроны агрегируют друг с другом и формируют ядра серого вещества, иннервирующие определенные двигательные или органные структуры. Избирательная адгезия с последующей пространственной ориентацией, определяющие конечную форму нейрона – это общие свойства нервных элементов, они обеспечиваются специализированными рецепторными молекулами клеточной мембраны. Одними из наиболее известных мембранных белков являются интегрины – поверхностные клеточные рецепторы, взаимодействующие с внеклеточным матриксом и передающие различные межклеточные сигналы. От них зависит форма клетки, её подвижность, они участвуют в регулировке клеточного цикла. Лигандами для интегринов чаще всего являются различные белки внеклеточного матрикса: коллагены, ламинин, фибронектины и др. В нейрогенезе наиболее заметную роль играют интегрины с кальций-зависимой адгезией, а также трансмембранные гликопротеины – кадгерины, обеспечивающие реакции слипания нейроцитов (рис. 3). Кадгерины являются тканеспецифичными молекулами. Для нервной системы особенно характерны кадгерины N-CAM (neuron cell adhesion molecule), молекулы адгезии L1 и нейрофасцин.
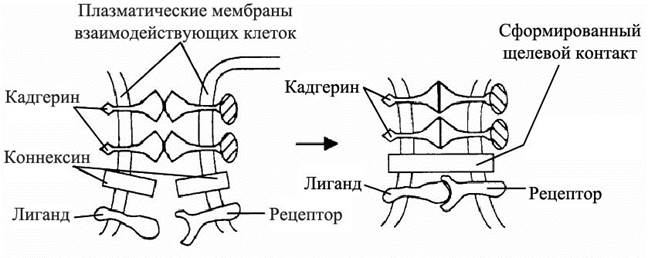
Рис. 3. Роль молекул адгезии в межклеточной коммуникации. Взаимодействие трансмембранных молекул адгезии (кадгерины) обеспечивает узнавание клеточных партнёров и их прикрепление друг к другу (адгезию), что позволяет клеткам-партнёрам сформировать щелевые контакты, а также передавать сигналы от клетки к клетке не только при помощи диффундирующих молекул, но и путём взаимодействия встроенных в мембрану лигандов со своими рецепторами в мембране клетки-партнёра.
В итоге, миграция и взаимная топографическая перестройка нервных элементов приводят к формированию новых отделов эмбрионального мозга, превращая эмбриональную нервную трубку в анатомически сложно организованную структуру.
Как показали исследования XVIII-XX веков, постнатальный нейрогенез возможен у некоторых эволюционных групп. В частности доказана возможность как репаративной, так и физиологической регенерации нервной ткани у рыб, амфибий и рептилий. Однако высшие позвоночные – птицы и млекопитающие – утратили способность к репаративной регенерации. Физиологическая регенерация у этих таксонов остается вопросом дискуссионным. Тем не менее, в мозгу взрослых млекопитающих нейрогенез все же имеет место, но в строго ограниченных зонах. Эти зоны являются рудиментами нейрогенных зон эмбрионального мозга. Предположительно в этих участках сохраняются клетки радиальной глии, которые, как выяснилось, могут быть не только предшественниками астроцитов, но и предшественниками нейронов, как в эмбриогенезе, так и в постнатальном онтогенезе позвоночных. Самая обширная из этих зон нейрогенеза – субвентрикулярная зона в стенках боковых желудочков (большие полушария). Здесь стволовые клетки генерируют нейробласты, которые мигрируют в обонятельную луковицу, где и превращаются в зрелые нейроны. Небольшая зона нейрогенеза расположена в субгранулярной зоне зубчатой фасции (связки) гиппокампа. Оттуда новые нейроны мигрируют на небольшое расстояние в пределах гиппокампа.
По мнению А.Г. Бойко (2007), дифференцировка клеток радиальной глии в астроциты является вероятным механизмом старения млекопитающих. Исчезновение радиальной глии в онтогенезе является причиной постмитотичности мозга млекопитающих, ведет к накоплению необратимых явлений в нейронах – клеток с ограниченными возможностями и сроком службы, и исключает замену структур, исчерпавших свой жизненный ресурс. По истечении определенного периода изменения в нейронах, особенно гормонсинтезирующих, начинают неблагоприятно сказываться на состоянии эндокринной системы и на общем гомеостазе организма. Данная гипотеза (как и другие, например теломеразная) не противоречит проявлению общего эволюционного принципа – замены внешних факторов смерти нестареющих многоклеточных организмов на внутренние.
Онтогенез нервной системы
Зачаток нервной системы у позвоночных животных начинает развиваться уже в конце стадии гаструлы. На следующей стадии – нейрулы – образуются нервная трубка, хорда и сомиты. Процесс, благодаря которому осуществляется формирование нервной пластинки и превращение ее в нервную трубку, называется нейруляцией.
Вначале из клеток эктодермального зародышевого листка образуется нейроэктодермальная, или медуллярная, пластинка, края которой в результате неравномерного размножения ее клеток сближаются, затем смыкаются – образуется медуллярная (нервная) трубка. В местах смыкания трубки возникает нервный гребень (рис.4).
В пределах каждой из областей в развивающейся нервной системе судьба клеток в значительной степени зависит от их расположения и клеточного окружения. Фенотип, в который дифференцируются клетки нервного гребня, определяется сигналами, поступающими от соседних клеток. Таким образом, если клетки нервного гребня были пересажены в раннем возрасте, они будут дифференцироваться в соответствии со своим новым расположением.
 |
Уже на третьей – четвертой неделе (у человека) выделяются две области нервной трубки: дорсальная (крыловидная пластинка) и вентральная (базальная пластинка). Из крыловидной пластинки развиваются чувствительные и ассоциативные элементы нервной системы, из базальной – моторные. Структуры переднего мозга у человека целиком развиваются из крыловидной пластинки.
Рис. 4. Микрофотография поперечного среза двухнедельного зародыша курицы (По Албертс Б. и др., 1994).
В дальнейшем из задней части нервной трубки, отстающей в росте, образуется спинной мозг, из передней, развивающейся более интенсивно, – головной мозг. Канал медуллярной трубки впоследствии сохраняется в виде центрального канала спинного мозга и желудочков головного мозга. Вследствие развития передней части медуллярной трубки образуются мозговые пузыри: вначале появляются два пузыря, затем задний пузырь делится еще на два. Образовавшиеся три пузыря дают начало переднему (ргоsencephalon), среднему (mesencephalon) и ромбовидному (rhombencephalon) мозгу.
Затем наступает этап формирования целой серии изгибов, которые предстоит претерпеть растущему мозгу, прежде чем он примет свои взрослые очертания. Изгибы разграничивают основные структурные единицы и разделяют широкие внутренние полости, которые потом станут желудочками мозга.
В полтора месяца эмбрионального развития человека большой пузырь переднего мозга подразделяется на конечный мозг (telencephalon), из которого позже разовьются большие полушария, и промежуточный мозг (diencephalon), из которого будут сформированы таламус и гипоталамус. Эти ранние стадии развития головного мозга напоминают соответствующие стадии формирования менее сложного мозга позвоночных. К двум месяцам уже образован основной (среднемозговой) изгиб головного мозга: передний мозг и промежуточный мозг загибаются вперед и вниз под прямым углом к продольной оси нервной трубки. Позже формируются еще два изгиба: шейный и мостовой. В этот же период первый и третий мозговые пузыри разделяются дополнительными бороздами на вторичные пузыри, при этом появляется 5 мозговых пузырей. Задний пузырь (rhombencephalon) делится на два пузыря, из которых образуется задний мозг (metencephalon) и продолговатый, или добавочный, мозг (medulla oblongata, myelencephalon).
В течение пятой – десятой недели развития начинается рост и дифференцировка конечного мозга: образуются кора и подкорковые структуры. На этой стадии развития появляются мозговые оболочки, формируются ганглии вегетативной нервной системы. Спинной мозг приобретает окончательное строение.
Конечный мозг в этот период проходит три стадии развития. На первой стадии развития появляются обонятельные доли мозга, гиппокамп, лимбическая система. Вторая стадия связана с ростом базальных ганглиев. Третья стадия развития конечного мозга включает формирование новой коры больших полушарий – неокортекса. Когда у приматов скорость роста неокортекса достигает максимума (ок. 250 тыс. клеток в минуту), поверхность его начинает образовывать извилины, которые позволяют увеличить объем корковой ткани без увеличения общих размеров мозга.
В следующие 10 – 20 недель беременности завершается формирование всех отделов головного мозга, идет процесс дифференцировки мозговых структур, который заканчивается только с наступлением половозрелости. Полушария становятся самой большой частью головного мозга. Выделяются основные доли (лобная, теменная, височная и затылочная), образуются извилины и борозды больших полушарий. В спинном мозге в шейном и поясничном отделах формируются утолщения, связанные с иннервацией соответствующих поясов конечностей. Окончательный вид приобретает мозжечок.
В процессе развития нервной системы важную роль играет миелинизация нервных волокон. Следы миелина обнаруживаются в нервных волокнах задних и передних корешков уже на четвертом месяце внутриутробной жизни плода человека. К концу четвертого месяца миелин выявляется в нервных волокнах, составляющих восходящие, или афферентные (чувствительные), системы боковых канатиков, тогда как в волокнах нисходящих, или эфферентных (двигательных), систем миелин обнаруживается на шестом месяце. Приблизительно в это же время наступает миелинизация нервных волокон задних канатиков. Формирование миелиновых оболочек нервных волокон корково-спинномозговых (пирамидных) путей начинается на последнем месяце внутриутробной жизни и продолжается в течение года после рождения. Это свидетельствует о том, что процесс миелинизации нервных волокон распространяется вначале на филогенетически более древние, а затем на более молодые структуры. От последовательности миелинизации определенных нервных структур зависит очередность формирования их функций. Этим объясняется позднее созревание пирамидной системы и постепенное начало проявления ее функции в первые два года жизни ребенка. В это время бурно развиваются нервные элементы коры большого мозга, где происходит не только миелинизация нервных волокон, но и функциональная дифференциация клеточных элементов и их постепенное созревание, которое длится в течение первого десятилетия.
В процессе эмбрионального развития головного мозга полости мозговых пузырей видоизменяются и превращаются в систему мозговых желудочков, которые сохраняют связь с полостью спинномозгового канала. Центральные полости больших полушарий головного мозга образуют боковые желудочки довольно сложной формы. Боковые желудочки соединяются с полостью промежуточного мозга, которая является III желудочком. Через специальный проток (сильвиев водопровод) III желудочек соединяется с IV желудочком; IV желудочек образует полость заднего мозга и переходит в спинномозговой канал.
В постнатальном периоде постепенно происходит окончательное созревание всей нервной системы, в частности ее самого сложного отдела — коры большого мозга, играющей особую роль в мозговых механизмах условно-рефлекторной деятельности, формирующейся с первых дней жизни.
Таким образом, нервная система проходит длительный путь развития, являясь самой сложной системой, созданной эволюцией. Филогенетические законы развития нервной системы были сформулированы в 30-х годах ХХ века выдающимся учеником В.М. Бехтерева – М.И. Аствацатуровым,– основателем биогенетического направления в неврологии.
Сущность этих законов сводится к следующему:
1. Нервная система возникает и развивается в процессе взаимодействия организма с внешней средой. Нервная система лишена стабильности, изменяясь и непрерывно совершенствуясь в фило- и онтогенезе.
2. Сложный и подвижный процесс взаимодействия организма с внешней средой вырабатывает, совершенствует и закрепляет новые виды реакций, лежащих в основе формирования новых функций.
3. Развитие, закрепление более совершенных и адекватных реакций и функций представляют собой результат действия на организм внешней среды, т. е. приспособления его к данным условиям существования. Борьба за существование как биологический процесс имеет место, но не является ведущим фактором в совершенствовании организма и его функций. Основное в развитии и совершенствовании функций нервной системы – приспособление (адаптация) организма к среде.
4. Функциональной эволюции (физиологической, биофизической, биохимической) соответствует эволюция морфологическая. Вновь приобретенные функции постепенно закрепляются. Наряду с совершенствованием функции происходит развитие и совершенствование ее морфологического субстрата.
5. Древние функции не исчезают с появлением новых, а вырабатывается их определенная иерархия, соподчиненность.
6. В процессе эволюции древние аппараты нервной системы не отмирают, а только видоизменяются, приспосабливаются к новым внешним условиям.
7. Онтогенез нервной системы повторяет в общих чертах ее филогенез.
8. При выпадении новых функций нервной системы проявляются ее древние функции. Многие клинические признаки заболеваний, наблюдаемые при нарушении функций эволюционно более молодых отделов нервной системы, являются проявлением функций более древних структур, т. е. в патологических условиях наступает определенный регресс нервной системы на низшую ступень филогенетического развития.
9. Самыми ранимыми отделами нервной системы являются филогенетически более молодые, в частности кора большого мозга, которая еще не выработала защитных механизмов, в то время как древние отделы на протяжении тысячелетий взаимодействия с внешней средой успели выработать и накопить определенные механизмы противодействия вредным факторам.
10. Чем филогенетически более молодыми являются нервные структуры, тем в меньшей степени они обладают способностью восстановления (регенерации).
Читайте также:
