
Физиология мышечной и нервной ткани
Согласно современным представлениям, нервная и мышечная ткани могут находиться в трех основных состояниях — физиологическом покое, возбуждении и торможении.
Физиологический покой — это такое состояние, когда ткань или орган не проявляет признаков присущей им деятельности.
Возбуждение — деятельное состояние живой ткани, в которое она приходит под влиянием раздражения.
Торможение — такое состояние, когда деятельность ткани или органа ослабляется или полностью прекращается.
Раздражители.При раздражении мышца приходит в состояние возбуждения, которое распространяется по мышце и вызывает ее сокращение. По биологическому действию раздражители бывают адекватные и неадекватные. Адекватный — это такой раздражитель, к которому данный орган или ткань приспособились в процессе эволюции. Для мышцы адекватным раздражителем является нервный импульс. Неадекватными — будут такие раздражители, действию которых ткань или орган в естественных условиях обычно не подвергаются. Так, сокращение мышцы можно вызвать механическим раздражением (укол, кислота, ток) и т.д.
Все раздражители по своей силе деляг на пороговые, подпороговые и сверхпороговые. Пороговыми называют минимальные раздражители, которые могут вызвать возбуждение. Подпороговые — это раздражители, сила которых меньше пороговой, сверхпороговые — раздражители более сильные, чем пороговые.
Биоэлектрические явления.Возникновение и распространение возбуждения связано с изменением электрического заряда на поверхности клеточной мембраны и внутри клетки (рис. 4.19).
Потенциал покоя.Мембрана нервной или мышечной клетки в состоянии покоя с наружной стороны заряжена положительно, а с внутренней — отрицательно. Разность зарядов между наружной и внутренней поверхностью клеточной мембраны в состоянии физиологического покоя клетки называется потенциалом покоя, или мембранным потенциалом.
Потенциал действия. Под влиянием раздражения проницаемость клеточной мембраны для ионов натрия повышается в сотни
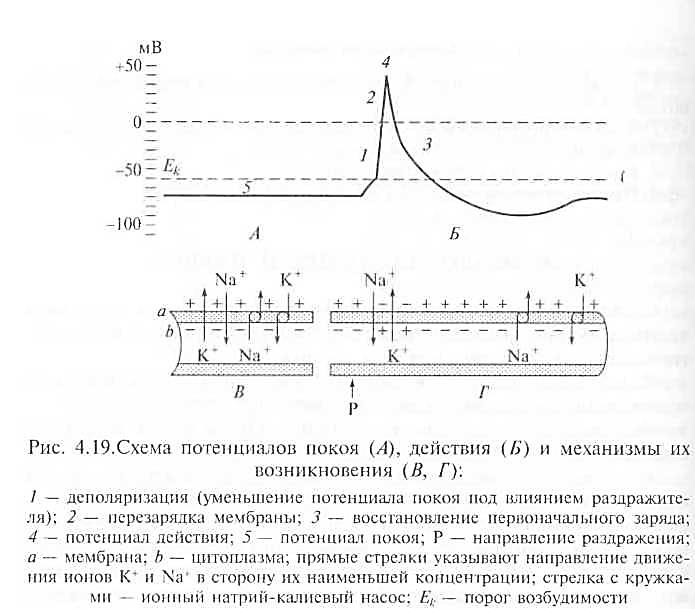 |
Рис. 4.19.Схема потенциалов покоя (А), действия (5) и механизмы их
возникновения (В, Г):
1 — деполяризация (уменьшение потенциала покоя под влиянием раздражите-
ля); 2 — перезарядка мембраны; 3 — восстановление первоначального заряда;
4 — потенциал действия; 5 — потенциал покоя; Р — направление раздражения;
а — мембрана; b — цитоплазма; прямые стрелки указывают направление движе-
ния ионов К + и в сторону их наименьшей концентрации; стрелка с кружка-
ми — ионный натрий-калиевый насос; Ек — порог возбудимости
раз. Происходит перезарядка клеточной мембраны: внутренняя ее сторона заряжается положительно, а наружная — отрицательно. Затем значительно повышается проницаемость мембраны для ионов калия, они начинают быстро перемещаться из клеток во внеклеточную жидкость и вновь восстанавливают первоначальный заряд мембраны. Пикообразное колебание потенциала, возникающее в результате кратковременной перезарядки мембраны и последующего восстановления ее заряда, называется потенциалом действия. Такие колебания потенциала продолжаются 1 — 5 мс.
Проведение возбуждения. Круговой ток, проходя через участок покоя, раздражает его и вызывает появление потенциала действия (рис. 4.20). Когда в одном из участков нервной клетки, нервного или мышечного волокна возникает возбуждение и появляется потенциал действия, между возбужденным и соседним невозбужденным участком возникает разность потенциалов. Эта разность потенциалов является причиной появления круговых (локальных) токов. Внутри клетки ток идет от возбужденного участка к участку покоя, по внешней стороне — от участка покоя к возбужденному.
В участке, который ранее был возбужден, происходит восста
Рис. 4.20. Схема распрост-
ранения возбуждения:
А — волокна мышц и безмякотных нервных волокон; Б —
мякотное нервное волокно; 1 — мембрана; 2 — волокно;
а, Ь, с — перехваты Ранвье; заштрихованы возбужденные
участки; светлые — участки покоя; стрелками показано движение круговых токов

новление потенциала покоя. Данный процесс многократно повторяется и обусловливает распространение импульсов возбуждения. Скорость проведения возбуждения по скелетным мышцам 12 —15 м/с, гладким мышцам — 1 — 18 см/с, в безмякотных нервах — 0,5 — 3 м/с.
Основные свойства живой ткани.Любая живая клетка обладает свойствами раздражимости, возбудимости и лабильности (функциональной подвижности).
Раздражимость. С раздражимостью связаны все процессы роста и размножения клеток и тканей. На разнообразные воздействия внешней или внутренней среды — раздражения протоплазма живой клетки отвечает специфической реакцией, при которой происходят усиление или ослабление обмена веществ, количественное и качественное его изменение. Эта реакция на раздражение получила название раздражимости, или реактивности.
Возбудимость — свойство нервной или мышечной ткани отвечать на действие раздражителя возбуждением. Для измерения величины возбудимости определяют порог возбудимости и хронаксию. Порогом возбудимости называется наименьшая сила раздражителя, способная вызвать возбуждение; его выражают в вольтах постоянного тока.
Лабильность — скорость, с которой в ткани возникает и успевает закончиться полный период отдельного импульса возбуждения. Мера лабильности — это максимальное число импульсов возбуждения за 1 св ответ на такое же максимальное число раздражений. Высокой лабильностью обладают мякотные нерпы, у без- мякотных нервов она низкая. Так, максимальный ритм возбуждений мякотного нервного волокна 500, безммкотного — 200. Максимальный ритм возбуждений скелетной мышцы 200 импульсов в 1 с, а гладкой мышцы — в десятки раз меньше.
При очень частых раздражениях сокращения мышцы уменьшаются или даже совсем прекращаются. Такая частота называется пессимальной, или пессимумом. Пессимум возникает вследствие того, что возбуждение еще не закончилось и ткань находится в состоянии абсолютной или относительной рефрактерное™, а на нее действует новое раздражение. Частые раздражения, превышающие меру лабильности, вызывают не возбуждение, а торможение.
Учение Н. Е. Введенского о парабиозе устанавливает связь между возбуждением и торможением. Возбуждение и торможение — это различные реакции ткани на раздражение, исход которогозависит от лабильности. При высокой лабильности возникает возбуждение, снижение лабильности вызывает торможение.
ФИЗИОЛОГИЯ МЫШЦ
Скелетные мышц, входящие в состав опорно-двигательного аппарата позвоночных животных, состоят из группы мышечных пучков, каждый из них составлен из тысяч мышечных волокон, которые представляют собой клетки цилиндрической формы длиной до 12 см и диаметром 10 — 100 мкм. Каждое волокно окружено оболочкой сарколеммой и содержит тонкие нити — миофибрил-лы (рис. 4.21). Поперечные мембраны делят каждую миофибриллу на отдельные участки — саркомеры. Сократимым веществом мышечного волокна являются миофибриллы, состоящие из множества (около 2500) тонких и толстых белковых нитей — протофибрилл. Толстые протофибриллы образованы из белка миозина, тонкие — из актина. Нити актина прикреплены к мембране саркомера; они образуют светлые участки миофибриллы. В темных участках находятся нити миозина. Акгиновые нити частично входят своими концами в промежутки между миозиновыми нитями. Нити актина и миозина соединяются между собой многочисленными поперечными мостиками, которые образованы скрученными в спираль отростками — мостиками миозиновой нити. Чередование нитей в миофибрилле обусловливает ее поперечную исчерченность.

Рис. 4.21. Схема чередова-
ния нитей в миофибрилле:
А — электронно-микроскопическая фотография мио-
фибриллы; Б — строение миофибриллы; Z — поперечная
мембрана, разделяющая миофибриллу на отдельные уча-
стки — саркомеры; 1 — миозиновая нить, 2 — актиновая нить
Гладкие мышцы, образующие стенки внутренних органов, состоят из клеток различной формы длиной от 30 до 500 мкм и диаметром 2—10 мкм; у клеток имеются боковые отростки, благодаря которым клетки гладких мышц группируются в длинные пучки. Они, в свою очередь, при помощи тяжей соединяются друг с другом, обеспечивая деятельность мышцы как единой системы. Сократительным аппаратом гладких мышц являются миофибриллы, в которых содержатся тонкие актиновьге нити с прикрепленными к ним короткими отростками миозиновых нитей, называемых димерами.
Свойства скелетных мышц.Проведение возбуждения в мышцах происходит изолированно, т. е. потенциалы действия не переходят с одного мышечного волокна на другое. Возбудимость скелетныхмышц меньше возбудимости нервов. Нервное волокно оканчивается в середине мышечного волокна, поэтому возбуждение распространяется в обе стороны со скоростью 4—15 м/с.
Скелетная мышца является упругим телом. Если к мышце подвесить груз, то она растягивается, это свойство называется растяжимостью. Эластичностью мышцы называется возвращение мышцы к первоначальной своей длине после растяжения, когда будет удален груз. Пластичностью мышцы называется свойство сохранять удлиненную форму после удаления груза, вызвавшего ее растяжение. В скелетных мышцах лучше выражено свойство эластичности, чем пластичности.
Виды мышечных сокращений.При нанесении на мышцу одиночного раздражения она отвечает одиночным сокращением. Записывая это сокращение на двигающейся ленте какого-либо самописца (например, в простейшем случае на кимограф), можно отметить три периода: латентный — период от раздражения до начала сокращения, период сокращения и период расслабления. У икроножной мышцы лягушки одиночное сокращение длится 0,1 с, у млекопитающих — 0,04 — 0,1 с. В латентный период в мышце происходят процессы, при которых освобождается энергия для мышечного сокращения.
Если к мышце поступает несколько частых импульсов возбуждения, наступает длительное сокращение мышцы, которое называется тетаническим, или тетанусом. В зависимости от частоты возбуждений тетанус будет зубчатым или гладким (рис. 4.22).
Зубчатый тетанус наблюдается при такой частоте импульсов возбуждений, когда каждый импульс действует на мышцу в тот момент, когда она уже начинает расслабляться. Если же импульсы возбуждения настолько частые, что они воздействуют на мышцу до начала ее расслабления, то получатся длительные непрерывные сокращения мышцы — гладкий тетанус.
 |
Рис. 4.22. Сокращение скелетной мышцы при различной частоте раздра-
жения:
А — одиночное сокращение в ответ на одно раздражение; Б — зубчатый тетанус
(до 15 раздражений в 1 с); В — гладкий тетанус (свыше 25 раздражений); М —
механограмма — запись сокращения мышц; Р — частота раздражений; 1 — фаза
сокращения; 2 — фаза расслабления
Механизм мышечного сокращения (теория скольжения).В состоянии покоя тонкие нити актина лежат частично своими концами в промежутках между толстыми миозиновыми нитями. При возбуждении мышцы под влиянием потенциала действия нити актина сдвигаются в промежутке между нитями миозина, т. е. скользят навстречу друг другу, вызывая сокращение миофибрилл (рис. 4.23). Считают, что скольжение происходит в результате укорочения мостиков — боковых отростков миозиновых нитей под влиянием ионов кальция. Укорачиваясь, мостики тянут актиновую нить навстречу миозиновой.
Работа и сила мышц.При сокращении мышца укорачивается и тем самым совершает работу. Она будет наибольшей при средних нагрузках и среднем ритме сокращений.
Силу мышцы определяют по максимальному сокращению ее при поднятии максимального груза. Наибольшей силой обладают мышцы с косыми волокнами, имеющими перистое строение.
Утомление мышц.В процессе сокращений мышцы утомляются, при этом понижаются их возбудимость, лабильность и величина сокращения. В утомленной мышце понижается содержание гликогена и накапливаются продукты обмена. В опытах на нервно-мышечном препарате Н. Е. Введенский установил, что прежде всего утомляются синапсы в связи с их низкой лабильностью. В целом организме утомление в первую очередь наступает в нервных центрах коры больших полушарий. И. М.Сеченов доказал, что быстрое восстановление работоспособности утомленных мышц наступает не при полном их покое, а при работе других, до этого не сокращавшихся мышц. Импульсы от вновь вовлеченных в работу мышц повышают возбудимость нервных центров, возбуждение одних нервных центров снижает и даже снимает утомление других центров. Утомление зависит от состояния симпатической нервной системы и желез внутренней секреции: утомленная мышца вновь
Рис. 4.23. Схема расслабленной (А) и сократившейся (Б) миофибриллы
 |
начинает сокращаться при раздражении симпатического нерва или введении адреналина, усиливающих обмен. Тренировка мышц позволяет отсрочить наступление утомления. При ней увеличивается объем мышц в результате утолщения мышечных волокон, повышается содержание гликогена, АТФ и креатинфосфата, ускоряются восстановительные процессы.
Тонус мышц. Тонус скелетных мышц играет важную роль для поддержания определенного положения тела в пространстве и деятельности двигательного аппарата. Скелетные мышцы в покое расслабляются не полностью, а находятся в некотором напряжении, т.е. тонусе. Он обусловлен поступлением к мышцам редких нервных импульсов, возбуждающих мышечные волокна не одновременно, а поочередно.
Гладкие мышцы обладают способностью длительно находиться в тонусе. Длительные тонические сокращения этих мышц особенно отчетливо выражены в сфинктерах полых органов, в стенках кровеносных сосудов.
Известно, что под влиянием раздражителя живые клетки и ткани из состояния физиологического покоя переходят в состояние активности. Наибольшая ответная реакция среди тканей на раздражение наблюдается со стороны нервной и мышечной ткани. Основными свойствами нервной и мышечной ткани являются возбудимость, возбуждение, проводимость, рефрактерность и сократимость.
Возбудимость — это способность живой ткани отвечать на действие раздражителя изменением физиологических свойств и возникновением процесса возбуждения. Возбуждение — это активный физиологический процесс, который возникает в ткани под влиянием раздражителей и характеризуется изменением уровня обменных процессов в тканях, выделением энергии, сокращением мышечной ткани, выделением секрета, генерацией нервного импульса. Проводимостью называют способность живой ткани проводить волны возбуждения (биопотенциалы). Рефрактерность — это временное снижение возбудимости ткани, которое возникает в результате возбуждения. Лабильность — это зависимость ткани от особенностей обменных процессов, которая может возбуждаться определенное количество раз за единицу времени.
Различают электрические, химические, механические и температурные раздражители, которые способны вызвать ответную реакцию со стороны возбудимых тканей. По биологическим признакам раздражители могут быть адекватными и неадекватными, а по силе — подпороговымя, пороговыми и надпороговыми.
Решающее значение для появления возбуждения принадлежит силе раздражителя (закон раздражения). Существует определенная зависимость между силой раздражения и ответной реакцией. Чем больше сила раздражителя, тем выше, до соответствующего уровня, ответная реакция со стороны возбудимой ткани. Большое значение имеет и продолжительность действия раздражителя. Зависимость между силой раздражителя и продолжительностью его действия, необходимого для появления минимальной ответной реакции, определяется кривой силы — времени (рис. 140). Минимальная сила тока (напряжения), способная вызвать возбуждение, называется реобазой (отрезок ординаты (ОА). Кроме реобазы, важным параметром кривой силы является хроноксия. Последняя отражает наименьший промежуток времени, во время которого ток, равный по силе удвоенной реобазе (отрезок абсциссы OF), вызывает в ткани возбуждение. По величине хроноксии судят о скорости появления возбуждения в ткани: чем меньше хроноксия, тем быстрее возникает возбуждение. Приспособление возбудимой ткани к медленному нарастанию силы раздражителя получило название аккомодации. Последняя обусловлена тем, что за время нарастания силы раздражителя в ткани происходят активные изменения, которые повышают порог раздражения и препятствуют развитию возбуждения. Таким образом, скорость нарастания раздражения во времени называют градиентом раздражения.
Рис. 140. Кривая силы — времени
Закон градиента раздражения — это реакция на раздражитель, которая зависит от срочности или крутизны нарастания раздражителя за определенное время: чем выше градиент раздражения, тем сильнее (до определенных пределов) ответная реакция возбудимого объекта.
Потенциал покоя (мембранный потенциал) — это разность потенциалов между наружной поверхностью клетки и ее внутренним содержимым; он составляет около 60—90 мВ в зависимости от особенности той или иной клетки.
Потенциал действия (потенциал возбуждения) возникает при воздействии на участок нервного или мышечного волокна раздражителя достаточной силы и продолжительности. Проявлением возникшего возбуждения является быстрое колебание мембранного потенциала (см. рис. 99). При этом возбужденный участок имеет электроотрицательный заряд по отношению к невозбужденному. В потенциале действия различают местные колебания мембранного потенциала, пик потенциала действия и следовые потенциалы — отрицательный и положительный. Пик потенциала действия представляет собой кратковременное изменение внутреннего потенциала и имеет очень быструю восходящую фазу и несколько замедленный спад. Вслед за пиком потенциала действия регистрируются более слабые и продолжительные отрицательные, а затем положительные следовые потенциалы. Продолжительность потенциала действия в нервных и мышечных волокнах составляет 0,1—5,0 мс. В развитии потенциала покоя и потенциала действия роль играет система проникающих каналов для ионов Na + , К + , С1 - , Са 2+ . В нервной мембране присутствуют специфические натриевые, калиевые, хлорные и кальциевые каналы, которые пропускают только вышеназванные ионы. Эти каналы обладают воротными механизмами и могут быть открытыми и закрытыми. Определение состояния ионных каналов мембраны очень важно для образования потенциала покоя, где ведущая роль принадлежит неравномерному распределению ионов калия. В образовании потенциала действия основную роль играют ионы натрия. Проведение возбуждения — специализированная функция нервных волокон. Скорость возбуждения по ним зависит в основном от диаметра и гистологических особенностей строения нервных волокон. Чем больше диаметр нервного волокна, тем выше в нем скорость распространения возбуждения. Например, по нервному волокну диаметром 12— 22 мкм скорость распространения возбуждения составляет 70—120 м/с, а по нервному волокну диаметром 8—12 мкм — только 40—70 м/с.
По гистологическому строению нервные волокна делят на миелиновые и безмиелиновые. Миелиновое волокно состоит из осевого цилиндра и покрывающей его миелино-вой, или шванновской, оболочкой. В ее состав входят жи-ровидные вещества, которые обладают высоким удельным сопротивлением и выполняют изолирующую роль. Через равные промежутки миелиновая оболочка прерывается и оставляет открытые участки осевого цилиндра шириной около 1мкм (перехват Ранвье). Поверхность осевого цилиндра представлена плазматической мембраной, а его содержимое — аксоплазмой. Безмиелиновые волокна не имеют миелиновой оболочки, а покрыты только шванновскими клетками. Пространство между шванновскими клетками и осевым цилиндром заполнено межклеточной жидкостью, что дает возможность поверхностной мембране осевого цилиндра сообщаться с окружающей нервное волокно средой.
Передача возбуждения по миелиновым и безмиелино-вым волокнам имеет свои особенности. Так, передача потенциала действия по миелиновому волокну происходит скачкообразно от одного перехвата Ранвье к другому, что дает возможность возбуждению распространяться без угасания. Скорость распространения по миелиновым волокнам значительно выше, чем по безмиелиновым. Если скорость возбуждения по двигательным нервным волокнам (покрытым миелиновой оболочкой) составляет 80—120 м/с, то по волокнам, которые не имеют миелиновых волокон, — только 0,5—2,0 м/с. Распространение возбуждения по нервному волокну при нанесении возбуждения подчиняется соответствующим законам.
Закон физиологической целостности — проведение возбуждения по нервному волокну возможно только в том случае, если сохраняется не только его анатомическая, но и физиологическая целостность (непрерывность).
Закон двухстороннего проведения возбуждения — передача возбуждения происходит в двух направлениях — центростремительном и центробежном.
Закон изолированного проведения возбуждения — при нанесении раздражения возбуждение проводится только по одному нервному волокну и не охватывает соседние волокна, что обусловливает строгую координацию рефлекторной деятельности. Нервные волокна мало устают. Это объясняется низкими энергетическими затратами и быстрыми восстановительными процессами.
Синапс — это специализированная структура, которая обеспечивает передачу нервного импульса из нервного волокна на эффекторную клетку — мышечное волокно, нейрон или секреторную клетку.
Синапсы классифицируют по анатомо-гистологическому принципу (нейросекреторные, нервно-мышечные, межнейронные); нейрохимическому принципу (адренергические — медиатор норадреналин и холинергические — медиатор ацетилхолин); функциональному (возбуждающие и тормозные). Нервно-мышечный синапс состоит из трех основных структур: пресинаптической мембраны, синапти-ческой щели и постсинаптической мембраны. Пресинаптическая мембрана покрывает нервное окончание, а постсинаптическая — эффекторную клетку. Между ними находится синаптическая щель. Постсинаптическая мембрана отличается от пресинаптической тем, что имеет белковые хеморецепторы, чувствительные не только к медиаторам, гормонам, но и к лекарственным и токсическим веществам. Строение нервно-мышечного синапса обусловливает его физиологические свойства: 1) односторонее проведение возбуждения (от пре- к постсинаптической мембране) при наличии чувствительных к медиатору рецепторов только в постсинаптической мембране; 2) синаптическая задержка проведения возбуждения, связанная с малой скоростью диффузии медиатора в сравнении со скоростью нервного импульса; 3) низкая лабильность и высокая усталость синапса; 4) высокая избирательная чувствительность синапса к химическим веществам.
Передача возбуждения в синапсе представляет собой сложный физиологический процесс, который проходит несколько стадий: 1) синтез медиатора; 2) секреция медиатора; 3) взаимодействие медиатора с рецепторами постсинаптической мембраны; 4) инактивация (полная утрата активности) медиатора. Известно, что некоторые химические вещества, в том числе и лекарственные, могут существенно влиять на возбуждение в синапсе. Это явление нашло применение в клинической практике.
Нейромоторная единица — это анатомическая и функциональная единица скелетных мышц, которая состоит из аксона (длинного отростка мотонейрона спинного мозга) и иннервируемых им определенного количества мышечных волокон. В состав нейромоторной единицы может входить разное количество мышечных волокон (от единиц до нескольких тысяч), которое зависит от специализации мышцы. Двигательная единица работает как единое целое. Импульсы, выработанные мотонейроном, приводят в действие все образующие ее мышечные волокна.
Основной функцией скелетных мышц является сокращение, которое выражено различными движениями человека. Скелетные мышцы выполняют также рецепторную, обменную и терморегулирующую функции. Они образуются большим количеством многоядерных мышечных волокон. Сократительной частью мышечного волокна являются длинные мышечные нити — миофибриллы, которые проходят внутри волокна от одного конца к другому и имеют поперечную очерченность. Последняя образована чередованием темных (анизотропных) А-дисков и светлых (изотропных) 1-дисков (см. рис. 53). Через середину 1-диска проходит Z-линия; две соседние Z-линии ограничивают саркомер, структурно-фунциональную единицу. При электронной микроскопии волокон видно, что в составе А-диска есть более светлый участок (Н-зона), а в центре этот диск пересекает темная полоса — М-линия. Темный диск образован толстыми нитями белка миозина, а светлый 1-диск — тонкими нитями белка актина. В мышечном волокне содержатся также фибриллярный палочковидный белок — тропомиозин и глобулярный белок — тропонин. Механизм сокращения состоит в перемещении (перетягивании) тонких нитей вдоль толстых к центру саркомера за счет поперечных актиномиозиновых мостиков. Основным источником энергии, необходимой для мышечного сокращения, служат аденозинтрифосфорная кислота (АТФ) и присутствие ионов Са 2+ и Mg 2+ . Преобразование химической энергии в механическую происходит в мышце как без кислорода, так и с его участием. Анаэробная (бескислородная) фаза характеризуется рядом последовательных реакций, которые приводят к распаду АТФ и креатинфосфата, и их восстановлением. Выделенная при этом энергия используется для сокращения мышц и восстановления (ресинтеза) этих веществ. Аэробная (кислородная) фаза химических преобразований связана с процессами окисления молочной кислоты до углекислого газа и воды. Возникшая энергия используется для дальнейшего преобразования остатков молочной кислоты в глюкозу, а затем в гликоген.
Деятельность скелетной мускулатуры регулируется ЦНС — корой головного мозга, через чувствительные, двигательные и симпатические нервные волокна. Скелетные мышцы обладают следующими физиологическими свойствами: возбудимостью, проводимостью, рефрактерностью, лабильностью и сокращением. Возбудимость, скорость распространения возбуждения, лабильность мышечной ткани ниже, чем в нервной, а рефракторный период продолжительнее нервного. Скелетные мышцы могут выполнять работу в режиме изотонического, изометрического и ауксото-ничного сокращения. При первом сокращении в основном происходит укорочение мышечного волокна, но напряжение остается постоянным, а при втором — длина мышечных волокон остается неизменной, но изменяются длина и напряжение. Характер сокращения скелетных мышц зависит от частоты раздражения (частотыпоступления нервныхимпульсов).
Раздражение одиночным импульсом ведет к одиночному мышечному сокращению, а следующих один за другим нервных импульсов — к тетаническому сокращению, или тетанусу.
Физиологические свойства гладких мышц связаны с особенностью их строения, уровнем обменных процессов и значительно отличаются от особенностей скелетных мышц. Гладкие мышцы менее возбудимы, чем поперечнополоса-тые. Сокращение гладкой мускулатуры происходит медленнее и продолжительнее. Рефракторный период у гладких мышц более удлиненный, чем у скелетных (до нескольких секунд). Характерная особенность гладких мышц — их способность к автоматической деятельности, которая обеспечивается нервными элементами. Гладкие мышцы иннервируют-ся симпатическими и парасимпатическими вегетативными нервами, обладают высокой чувствительностью к некоторым биологически активным веществам (ацетилхолин, адреналин, норадреналин, серотонин и др.).
А.П.Осипов
ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Краткий курс лекций
Автор - кандидат медицинских наук, доцент кафедры инфекционных болезней Пермской ГСХА А.П. Осипов.
О-741 Осипов А.П. Основы физиологии человека.: Краткий курс лекций/ А.Осипов; ФГБОУ ВПО Пермская ГСХА. - Пермь: Изд-во ФГБОУ ВПО Пермская ГСХА, 2012. - 120 с.
В кратком курсе лекций обобщены современные сведения по физиологии человека.
Е.И. Самоделкин, УДК
Рекомендовано к изданию на заседании методической комиссии факультета ветеринарной медицины и зоотехнии Пермской государственной сельскохозяйственной академии имени академика Д.Н. Прянишникова (протокол № от 2012 г.)
ã ФГБОУ ВПО Пермская ГСХА, 2012
ВВЕДЕНИЕ
Физиология - наука о жизнедеятельности целостного организма, физиологических систем, органов, клеток и субклеточных структур. Физиология изучает механизмы регуляции и закономерности жизнедеятельности организма и взаимодействия его с окружающей средой, направленные на достижение полезного результата и обладающие приспособительными свойствами. Она использует сведения, полученные такими науками, как анатомия, гистология, биология, биохимия, экология, биофизика и многими другими. Гиппократ заложил основы для понимания роли отдельных систем и функций организма как целого. Декарт сформулировал рефлекторный принцип организации движений. Вильям Гарвей открыл кровообращение, а М. Мальпиги показал, что оно осуществляется по замкнутой сети сосудов. Ф. Мажанди доказал существование чувствительных и двигательных нервов. Клод Бернар обнаружил нервную регуляцию тонуса кровеносных сосудов и обмена углеводов, а также сформулировал представление о внутренней среде организма. Основные принципы деятельности мозга сформулировал Ч.С. Шеррингтон, капиллярное кровообращение исследовал А. Крог.
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
В основе приспособительных реакций организма лежит раздражимость - способность реагировать на воздействия изменением структуры и функций. Раздражимостью обладают все клетки животных и растений. В ходе эволюции раздражимость некоторых тканей достигла наивысшего развития и трансформировалась в возбудимость (способность отвечать на раздражение возбуждением). К возбудимым относят нервную, мышечную и секреторную ткани. Возбудимость оценивают по порогу раздражения (минимальной силе раздражителя, которая способна вызвать возбуждение). Раздражители по их природе делят на физические, химические, биологические (вирусы, бактерии и др.), адекватные и неадекватные. Адекватными называют раздражители, к восприятию которых биологическая структура специально приспособлена. Поэтому пороговая сила адекватных раздражителей наименьшая. Например, адекватным для фоторецепторов является свет, для мышц - нервный импульс. Неадекватными называют раздражители, которые действуют на структуру, не приспособленную для их восприятия. Например, скелетная мышца реагирует сокращением и на электрические раздражения.
Биоэлектрические явления в возбудимых тканях. Возбуждение - это совокупность процессов, в результате которых кратковременная деполяризация цитоплазматической мембраны вызывает специализированную реакцию клетки (проведение нервного импульса, сокращение мышцы и т.д.).
В середине 20 века. А.Ходжкин, Э.Хаксли и Б.Катц создали мембранно-ионную теорию, согласно которой МП обусловлен разными концентрациями ионов калия, натрия и хлора по обе стороны клеточной мембраны. По сравнению с внеклеточной жидкостью, цитоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора. Высокая проницаемость мембраны для калия приводит к выходу части внутриклеточного калия в окружающую клетку среду и к появлению положительного заряда на наружной поверхности мембраны. Органические анионы, для которых мембрана непроницаема, создают на внутренней поверхности мембраны отрицательный заряд, а поддерживает высокие концентрации калия в клетке и натрия вне ее натрий-калиевый насос.
Раздражение возбудимой клетки может вызвать локальный ответ или потенциал действия. Локальный ответ возникает при подпороговых раздражениях. Он находится в прямой зависимости от силы стимула, локализуется на поверхности клетки только в месте ее раздражения и увеличивает возбудимость клетки. Потенциал действия (ПД) возникает под влиянием порогового или сверхпорогового раздражений. При этом проницаемость мембраны для натрия увеличивается и в результате проникновения натрия в клетку ее мембрана заряжается положительно по отношению к наружной среде. Затем закрываются натриевые и открываются дополнительные калиевые каналы. В результате выхода калия из клетки начинается восстанавление МП (реполяризация мембраны).
В ПД различают (рис. 1.):
1. Предспайк (локальный ответ) - деполяризация мембраны до критического уровня.
2. Спайк - состоит из восходящей (деполяризация) и нисходящей (реполяризация) частей.
3. Следовой потенциал - состоит из следовой деполяризации и гиперполяризации.
Возбудимость в период предспайка повышена (фаза повышенной возбудимости) и даже слабый дополнительный стимул может вызвать формирование ПД. В период спайка мембрана не возбудима (абсолютная рефрактерность). Затем возбудимость постепенно восстанавливается (относительная рефрактерность). В это время для нового возбуждения нужно сверхпороговое раздражение. При следовой деполяризации возбудимость повышена (экзальтация), а при гиперполяризации - понижена (субнормальная возбудимость).
Законы раздражения отражают зависимость ответной реакции возбудимой ткани от силы раздражителя.
Закон силы: чем сильнее раздражение, тем больше ответная реакция. В соответствии с этим законом функционирует скелетная мышца. Она состоит из мышечных волокон с разной возбудимостью. На пороговые раздражители отвечают наиболее возбудимые волокна. Увеличение силы раздражителя дополнительно вовлекает в ответ волокна с меньшей возбудимостью и амплитуда сокращения мышцы растет.
Закон раздражения Дюбуа-Реймона: действие постоянного тока зависит от его силы и скорости нарастания. При медленном нарастании ткань приспосабливается к раздражителю (аккомодация) и возбуждение может не возникать.
Закон силы-времени отражает зависимость пороговой величины постоянного тока от времени его действия. Чем короче импульсы тока, тем меньшую раздражающую способность они имеют. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение которого ток, равный реобазе, вызывает возбуждение, называется полезным временем. Хронаксия - минимальное время, в течение которого ток, равный двум реобазам, вызывает реакцию.
Закон полярного действия постоянного тока: при замыкании тока возбуждение возникает под катодом, а при размыкании - под анодом.
Закон физиологического электротона: в области катода возбудимость повышается (катэлектротон), а у анода - снижается (анэлектротон). При длительном действии постоянного тока возбудимость под катодом снижается (катодическая депрессия), а под анодом - растет (анодная экзальтация).
Нервное волокно обладает: возбудимостью, проводимостью и лабильностью. Возбуждения распространяется по нервному волокну только при его анатомической и физиологической целостности, не переходит на соседнее нервное волокно(закон изолированного проведения), не изменяется по амплитуде (закон незатухающего или бездекрементного проведения) и проводится в обе стороны от места раздражения (закон двустороннего проведения).
Возбужденный участок поверхности нервного волокна приобретает отрицательный заряд. Так как соседний невозбужденный участок заряжен положительно, то между ними потечет электрический ток. Это вызовет возбуждение покоящегося участка и тоже изменит его заряд. В конечном итоге возбуждение распространится по всей поверхности не покрытого миелиновой оболочкой (безмякотного) нервного волокна (рис. 2а.). В миелиновых (мякотных) отростках нейронов, возбуждение может возникать только в перехватах Ранвье. Поэтому оно распространяется скачками от одного перехвата к другому (рис. 2б.) и движется гораздо быстрее, чем в безмякотных волокнах.
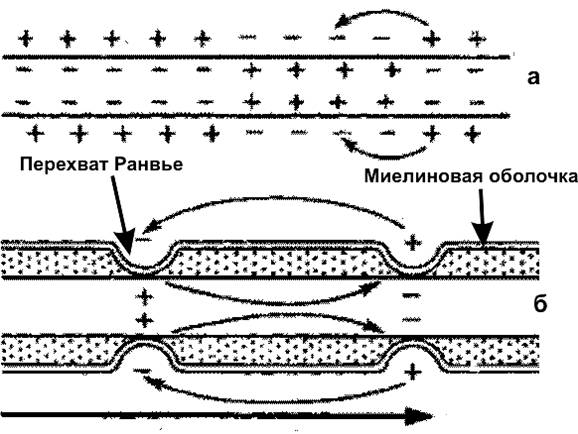 |
| Рис. 2. Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б) нервных волокнах |
По диаметру и скорости проведения возбуждения нервные волокна делят на типы А, В и С. Самые толстые волокна типа А (диаметр 12-22 мкм) с наибольшей скоростью (70-120 м/с) проводят возбуждение от мозга к скелетным мышцам и от рецепторов мышц к мозгу. От многих других рецепторов идут волокна типа А с несколько меньшими диаметром (от 8 до 1 мкм) и скоростью проведения возбуждения (5-70 м/с). К волокнам типа В относятся преганглионарные вегетативные волокна (диаметр - 1-3,5 мкм, скорость проведения возбуждения - 3-18 м/с). Только волокна типа С являются безмякотными (их диаметр 0,5-2 мкм, скорость проведения возбуждения менее 3 м/с). Они являются постганглионарными симпатическими волокнами, а также идут от болевых рецепторов, части терморецепторов и рецепторов давления.
Нервные волокна обладают лабильностью (функциональной подвижностью). Ее измеряют по максимальному количеству возбуждений, которое способно воспроизвести нервное волокно. У нервных волокон лабильность выше (до 1000 Гц) чем в других возбудимых структурах. Если нерв повредить (химическим веществом, нагреванием, охлаждением или током) не нарушая анатомическую целостность, то в нем развивается состояние парабиоза. При этом последовательно сменяются уравнительная, парадоксальная и тормозная фазы. В уравнительную фазу - при редких раздражениях все импульсы проводятся через поврежденный участок, а при высоком ритме только часть. В парадоксальную - ответная реакция на частые раздражения меньше, чем на редкие. В тормозную - нерв не проводит любые возбуждения. При выходе из парабиоза наблюдаются те же фазы, но в обратной последовательности.
Мышцы обладают возбудимостью (возбуждаются при действии раздражителей), проводимостью (проводить возбуждение) и сократимостью (способны изменять свою длину или напряжение при возбуждении). Для сердечных и части гладких мышечных волокон, дополнительно к перечисленным свойствам, характерна автоматия (способность к самопроизвольному возбуждению), а уникальным свойством гладких мышц является пластичность (долго сохраняют приданную им длину).
Сила мышцы определяется максимальным грузом, который она может поднять, а работа - произведением величины поднятого груза на высоту подъема. Максимальная работа производится при средних величинах нагрузок. При изотоническом сокращении мышцы изменяется ее длина, а напряжение постоянно (так сокращаются мышцы при отсутствии сопротивления изменению длины). При изометрическом сокращении длина мышцы постоянна, а ее напряжение растет (например, при попытке поднять чрезмерный груз). В естественных условиях наблюдаются смешанные сокращения (изменяются и длина и напряжение мышцы).
При однократном раздражении возникает одиночное мышечное сокращение. В нем выделяют: латентный период (время от начала раздражения до начала ответной реакции), фазу укорочения и фазу расслабления. Если каждый следующий стимул поступает к скелетной мышце в период ее укорочения (рис. 3) - возникает гладкий тетанус, а в фазу расслабления - зубчатый тетанус. В естественных условиях к скелетной мышце поступает такая серия импульсов, на которую мышца отвечает гладким тетанусом. Его амплитуда, как правило, выше амплитуды одиночного сокращения. Н.Е. Введенский объяснил это оптимумом и пессимумом. Оптимум - частота, при которой раздражения наносятся в фазу повышенной возбудимости (тетанус максимальный). Пессимум - частота, при которой новое раздражение наносится в фазу пониженной возбудимости (тетанус – минимален).
При изучении скелетных и сердечных мышц в поляризованном свете, видны чередующиеся зоны с разной оптической плотностью (рис. 4). Это позволило разделить сократительные элементы поперечнополосатых мышечных волокон (миофибрилл) на функциональные единицы - саркомеры (участки между соседними Z-мембранами).

Рис. 4. Микрофотография участка поперечнополосатой мышцы
Характерная оптическая плотность участков саркомера обусловлена особенностями расположения в них (рис. 5) сократительных белков (актина и миозина).

Рис. 5. Схема расположения сократительных белков в саркомере
(в Н-зоне миозиновые фибриллы не имеют мостиков)
Около Z-мембран содержится актин (светлая область – изотропный или I -диск). Ближе к середине саркомера видны темные (анизотропные) А-диски с миозином и актином. В центре саркомера проходит М-линия. По обе стороны от нее расположен миозин (просветление или Н-зона). Актин с одной стороны прикреплен к Z-мембране, а с другой заканчивается между миозиновыми волокнами в области А-диска (на границе с Н-зоной). При сокращении саркомера уменьшается ширина только изотропных дисков. Миозиновые нити при этом могут достигать свободными концами Z- линий, а длина саркомеров - укорачиваться на 30-50%.
В скелетных мышцах используется кальций из внутриклеточных цистерн саркоплазматической сети. Гладкие мышцы получают кальций только из межклеточных пространств, а сердечная мускулатура использует оба источника данного иона. Использование только внутриклеточных источников кальция позволяет скелетным мышцам сокращаться и расслабляться с наибольшей скоростью, а гладкая мускулатура - медленно изменяет свой тонус.
Синапс - функциональное соединение между нейроном и другими клетками. Существуют электрические и химические синапсы. Электрическим синапсам свойственно низкое электрическое сопротивление в области контакта клеток и ПД быстро передается на соседнюю мембрану. Химические синапсы состоят из пресинаптической мембраны, постсинаптической мембраны и синаптической щели (рис. 6.).
 |
| Рис. 6. Схема строения химического синапса |
Под влиянием нервных импульсов, в химических синапсах происходит высвобождение химического вещества - медиатора (например, ацетилхолина, норадреналина) из пресинаптических утолщений аксона в синаптическую щель и его взаимодействие с рецепторами на постсинаптической мембране. В возбуждающих синапсах это приводит к возникновению возбуждающего (ВПСП), а в тормозных - тормозного (ТПСП) постсинаптических потенциалов. После того как медиатор выполнил свою функцию, он разрушается или поглощается клетками.
За пределами ЦНС наиболее распространен медиатор ацетилхолин. Он облегчает проведение возбуждения через вегетативные ганглии, повышает секрецию адреналина надпочечниками и соляной кислоты желудочными железами, угнетает работу сердца, вызывает сокращение гладких мышц некоторых внутренних органов и экзокринных желез. В гладких мышцах бронхов, кишечника, мочевого пузыря, матки, круговой и цилиарной мышцах глаза ацетилхолин приводит, соответственно, к бронхоспазму, усилению перистальтики кишечника и желудка (при расслаблении сфинктеров), сокращению мочевого пузыря и сужению зрачка.
По расположению синапсы делят на периферические (нервно-мышечные, рецепторно-нейрональные и т.д.); центральные (аксосоматические, аксодендритные и др.); по знаку действия (возбуждающие и тормозящие) и по выделяемому медиатору (холинергические, адренергические и др.).
Химические синапсы проводят возбуждение в одном направлении; передают возбуждение медленнее, чем по нервному волокну (синаптическая задержка); имеют низкую лабильность, а также высокую утомляемость и чувствительность к химическим веществам.
Читайте также:
