
Ганглионарная нервная система характерна для
Существует несколько типов организации нервной системы, представленные у различных систематических групп животных.
С появлением ганглионарной (узловой) нервной системы (черви, моллюски, иглокожие) происходит специализация ответных реакций. Становится возможной передача возбуждения от одних узлов к другим. Структура и функция нервной системы на этом этапе эволюции находятся в прямой связи с рецепторными образованиями. Чувствительные клетки нервной системы в процессе эволюции совершенствовались параллельно с развитием аппаратов рецепции. Этому в значительной мере способствовала морфологическая близость аппаратов рецепции и чувствительных нервных клеток.
Дальнейшее совершенствование функций нервной системы, наблюдающееся у хордовых, связано с централизацией нервных узлов. В структуре нервной системы позвоночных животных развиваются специализированные синапсы, а вместе с ними и множественные связи между нервными клетками. Появление многосинаптической связи создало предпосылки для качественно новых форм взаимоотношений между системами организма, а также между организмом и средой.
У рыб хорошо развит обонятельный мозг, структурно обособлены бледный шар и нервные центры среднего мозга — красное ядро и черная субстанция. В регуляции жизнедеятельности рептилий ведущую роль приобретают большие полушария головного мозга и подкорковые ядра. У отдельных представителей этого класса появляется новая кора, достигающая совершенства у млекопитающих и высшего их представителя — человека.
| Головной мозг | Передний мозг | Конечный мозг | Обонятельный мозг, Базальные ганглии, Кора больших полушарий, Боковые желудочки |
| Промежуточный мозг | Эпиталамус, Таламус, Гипоталамус, Третий желудочек | ||
| Ствол мозга | Средний мозг | Четверохолмие, Ножки мозга, Сильвиев водопровод | |
| Ромбовидный мозг | Задний мозг | Варолиев мост, Мозжечок | |
| Продолговатый мозг | |||
| Спинной мозг |
Эволюция нервной системы тесно связано с эволюцией мышечных тканей. Клетки многоклеточных животных постепенно специализируется для выполнения различных функций. Мышечные клетки появляются в эволюции раньше, чем нервные клетки. Эти первопредки мышечных клеток находятся на поверхности тела и способны реагировать на внешние воздействия сокращением. Хлопин называл их мионейроэпителиальными клетками.
В ходе дальнейшего развития многоклеточных организмов мышечные клетки уходят в более глубокие слои тела, поэтому появляется необходимость в чувствительных клетках, доступных к поверхностной стимуляции раздражителями и способные передавать возбуждение глубже лежащим мышечным клеткам. Так появились организмы, имеющие нейроны на поверхности тела, отростки которых находятся в прямом контакте с мышечными клетками.
Следующей ступенью развития нервной системы является появление нервных цепей, сначала из 2-х нейронов, а затем и с большим количеством нейронов. Например, такие 2-х нейронные цепи имеются в каждом сегменте дождевого червя. 1-й нейрон (афферентный, чувствительный) лежит на поверхности тела, аксон 1-го нейрона передает импульс глубже лежащему 2-му нейроны (эфферентный, моторный), а 2-й нейрон вызывает сокращение мышечных клеток сегмента.
На следующем этапе появляются межсегментные нейроны у сегментированных животных. Это позволяет координировать совгласованные действия сегментов.
Увеличение числа этих соединений привело к появлению пучка, тянущегося вдоль тела близко к центральной оси, в конечном виде - спинного мозга и головного мозга.
В целом для эволюции нервной системы характерно консервативность: у высших сохраняется признаки сегментарности, присущие низшим; химическая передача импульсов в синапсах и у низших, и у высших. Чем выше уровень организации, тем выраженнее в эмбриональном периоде опережающее развитие и созревание нервной системы. Чем выше уровень организации вида, тем большее число бластомеров зародыша используется для закладки нервной системы. Так, у человека 1/3 площади поверхности оплодотворенной яйцеклетки является презумптивной зоной (будущей зоной) нервной трубки.
- Функции игровой активности животных.
Функции игровой активности по Фабри.1) Развивающая деятельность. На примере манипуляционных игр. Качественные изменения в поведении детеныша связаны с результатами манипуляционных игр, созревание моторных и сенсорных компонентов этого первичного манипулирования. (возможность взять разные предметы в рот у лисенка связаны с первичным захватыванием соска)Значение – формирование моторных компонентов определяется качественными преобразованиями в двигательной сфере , расширение функций и переход некоторых функций от ротового аппарата и наоборот (иногда и смена функций лакание – сосание). Биологическая обусловленность манипулирования – у кошек ↑мультифункциональность конечностей, у барсуков – специализация к рытью нор →↑развиты передние конечности. →игры обусловлены образом жизни ж. Но противоречивая картина у безьян - передние конечности менее специализированы и их дополнительные функции получили предельное развитие среди млекопитающих. 1.2. Ювенильное поведение и взрослое поведение Двиг.репертуар взрослого формируется путем обрастания и дополнения инстинктивной , врожденной основы поведения видотипичным инд.опытом т\е путем облигарного начения. С возрастом манипулирование приобретает все больше видотипичных черт. Значение – повышается моторно-сенсорный опыт, устанавливаются биологически значимые связи с компонентами среды. 2.Функция это формирование общения.В процессе совместной игры формируется групповое поведение.Под совместными следует понимать те игры где имеет место согласованность действия. Совместные игры без предметов.В совместных манипуляционных играх ж включают в игру какие нибудь предметы в качестве объекта игры такие игры выполняют коммуникативную роль и предметы могут служить и заменой натурального пищевого объекта.2.2. Игровая сигнализация – согласованность действий основывается на обоюдной врожденной сигнализации эти сигналы выполняют функцию ключевых стимулов игрового поведения. Это позы, движения, звуки оповещают партнера о готовности к игре.Важными являются и сигналы предотворощающие серьезный исход игры без подобного оповещения о том, что агрессия ненастоящая игра может перейти в борьбу.2.3. Значние совместных игр для взрослого поведения – если детеныша лишить игры он будет несостоятелен во взрослой жизни. Учатся половому поведению, материнскому. Молодые обезьяны учатся общаться друг с другом в игре.3. Познавательная функция игры – в ходе игры молодь приобретает информацию о свойствах и качествах предметов в окружающей его среде.Это позволяет уточнять и дополнять видовой опыт применительно к конкретным условиям жизни. Менее всего исследовательский компонент в играх служащих лишь физическими упражнениями, в большей где имеет место активное воздействие на объект т\е в манипуляционных играх. Особое мето – опосредованные игры – трофейные т\е совместное познавание предмета следует общение м\д ж.Итак в ходе онтогенезе расширяется и усложняется познавательная деят.ж , имеет место расширение функций, после выхода из гнезда поведение обращается на качественно новые объекты и можно говорить о смене функций
- Характеристика видов научения у животных по У.Торпу.
Классификация видов научения, предложенная в 1963 г. У.Торпом - описательная по историческому принципу с моментами обобщения. Торп выделяет виды научения, изучавшиеся зоопсихологами в тот или иной период развития науки зоопсихологии. Торп “считает, что у различных видов могут быть разные механизмы, ответственные за обучение; он оставляет открытым вопрос о том, в какой мере случаи однотипного обучения у представителей разных таксономичских типов обусловлены сходными механизмами.
Классификация научения по У.Торпу:
1. Привыкание (габитуация);
2. Ассоциативное научение:
а) классический условный рефлекс.
Синонимы: респондентное научение, условный рефлекс первого рода;
б) оперантный условный рефлекс.
Синонимы:“пробы и ошибки”, условный рефлекс второго рода,
инструментальное научение, научение по Скиннеру;
3. Латентное (скрытое) научение;
4. Инсайт (озарение):
а) собственно инсайт (“улавливание отношений”);
б) подражание типа социального облегчения;
в) истинное подражание (“копирование поведенческих актов”);
5. Импринтинг (запечатление):
а) запечатление привязанности;
б) половой импринтинг.
Торп выделяет научение неассоциативное и ассоциативное.
К неассоцативному относят привыкание, характерное для всех животных, от одноклеточных до человека. При ассоциативном научении образуется ассоциативная связь между двумя психическими явлениями.
Научение: Имитационное научение
Облигатное имитационное научение
Факультативное имитационное научение
Невидотипичное имитационное манипулирование
Имитационное решение задач
Латентное, или скрытое, научение исследовал и пытался объяснять Толмен, наблюдая за крысами в лабиринте. В основе этого вида научения лежит исследовательская мотивация. В ходе исследовательского поведения строится то, что Толмен назвал когнитивной картой. У животного формируется психический образ компонентов среды и собственных действий в среде. После этого животное может переходить к нормальной повседневной жизни. Кроме этих ситуаций, латентное научение происходит у детенышей зверей и детей в процессе игры.
Инсайт — высшая форма научения, основывается на опыте, полученном раньше при других сходных обстоятельствах. Присущ только птицам и млекопитающим, обладающим интеллектом. Оказавшись в проблемной ситуации, животное остается неподвижным и только оценивает обстановку, не совершая никаких действий, после чего начинает действовать с учетом реально существующих связей между компонентами среды.
- Характеристика облигатного и факультативного научения.
Ганглии нервной системы — скопление нейронов и глии, находящиеся за пределами головного и спинного мозга.
Подобные образования в центральной нервной системе называют ядрами. Они выступают связующими звеньями структур нервной системы, осуществляют первичную обработку импульсов, отвечают за функции висцеральных органов.
Человеческий организм осуществляет два типа функций – соматические и вегетативные. Соматические подразумевают восприятие внешних раздражителей и реакцию на них при помощи скелетной мускулатуры. Эти реакции могут контролироваться сознанием человека, а за их выполнение отвечает центральная нервная система.
Вегетативные функции – пищеварение, метаболизм, кроветворение, кровообращение, дыхание, потоотделение и другое, контролирует вегетативная система, которая не зависит от человеческого сознания. Кроме регуляции работы висцеральных органов, вегетативная система обеспечивает трофику мускулатуры и ЦНС.
Ганглии, отвечающие за соматические функции, представляют спинномозговые узлы и узлы черепных нервов. Вегетативные, в зависимости от расположения центров в ЦНС, делят на: парасимпатические и симпатические.
Первые находятся в стенках органа, а симпатические дистанционно – в структуре, называемой пограничным стволом.
Строение ганглия
В зависимости от морфологических особенностей, размер ганглиев колеблется от нескольких микрометров до нескольких сантиметров. По сути это скопление нервных и глиальных клеток, покрытое соединительной оболочкой.
Соединительнотканный элемент пронизан лимфатическими и кровеносными сосудами. Каждый нейроцит (или группа нейроцитов) окружен капсульной оболочкой, выстланной изнутри эндотелием, а снаружи волокнами соединительной ткани. Внутри капсулы располагается нервная клетка и глиальные структуры, которые обеспечивают жизнедеятельность нейрона.
От нейрона отходит один аксон, покрытый миелиновой оболочкой, который разветвляется на две части. Одна из них входит в состав периферического нерва и образует рецептор, а вторая направляется в ЦНС.
Вегетативные центры находятся в стволе головного мозга и спинном мозге. Парасимпатические центры локализуются в краниальном и сакральном отделах, а симпатические – в тораколюмбальном.
Ганглии вегетативной нервной системы
Симпатическая система включает два типа узлов: вертебральные и превертебральные.
Вертебральные располагаются с двух сторон от позвоночного столба, образуя пограничные стволы. Они связаны со спинным мозгом с помощью нервных волокон, которые дают начало белым и серым соединительным веткам. Выходящие из узла нервные волокна направляются к висцеральным органам.
Превертебральные расположены на большем расстоянии от позвоночника, при этом также дистанционно они находятся от органов, за которые они ответственны. Примером превертебральных узлов служат шейные, брыжеечные скопления нейронов, солнечное сплетение.
Парасимпаческий отдел образован ганглиями, расположенными на органах, или в непосредственной к ним близости.
Внутриорганные нервные сплетения размещены на органе или в его стенке. Крупные внутриорганные сплетения расположены в сердечной мышце, в мышечном слое стенки кишечника, в паренхиме железистых органов.
Ганглии вегетативной и центральной нервной системы имеют свойства:
- проведение сигнала в одну сторону;
- входящие в узел волокна перекрывают зоны влияния друг друга;
- пространственная суммация (сумма импульсов способна породить потенциал в нейроците);
- окклюзия (стимуляция нервов вызывает малый ответ, чем стимуляция каждого в отдельности).
Синоптическая задержка в вегетативных ганглиях больше, чем в аналогичных структурах ЦНС, а постсинаптический потенциал продолжительный. Волну возбуждения в ганглионарных нейроцитах сменяет депрессия. Эти факторы приводят к относительно низкому ритму импульсов, сравнительно с ЦНС.
Какие функции выполняют ганглии
Основным назначением вегетативных узлов является распределение и передача нервных импульсов, а также генерация локальных рефлексов. Каждый ганглий, зависимости от расположения и особенностей трофики, отвечает за функции определенного участка тела.
Ганглиям присуща автономность от ЦНС, что позволяет им регулировать деятельность органов без участия головного и спинного мозга.
В структуре внутристеночных узлов содержатся клетки – водители ритма, способные задавать периодичность сокращений гладкой мускулатуры кишечника.
Особенность связана с прерыванием, направляющимся к внутренним органам, волокон ЦНС на периферических узлах вегетативной системы, где они образуют синапсы. При этом непосредственное влияние на внутренний орган оказывают аксоны, выходящие из ганглия.
Каждое нервное волокно, приходящее в симпатический ганглий, осуществляет иннервацию до тридцати постганглионарных нейроцитов. Это дает возможность умножать сигнал и распространять импульс возбуждения, выходящий из нервного узла.
В парасимпатические узлах одно волокно осуществляет иннервацию не более четырех нейроцитов, а передача импульса осуществляется локально.
Ганглии – рефлекторные центры
Ганглии нервной системы принимают участие в рефлекторной дуге, что позволяет корректировать деятельность органов и тканей без участия мозга. В конце девятнадцатого века русский гистолог Догель в результате опытов по изучению нервных сплетений в желудочно-кишечном тракте, выявил три типа нейронов – моторные, вставочные и рецепторные, а также синапсы между ними.
Наличие рецепторных нервных клеток подтверждает и возможность трансплантации сердечной мышцы от донора реципиенту. Если бы регуляция сердечного ритма осуществлялась через ЦНС, после пересадки сердца нервные клетки подверглись дегенерации. Нейроны и синапсы в пересаженном органе продолжают функционировать, что свидетельствует об их автономности.
В конце двадцатого века экспериментальным путем установлены механизмы периферических рефлексов, которые делают превертебральные и интрамуральные вегетативные узлы. Возможность создавать рефлекторную дугу свойственна некоторым узлам.
Местные рефлексы позволяют разгрузить центральную нервную систему, делают регуляцию важных функций более надежность, способны продолжать автономную работу внутренних органов в случае прерывания связи с ЦНС.
Вегетативные узлы получают и обрабатывают информацию о работе органов, после чего направляют ее в мозг. Это вызывает рефлекторную дугу как в вегетативной, так и в соматической системе, что запускает не только рефлексы, но и сознательные поведенческие реакции.
Центральная нервная система человека осуществляет контроль над деятельностью его организма и разделяется на несколько отделов. Головной мозг посылает и получает сигналы из организма и после их обработки имеет информацию о процессах. Нервная система разделяется на вегетативную и соматическую нервную системы.
Отличия вегетативной и соматической нервной системы
Соматическая нервная система регулируется сознанием человека и может управлять деятельностью скелетной мускулатуры. Все компоненты реакции человека на внешние факторы находятся под контролем полушарий мозга. Она обеспечивает сенсорные и моторные реакции человека, контролируя их возбуждение и торможение.
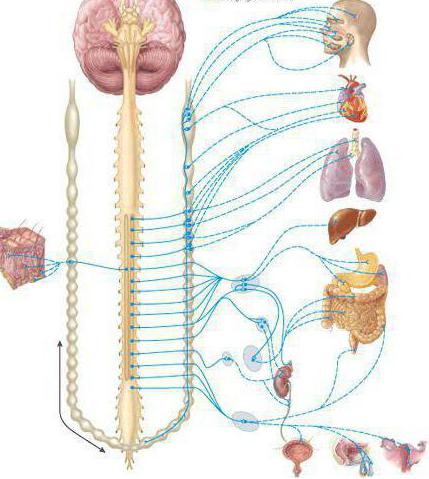
Вегетативная нервная система контролирует периферическую деятельность организма и не контролируется сознанием. Для нее характерны автономность и генерализованность воздействия на организм при полном отсутствии сознания. Эфферентная иннервация внутренних органов позволяет ей контролировать обменные процессы в организме и осуществлять обеспечение трофических процессов скелетной мускулатуры, рецепторов, кожи и внутренних органов.
Строение вегетативной системы
Работа вегетативной нервной системы контролируется гипоталамусом, который находится в центральной нервной системе. Вегетативная нервная система имеет метасегментарное строение. Ее центры находятся в головном, спинном мозге и коре головного мозга. Периферические отделы образованы стволами, ганглиями, сплетениями.
В вегетативной нервной системе различают:
- Симпатическую. Ее центр расположен в грудопоясничном отделе спинного мозга. Для нее характерны паравертебральные и предвертебральные ганглии ВНС.
- Парасимпатическую. Ее центры сосредоточены в среднем и продолговатом мозге, крестцовом отделе спинного мозга. Нервные узлы в основном интрамуральные.
- Метасимпатическую. Иннервирует желудочно-кишечный тракт, сосуды, внутренние органы организма.
В состав ее входит:
- Ядра нервных центров, расположенных в головном и спинном мозге.
- Вегетативные ганглии, которые расположены по периферии.
- Нервные волокна.
Рефлекторная дуга автономной нервной системы
Рефлекторная дуга вегетативной нервной системы состоит из трех звеньев:
- чувствительное или афферентное;
- вставочное или ассоциативное;
- эффекторное.

Чувствительное звено
Чувствительное звено расположено в спинномозговом ганглии. Этот ганглий имеет нервные клетки, сформированные группами, и их контроль осуществляется ядрами центрального головного мозга, большими полушариями и их структурами.
Чувствительное звено представлено частично униполярными клетками, которые имеют один приносящий или уносящий аксон, и они принадлежат спинальным или черепно-мозговым узлам. А также узлами блуждающих нервов, имеющих строение, похожее на спинальные клетки. В это звено входят клетки Догеля II типа, которые являются компонентами вегетативных ганглиев.
Вставочное звено
Вставочное звено в вегетативной нервной системе служит для передачи через низшие нервные центры, которыми являются вегетативные ганглии, и осуществляется это через синапсы. Расположено оно в боковых рогах спинного мозга. Нет прямой связи от афферентного звена на преганглионарные нейроны для их связи, существует кратчайший путь от афферентного нейрона до ассоциативного и от него до преганглионарного нейрона. Передача сигналов и нервных импульсов от афферентных нейронов в различных центрах осуществляется с различным количеством вставочных нейронов.
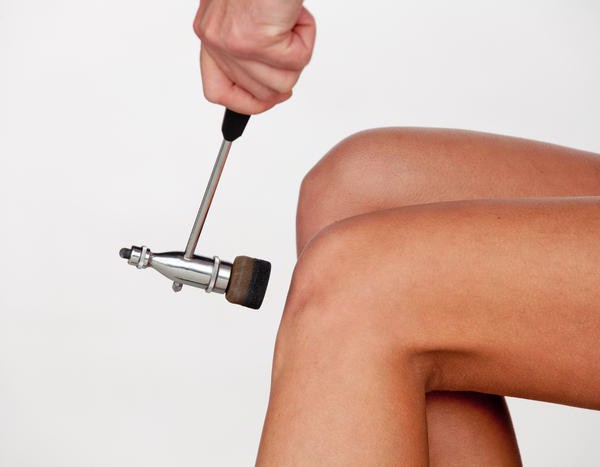
Например, в дуге спинального автономного рефлекса между чувствительным и эффекторным звеном существует три синапса, два из которых расположены в спинном мозге, а один в вегетативном узле, в котором расположен эфферентный нейрон.
Эфферентное звено
Эфферентное звено представлено эффекторными нейронами, которые расположены в вегетативных узлах. Их аксоны образуют безмиелиновые волокна, которые в составе со смешанными нервными волокнами иннервируют внутренние органы.
Вегетативные рефлекторные дуги расположены в боковых рогах.
Строение нервного узла
Ганглий – это скопление нервных клеток, которые имеют вид узелковых расширений толщиной около 10 мм. По своему строению вегетативный ганглий сверху покрыт соединительнотканной капсулой, которая образует строму из рыхлой соединительной ткани внутри органов. Мультиполярные нейроны, которые строятся из округлого ядра и крупных ядрышек, состоят из одного эфферентного нейрона и нескольких расходящихся афферентных нейронов. Эти клетки относятся по своему типу к клеткам головного мозга и являются двигательными. Их окружает неплотная оболочка – мантийная глия, которая создает постоянную среду для нервной ткани и обеспечивает полноценное функционирование нервных клеток.
Вегетативный ганглий имеет диффузное расположение нервных клеток и множество отростков, дендритов и аксонов.
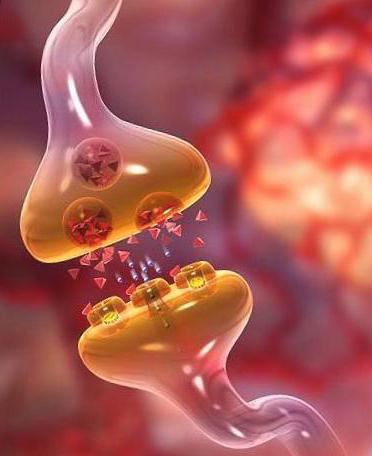
Спинномозговой ганглий имеет нервные клетки, которые расположены группами, и их расположение имеет порядок обусловленный.
Вегетативные нервные ганглии разделяются на:
- Сенсорные нейроны, которые расположены близко к спинному или центральному отделу мозга. Униполярные нейроны, из которых состоит этот ганглий, представляют собой приносящий или уносящий отросток. Они служат для афферентной передачи импульсов, и их нейроны образуют бифуркацию при ветвлении отростков. Эти отростки передают информацию от периферии к центральному афферентному нейрону – это периферический отросток, центральный – от тела нейрона в мозговой центр.
- Моторные, двигательные состоят из эфферентных нейронов, и в зависимости от их положения их называют паравертебральные, предвертебральные.
Симпатические ганглии
Паравертебральные цепочки ганглиев расположены вдоль позвоночного столба в симпатических стволах, которые идут длинной вереницей от основания черепа до копчика.
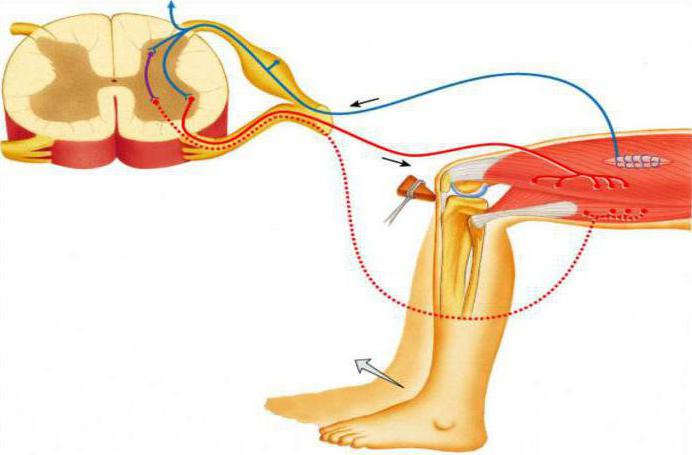
Предвертебральные нервные сплетения находятся ближе к внутренним органам, и их локализация сосредоточена перед аортой. Они образуют брюшное сплетение, которое состоит из солнечного, нижне- и верхнебрыжеечного сплетений. Они представлены двигательными адренергическими и тормозящими действие холинергическими нейронами. Также связь между нейронами осуществляется преганглионарными и постганглионарными нейронами, которые используют медиаторы ацетилхолин и норадреналин.
Интрамуральные нервные узлы имеют нейроны трех типов. Их описание было сделано русским ученым Догелем А.С., который, исследуя гистологию нейронов вегетативной нервной системы, выделил такие нейроны, как длинноаксонные эфферентные клетки первого типа, равноотросчатые афферентные клетки второго типа и ассоциативные клетки третьего типа.
Рецепторы ганглиев
Афферентные нейроны выполняют узкоспециализированую функцию, и их роль заключается в восприятии раздражителей. Такими рецепторами являются механорецепторы (реакция на растяжение или давление), фоторецепторы, терморецепторы, хеморецепторы (отвечают за реакции в организме, химические связи), ноцицепторы (реакция организма на болевые раздражители – это повреждение кожи и другие).
В симпатических стволах эти рецепторы по рефлекторной дуге передают в центральную нервную систему информацию, которая служит сигналом о повреждениях или нарушениях в организме, а также нормальной его работе.
Функции ганглиев
Каждый ганглий имеет свое местоположение, кровоснабжение, и его функции определяются этими параметрами. Спинномозговой ганглий, имеющий иннервацию из ядер головного мозга, обеспечивает непосредственную связь процессов в организме через рефлекторную дугу. От этих структурных компонентов спинного мозга иннервируются железы, гладкая мускулатура мышц внутренних органов. Сигналы, поступающие по рефлекторной дуге, идут медленнее, чем в ЦНС, и они полностью регулируются вегетативной системой, также она обладает трофической, сосудодвигательной функцией.
ГАНГЛИИ (ganglia нервные узлы)— скопления нервных клеток, окруженные соединительной тканью и клетками глии, расположенные по ходу периферических нервов.
Различают Г. вегетативной и соматической нервной системы. Г. вегетативной нервной системы делятся на симпатические и парасимпатические и содержат тела постганглионарных нейронов. Г. соматической нервной системы представлены спинномозговыми узлами и Г. чувствительных и смешаных черепных нервов, содержащих тела чувствительных нейронов и дающих начало чувствительным порциям спинномозговых и черепных нервов.
Содержание
- 1 Эмбриология
- 2 Анатомия
- 3 Гистология
- 4 Физиология
- 5 Патология
- 6 Краткая анатомическая характеристика нервных ганглиев (узлов)
Эмбриология
Зачатком спинномозговых и вегетативных узлов является ганглиозная пластинка. Она образуется у эмбриона в тех отделах нервной трубки, которые граничат с эктодермой. У зародыша человека на 14—16-й день развития ганглиозная пластинка располагается по дорсальной поверхности замкнувшейся нервной трубки. Затем она расщепляется по всей длине, обе ее половины перемещаются вентрально и в виде нервных валиков лежат между нервной трубкой и поверхностной эктодермой. В дальнейшем соответственно сегментам дорсальной стороны зародыша в нервных валиках возникают очаги пролиферации клеточных элементов; эти участки утолщаются, обособляются и превращаются в спинномозговые узлы. Из ганглиозной пластинки развиваются также чувствительные ганглии У, VII—X пар черепных нервов, аналогичные спинномозговым ганглиям. Зародышевые нервные клетки, нейробласты, формирующие спинномозговые ганглии, представляют собой биполярные клетки, т. е. имеют два отростка, отходящих от противоположных полюсов клетки. Биполярная форма чувствительных нейронов у взрослых млекопитающих и человека сохраняется только в чувствительных клетках преддверно-улит кового нерва, преддверного и спирального ганглиев. В остальных, как спинномозговых, так и черепных чувствительных узлах, отростки биполярных нервных клеток в процессе их роста и развития сближаются и сливаются в большинстве случаев в один общий отросток (processus communis). По этому признаку чувствительные нейроциты (нейроны) называют псевдоуниполярными (neurocytus pseudounipolaris), реже протонейронами, подчеркивая древность их происхождения. Спинномозговые узлы и узлы в. н. с. отличаются характером развития и строения нейронов. Развитие и морфология вегетативных ганглиев — см. Вегетативная нервная система.
Анатомия
Основные сведения об анатомии Г. приведены в таблице.
Гистология


Возраст и перенесенные заболевания сказываются на структуре спинномозговых ганглиев — в них появляется значительно большее, чем у здоровых, количество различных атипичных нейронов, в особенности с дополнительными отростками, снабженными колбовидными утолщениями, как, напр., при ревматическом пороке сердца (рис. 2), стенокардии и др. Клинические наблюдения, а также экспериментальные исследования на животных показали, что чувствительные нейроны спинномозговых узлов реагируют значительно быстрее интенсивным ростом дополнительных отростков на различные эндогенные и экзогенные вредности, нежели моторные соматические или вегетативные нейроны. Эта способность чувствительных нейронов иногда выражена значительно. В случаях хрон, раздражений вновь образованные отростки могут обвиваться (в виде намотки) вокруг тела собственного или соседнего нейрона, напоминая кокон. Чувствительные нейроны спинномозговых узлов, как и другие типы нервных клеток, имеют ядро, различные органеллы и включения в цитоплазме (см. Нервная клетка). Т. о., отличительным свойством чувствительных нейронов спинномозговых и узлов черепных нервов является их яркая морфол, реактивность, выражающаяся в изменчивости их структурных компонентов. Это обеспечено высоким уровнем синтеза белков и различных активных веществ и свидетельствует об их функциональной подвижности.
Физиология
У беспозвоночных Г. играют ту же роль, что и ц. н. с. у позвоночных, являясь высшими центрами координации соматических и вегетативных функций. В эволюционном ряду от червей до головоногих моллюсков и членистоногих Г., перерабатывающие всю информацию о состоянии окружающей и внутренней среды, достигают высокой степени организации. Это обстоятельство, а также простота анатомической препаровки, относительно большие размеры тел нервных клеток, возможность введения в сому нейронов под прямым визуальным контролем одновременно нескольких микроэлектродов сделали Г. беспозвоночных общераспространенным объектом нейрофизиол, экспериментов. На нейронах круглых червей, октапод, декапод, брюхоногих и головоногих моллюсков методами электрофореза, прямого измерения активности ионов и фиксации напряжения проводят исследования механизмов генерации потенциалов и процесса синаптической передачи возбуждения и торможения, зачастую невыполнимые на большинстве нейронов млекопитающих. Несмотря на эволюционные различия, основные электрофизиол, константы и нейрофизиол, механизмы работы нейронов во многом одинаковы у беспозвоночных и высших позвоночных животных. Поэтому исследования Г., беспозвоночных имеют общефизиол. значение.
У позвоночных соматосенсорные черепные и спинномозговые Г. функционально однотипны. Они содержат тела и проксимальные части отростков афферентных нейронов, передающих импульсацию от периферических рецепторов в ц. н. с. В сомато-сенсорных Г. нет синаптических переключений, эфферентных нейронов и волокон. Так, нейроны спинномозговых Г. у жабы характеризуются следующими основными электрофизиол, параметрами: специфическим сопротивлением — 2,25 кОм/см 2 для деполяризующего и 4,03 кОм/см 2 для гиперполяризующего тока и удельной емкостью 1,07 мкФ/см 2 . Полное входное сопротивление нейронов сомато-сенсорных Г. существенно ниже, чем соответствующий параметр аксонов, поэтому при высокочастотной афферентной импульсации (до 100 импульсов в 1 сек.) проведение возбуждения может блокироваться на уровне тела клетки. В этом случае потенциалы действия, хотя и не регистрируются от тела клетки, продолжают проводиться с периферического нерва в задний корешок и сохраняются даже после экстирпации тел нервных клеток при условии интактности Т-образных ветвлений аксонов. Следовательно, возбуждение сомы нейронов сомато-сенсорных Г. для передачи импульсов от периферических рецепторов к спинному мозгу не обязательно. Эта особенность впервые появляется в эволюционном ряду у бесхвостых амфибий.
Вегетативные Г. позвоночных в функциональном плане принято разделять на симпатические и парасимпатические. Во всех вегетативных Г. происходит синаптическое переключение с преганглионарных волокон на постганглионарные нейроны. В подавляющем большинстве случаев синаптическая передача осуществляется хим. путем с помощью ацетилхолина (см. Медиаторы). В парасимпатическом цилиарном Г. птиц обнаружена электрическая передача импульсов при помощи так наз. потенциалов соединения, или потенциалов связи. Электрическая передача возбуждения через один и тот же синапс возможна в двух направлениях; в процессе онтогенеза она формируется позже химической. Функциональное значение электрической передачи пока не ясно. В симпатических Г. амфибий выявлено небольшое количество синапсов с хим. передачей нехолинергической природы. В ответ на сильное одиночное раздражение преганглионарных волокон симпатического Г. в постганглионарном нерве прежде всего возникает ранняя отрицательная волна (O-волна), обусловленная возбуждающими постсинаптическими потенциалами (ВПСП) при активации н-холинорецепторов постганглионарных нейронов. Тормозной постсинаптический потенциал (ТПСП), возникающий в постганглионарных нейронах под действием катехоламинов, выделяемых хромаффинными клетками в ответ на активацию их м-холинорецепторов, формирует следующую за 0-волной положительную волну (П-волна). Поздняя отрицательная волна (ПО-волна) отражает ВПСП постганглионарных нейронов при активации их м-холинорецепторов. Завершает процесс длительная поздняя отрицательная волна (ДПО-волна), возникающая вследствие суммации ВПСП нехолинергической природы в постганглионарных нейронах. В нормальных условиях на высоте О-волны при достижении ВПСП величины 8—25 мв возникает распространяющийся потенциал возбуждения с амплитудой 55—96 мв, длительностью 1,5—3,0 мсек, сопровождаемый волной следовой гиперполяризации. Последняя существенно маскирует волны П и ПО. На высоте следовой гиперполяризации возбудимость снижается (период рефрактерности), поэтому обычно частота разрядов постганглионарных нейронов не превышает 20—30 импульсов в 1 сек. По основным электрофизиол. характеристикам нейроны вегетативных Г. идентичны большинству нейронов ц. н. с. Нейрофизиол. особенностью нейронов вегетативных Г. является отсутствие истинной спонтанной активности при деафферентации. Среди пре- и постганглионарных нейронов преобладают нейроны групп В и С по классификации Гассера — Эрлангера, основанной на электрофизиол, характеристиках нервных волокон (см.). Преганглионарные волокна обширно ветвятся, поэтому раздражение одной преганглионарной ветви приводит к возникновению ВПСП во многих нейронах нескольких Г. (феномен мультипликации). В свою очередь на каждом постганглионарном нейроне оканчиваются терминали многих преганглионарных нейронов, отличающихся по порогу раздражения и скорости проведения (феномен конвергенции). Условно мерой конвергенции можно считать отношение количества постганглионарных нейронов к количеству преганглионарных нервных волокон. Во всех вегетативных Г. оно больше единицы (за исключением цилиарного ганглия птиц). В эволюционном ряду это отношение возрастает, достигая в симпатических Г. человека величины 100:1. Мультипликация и конвергенция, обеспечивающие пространственную суммации) нервных импульсов, в сочетании с временной суммацией являются основой интегрирующей функции Г. при обработке центробежной и периферической импульсации. Через все вегетативные Г. проходят афферентные пути, тела нейронов которых лежат в спинномозговых Г. Для нижнего брыжеечного Г., чревного сплетения и некоторых интрамуральных парасимпатических Г. доказано существование истинных периферических рефлексов. Афферентные волокна, проводящие возбуждение с малой скоростью (ок. 0,3 м/сек), входят в Г. в составе постганглионарных нервов и оканчиваются на постганглионарных нейронах. В вегетативных Г. обнаружены окончания афферентных волокон. Последние информируют ц. н. с. о происходящих в Г. функционально-хим. изменениях.
Патология
В клин, практике наиболее часто встречается ганглионит (см.), называемый также симпато-ганглионитом, — заболевание, связанное с поражением ганглиев симпатического ствола. Поражение нескольких узлов определяют как полиганглионит, или трунцит (см.).
Спинальные ганглии нередко вовлекаются в патологический процесс при радикулитах (см.).
Читайте также:
