
Ганглиях их функциональная характеристика
Вегетативные ганглии образованы скоплением нервных клеток, среди которых можно выделить следующие типы (клеточные типы в вегетативных узлах впервые были описаны А.С. Догелем):
Ø постганглионарные симпатические или парасимпатические нейроны, аксоны которых вступают в синаптический контакт со структурами иннервируемого органа (т.е. в функциональном плане они являются собственно эфферентными нейронами узла)
Ø чувствительные (афферентные) нейроны, дендриты которых образуют рецепторы в самом иннервируемом органе, а аксоны могут переключаться как на собственные клетки узла, так и отдавать коллатерали в центральную нервную систему, доставляя туда чувствительную информацию о функциональном состоянии органа. Этот клеточный тип преимущественно характерен для парасимпатических узлов, а также для некоторых симпатических (в основном превертебральных)
Ø вставочные нейроны, тела и отростки которых не покидают вегетативного узла, обеспечивают передачу информации от чувствительных в конечном итоге на эфферентные (постганглионарные вегетативные) нейроны. Подобно чувствительным нейронам узла данный клеточный тип преимущественно характерен для парасимпатических узлов, а также для некоторых симпатических (в основном превертебральных).
Преганглионарные нервные волокна, приносящие эфферентную информацию из ц.н.с. к вегетативному узлу, как правило, многократно разветвляясь, вступают в синаптический контакт с постганглионарными нейронами узла. Причем на уровне вегетативных узлов происходит значительное расширение зоны влияния преганглионарных волокон, поскольку количество постганглионарных нейронов в узле гораздо больше (в верхнем шейном симпатическом узле в 32раза, а в ресничном ганглии в 2раза), чем приходящих к нему преганглионарных нервных волокон. Более того, на каждом постганглионарном нейроне обычно образует синапсы ветвь не одного преганглионарного волокна, а множества различных волокон, что делает возможным пространственную суммацию нервных влияний, приходящих к постганглионарным нейронам из ц.н.с.
Вегетативные ганглии обладают теми жесвойствами, что инервные центры:
Ø способностью к пространственной и временной суммации поступающей к ним информации
Ø односторонним проведением возбуждения (благодаря наличию синапсов химического типа между пре- и постганглионарными нейронами)
Ø некоторой инерцией проведения возбуждения (в связи с синаптической задержкой, которая является более длительной (1,5-30мс), чем в синапсах ц.н.с (0,3-0,5мс))
Ø 
трансформацией ритма возбуждений (в связи с тем, что постганглионарные нейроны, как правило, характеризуются меньшей лабильностью по сравнению с преганглионарными).
Рис. 5. Схема симпатической и парсимпатической иннервации гладкой мускулатуры желудка
Наличие в большинстве парасимпатических вегетативных узлов трех функциональных типов клеток (чувствительных, вставочных и собственно эфферентных нейронов) делает возможным замыкание на их уровне периферических (коротких) рефлекторных дуг, обеспечивающих осуществление разнообразных механизмов местной регуляции функции органа в зависимости от его исходной (базальной) функциональной активности. Примером таких периферических рефлексов может служить ослабление сердечной деятельности (уменьшение частоты и силы сердечных сокращений), возникающее на фоне повышенного артериального давления, несмотря на довольно большой объем крови, возвращающейся к сердцу в момент диастолы (тогда как само по себе увеличение венозного возврата крови к сердцу в момент диастолы должно приводить к повышению силы сокращения при последующей систоле).
Таким образом, постганглионарные преимущественно парасимпатические нейроны являются общим конечным путем как для эфферентной информации, которая поступает из ц.н.с., так и для чувствительной информации об уровне функциональной активности органа, воспринимаемой афферентными нейронами самого узла. В результате этого деятельность большинства внутренних органов в каждый данный момент лишь отчасти зависит от характера влияний центральной нервной системы на них. Более того, эти центральные влияния могут модифицироваться на уровне самих парасимпатических ганглиев чувствительной информацией, поступающей от самого органа. Наличие местных механизмов нервной регуляции функций внутренних органов, с одной стороны, освобождает ц.н.с. от переработки избыточной информации, а с другой – увеличивает надежность и точность регуляции физиологических функций.
Центральная нервная система человека осуществляет контроль над деятельностью его организма и разделяется на несколько отделов. Головной мозг посылает и получает сигналы из организма и после их обработки имеет информацию о процессах. Нервная система разделяется на вегетативную и соматическую нервную системы.
Отличия вегетативной и соматической нервной системы
Соматическая нервная система регулируется сознанием человека и может управлять деятельностью скелетной мускулатуры. Все компоненты реакции человека на внешние факторы находятся под контролем полушарий мозга. Она обеспечивает сенсорные и моторные реакции человека, контролируя их возбуждение и торможение.
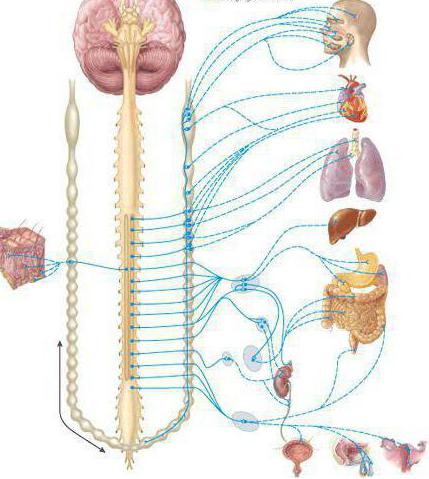
Вегетативная нервная система контролирует периферическую деятельность организма и не контролируется сознанием. Для нее характерны автономность и генерализованность воздействия на организм при полном отсутствии сознания. Эфферентная иннервация внутренних органов позволяет ей контролировать обменные процессы в организме и осуществлять обеспечение трофических процессов скелетной мускулатуры, рецепторов, кожи и внутренних органов.
Строение вегетативной системы
Работа вегетативной нервной системы контролируется гипоталамусом, который находится в центральной нервной системе. Вегетативная нервная система имеет метасегментарное строение. Ее центры находятся в головном, спинном мозге и коре головного мозга. Периферические отделы образованы стволами, ганглиями, сплетениями.
В вегетативной нервной системе различают:
- Симпатическую. Ее центр расположен в грудопоясничном отделе спинного мозга. Для нее характерны паравертебральные и предвертебральные ганглии ВНС.
- Парасимпатическую. Ее центры сосредоточены в среднем и продолговатом мозге, крестцовом отделе спинного мозга. Нервные узлы в основном интрамуральные.
- Метасимпатическую. Иннервирует желудочно-кишечный тракт, сосуды, внутренние органы организма.
В состав ее входит:
- Ядра нервных центров, расположенных в головном и спинном мозге.
- Вегетативные ганглии, которые расположены по периферии.
- Нервные волокна.
Рефлекторная дуга автономной нервной системы
Рефлекторная дуга вегетативной нервной системы состоит из трех звеньев:
- чувствительное или афферентное;
- вставочное или ассоциативное;
- эффекторное.

Чувствительное звено
Чувствительное звено расположено в спинномозговом ганглии. Этот ганглий имеет нервные клетки, сформированные группами, и их контроль осуществляется ядрами центрального головного мозга, большими полушариями и их структурами.
Чувствительное звено представлено частично униполярными клетками, которые имеют один приносящий или уносящий аксон, и они принадлежат спинальным или черепно-мозговым узлам. А также узлами блуждающих нервов, имеющих строение, похожее на спинальные клетки. В это звено входят клетки Догеля II типа, которые являются компонентами вегетативных ганглиев.
Вставочное звено
Вставочное звено в вегетативной нервной системе служит для передачи через низшие нервные центры, которыми являются вегетативные ганглии, и осуществляется это через синапсы. Расположено оно в боковых рогах спинного мозга. Нет прямой связи от афферентного звена на преганглионарные нейроны для их связи, существует кратчайший путь от афферентного нейрона до ассоциативного и от него до преганглионарного нейрона. Передача сигналов и нервных импульсов от афферентных нейронов в различных центрах осуществляется с различным количеством вставочных нейронов.
Например, в дуге спинального автономного рефлекса между чувствительным и эффекторным звеном существует три синапса, два из которых расположены в спинном мозге, а один в вегетативном узле, в котором расположен эфферентный нейрон.
Эфферентное звено
Эфферентное звено представлено эффекторными нейронами, которые расположены в вегетативных узлах. Их аксоны образуют безмиелиновые волокна, которые в составе со смешанными нервными волокнами иннервируют внутренние органы.
Вегетативные рефлекторные дуги расположены в боковых рогах.
Строение нервного узла
Ганглий – это скопление нервных клеток, которые имеют вид узелковых расширений толщиной около 10 мм. По своему строению вегетативный ганглий сверху покрыт соединительнотканной капсулой, которая образует строму из рыхлой соединительной ткани внутри органов. Мультиполярные нейроны, которые строятся из округлого ядра и крупных ядрышек, состоят из одного эфферентного нейрона и нескольких расходящихся афферентных нейронов. Эти клетки относятся по своему типу к клеткам головного мозга и являются двигательными. Их окружает неплотная оболочка – мантийная глия, которая создает постоянную среду для нервной ткани и обеспечивает полноценное функционирование нервных клеток.
Вегетативный ганглий имеет диффузное расположение нервных клеток и множество отростков, дендритов и аксонов.
Спинномозговой ганглий имеет нервные клетки, которые расположены группами, и их расположение имеет порядок обусловленный.
Вегетативные нервные ганглии разделяются на:
- Сенсорные нейроны, которые расположены близко к спинному или центральному отделу мозга. Униполярные нейроны, из которых состоит этот ганглий, представляют собой приносящий или уносящий отросток. Они служат для афферентной передачи импульсов, и их нейроны образуют бифуркацию при ветвлении отростков. Эти отростки передают информацию от периферии к центральному афферентному нейрону – это периферический отросток, центральный – от тела нейрона в мозговой центр.
- Моторные, двигательные состоят из эфферентных нейронов, и в зависимости от их положения их называют паравертебральные, предвертебральные.
Симпатические ганглии
Паравертебральные цепочки ганглиев расположены вдоль позвоночного столба в симпатических стволах, которые идут длинной вереницей от основания черепа до копчика.
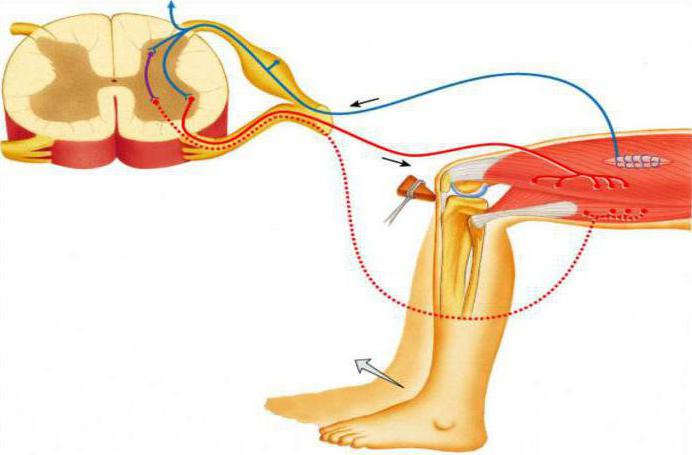
Предвертебральные нервные сплетения находятся ближе к внутренним органам, и их локализация сосредоточена перед аортой. Они образуют брюшное сплетение, которое состоит из солнечного, нижне- и верхнебрыжеечного сплетений. Они представлены двигательными адренергическими и тормозящими действие холинергическими нейронами. Также связь между нейронами осуществляется преганглионарными и постганглионарными нейронами, которые используют медиаторы ацетилхолин и норадреналин.
Интрамуральные нервные узлы имеют нейроны трех типов. Их описание было сделано русским ученым Догелем А.С., который, исследуя гистологию нейронов вегетативной нервной системы, выделил такие нейроны, как длинноаксонные эфферентные клетки первого типа, равноотросчатые афферентные клетки второго типа и ассоциативные клетки третьего типа.
Рецепторы ганглиев
Афферентные нейроны выполняют узкоспециализированую функцию, и их роль заключается в восприятии раздражителей. Такими рецепторами являются механорецепторы (реакция на растяжение или давление), фоторецепторы, терморецепторы, хеморецепторы (отвечают за реакции в организме, химические связи), ноцицепторы (реакция организма на болевые раздражители – это повреждение кожи и другие).
В симпатических стволах эти рецепторы по рефлекторной дуге передают в центральную нервную систему информацию, которая служит сигналом о повреждениях или нарушениях в организме, а также нормальной его работе.
Функции ганглиев
Каждый ганглий имеет свое местоположение, кровоснабжение, и его функции определяются этими параметрами. Спинномозговой ганглий, имеющий иннервацию из ядер головного мозга, обеспечивает непосредственную связь процессов в организме через рефлекторную дугу. От этих структурных компонентов спинного мозга иннервируются железы, гладкая мускулатура мышц внутренних органов. Сигналы, поступающие по рефлекторной дуге, идут медленнее, чем в ЦНС, и они полностью регулируются вегетативной системой, также она обладает трофической, сосудодвигательной функцией.
Базальные ганглии (стриопаллидарная система)– это совокупность трех парных образований, расположенных в конечном мозге на основании больших полушарий. Включает в себя 3 парных образования (рис.17):

Рис.17. Анатомия базальных ядер.
- бледный шар – состоит из наружного и внутреннего сегмента, является наиболее филогенетически древней частью;
- полосатое тело – состоит из хвостатого ядра и скорлупы;
- ограда – расположена между скорлупой и островковой корой, является наиболее молодой частью.
Установлены внутренние связи между базальными ганглиями. За их счет компоненты базальных ганглиев тесно взаимодействуют и образуют единую стриопаллидарную систему. Кроме того, базальные ганглии имеют многочисленные афферентные и эфферентные связи с другими структурами мозга.
Афферентные связи поступают (рис. 17) в базальные ядра от всех областей коры прямо и через таламус, от неспецифических ядер таламуса, от черного вещества.

Рис. 17. Схема основных афферентных и эфферентных связей базальных ядер:
ЧВ – черное вещество; ВЯ – вентральные ядра; ИЛЯ – интраламинарное ядро;
Имеются три эфферентных выхода:
1. от полосатого тела тормозящие пути идут к бледному шару, от бледного шара к двигательным ядрам таламуса и от них к двигательной коре (поля 4 и 6);
2. часть эфферентных волокон из бледного шара и полосатого тела идет к ретикулярной формации и красному ядру ствола мозга и далее в спинной мозг, а также через нижнюю оливу в мозжечок;
3. от полосатого тела тормозящие пути идут также к черному веществу и далее к ядрам таламуса.
В последнее время было выявлено, что связи базальных ганглий образуют параллельные функциональные петли (рис.18): скелетно-моторную и глазодвигательную петли.
Скелетно-моторная петля служит для регуляции таких параметров движения как амплитуда, сила и направление. Она соединяет премоторную, двигательную и соматосенсорную области коры со скорлупой базальных ядер, далее импульсация идет в бледный шар и черное вещество среднего мозга и через двигательное ядро (вентролатеральное ядро) таламуса возвращается в премоторную кору.
Глазодвигательная петля участвует в регуляции движений глаз (например, скачкообразных движений глаз – саккад). Она соединяет области коры, контролирующие направление взгляда (поле 8 лобной коры и поле 7 теменной коры), с хвостатым ядром. Оттуда импульсы поступают в бледный шар и черное вещество из которых она поступает в ассоциативное медиадорсальное и переднее релейное вентральное ядро таламуса и возвращается в лобное глазодвигательное поле 8.
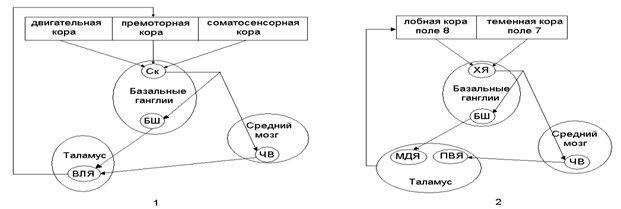
Рис.18. Схема функциональных петель, проходящих через
базальные ганглии (по Р.Шмидту, 1996, с изменениями)
1 – скелетно-моторная петля; 2 – глазодвигательная петля;
Ск – скорлупа; БШ – бледный шар; ЧВ – черное вещество;
ВЛЯ – вентролатеральное ядро; ХЯ - хвостатое ядро; МДЯ – медиодорсальное ядро; ПВЯ – переднее вентральное ядро
Функции базальных ядер
Основными функциями базальных ядер являются:
1. Регуляция двигательной активности. Важную роль базальные ядра играют при переходе от замысла движения (фазы подготовки к движению) к выбранной программе действия (фазе выполнения).
2. Участие в интеграции (объединении) тонических рефлексов и в обеспечении позы.
3. Участие, наряду с мозжечком, в выработке сложных двигательных программ.
4. Участие в контроле таких параметров движения как сила, амплитуда, скорость и направление движения.
Функции полосатого тела.Полосатое тело состоит из более крупных нейронов с длинными отростками, которые выходят за пределы стриапаллидарной системы. Полосатое тело участвует в регуляции мышечного тонуса, уменьшая его, а также в организации и регуляции движений. Стимуляция полосатого тела вызывает:
- простые двигательные реакции;
- торможение активности коры, подкорковых образований, безусловных и условных рефлексов;
- торможение поведенческих реакций (пищедобывательной и ориентировочной).
Полосатое тело оказывает на бледный шар тормозящее влияние (медиатор ГАМК), снижая активность бледного шара, и тем самым, снижая двигательную активность.
Полосатое тело имеет двухсторонние связи с черным веществом среднего мозга. Нейроны полосатого тела оказывают тормозное влияние (медиатор ГАМК) на нейроны черного вещества. Нейроны черного вещества секретируют медиатор дофамин, который посредством аксонного транспорта поступает к базальным ганглиям. При резком снижении концентрации дофамина в хвостатом ядре появляются симптомы болезни Паркинсона: сильное дрожание пальцев, губ, головы и других частей тела, ригидность мышц и т.д.
На двигательную кору полосатое тело также оказывает тормозное влияние, сдерживая ненужную в данный момент степень свободы движения, и тем самым обеспечивая четкость и направленность двигательных реакций.
Поражение полосатого тела сопровождается возникновением так называемых гиперкинезов. Гиперкинезы – это избыточные, насильственные, непроизвольные движения. Движения у больных становятся размашистыми, избыточным, широко амплитудными. Больные не могут усидеть на одном месте, все время вертятся, меняют положение тела и т.д. Кроме того, могут наблюдаться следующие симптомы:
· Тремор – подергивание конечностей;
· Ригидность мышц – повышение мышечного тонуса;
· Баллизм – крупноразмашистый гиперкинез конечностей;
· Атетоз – больше наблюдается в дистальных отделах конечностей – в кистях и пальцах рук; проявляется медленными, извивающимися, червеобразными движениями;
Скорлупа отвечает за организацию пищевого поведения. При раздражении скорлупы появляется слюноотделение и изменение дыхания, при поражении наблюдаются трофические изменения в коже.
Функции бледного шара. Бледный шар оказывает модулирующее влияние на двигательную кору, мозжечок, ретикулярную формацию и красное ядро. Его функцией является провоцирование ориентировочной реакции, движения конечностей, активация пищевого поведения (жевание, глотание).
Ограда – функция мало изучена. Имеет двухсторонние связи с лобной, затылочной, височной корой, обонятельной луковицей, таламусом и другими базальными ядрами. При раздражении ограды вызываются моторные реакции со стороны пищеварительного тракта (жевание, глотание, рвотные движения), ориентировочные реакции. При повреждении – теряется способность говорить.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.


Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.
ГАНГЛИИ (ganglia нервные узлы)— скопления нервных клеток, окруженные соединительной тканью и клетками глии, расположенные по ходу периферических нервов.
Различают Г. вегетативной и соматической нервной системы. Г. вегетативной нервной системы делятся на симпатические и парасимпатические и содержат тела постганглионарных нейронов. Г. соматической нервной системы представлены спинномозговыми узлами и Г. чувствительных и смешаных черепных нервов, содержащих тела чувствительных нейронов и дающих начало чувствительным порциям спинномозговых и черепных нервов.
Содержание
- 1 Эмбриология
- 2 Анатомия
- 3 Гистология
- 4 Физиология
- 5 Патология
- 6 Краткая анатомическая характеристика нервных ганглиев (узлов)
Эмбриология
Зачатком спинномозговых и вегетативных узлов является ганглиозная пластинка. Она образуется у эмбриона в тех отделах нервной трубки, которые граничат с эктодермой. У зародыша человека на 14—16-й день развития ганглиозная пластинка располагается по дорсальной поверхности замкнувшейся нервной трубки. Затем она расщепляется по всей длине, обе ее половины перемещаются вентрально и в виде нервных валиков лежат между нервной трубкой и поверхностной эктодермой. В дальнейшем соответственно сегментам дорсальной стороны зародыша в нервных валиках возникают очаги пролиферации клеточных элементов; эти участки утолщаются, обособляются и превращаются в спинномозговые узлы. Из ганглиозной пластинки развиваются также чувствительные ганглии У, VII—X пар черепных нервов, аналогичные спинномозговым ганглиям. Зародышевые нервные клетки, нейробласты, формирующие спинномозговые ганглии, представляют собой биполярные клетки, т. е. имеют два отростка, отходящих от противоположных полюсов клетки. Биполярная форма чувствительных нейронов у взрослых млекопитающих и человека сохраняется только в чувствительных клетках преддверно-улит кового нерва, преддверного и спирального ганглиев. В остальных, как спинномозговых, так и черепных чувствительных узлах, отростки биполярных нервных клеток в процессе их роста и развития сближаются и сливаются в большинстве случаев в один общий отросток (processus communis). По этому признаку чувствительные нейроциты (нейроны) называют псевдоуниполярными (neurocytus pseudounipolaris), реже протонейронами, подчеркивая древность их происхождения. Спинномозговые узлы и узлы в. н. с. отличаются характером развития и строения нейронов. Развитие и морфология вегетативных ганглиев — см. Вегетативная нервная система.
Анатомия
Основные сведения об анатомии Г. приведены в таблице.
Гистология


Возраст и перенесенные заболевания сказываются на структуре спинномозговых ганглиев — в них появляется значительно большее, чем у здоровых, количество различных атипичных нейронов, в особенности с дополнительными отростками, снабженными колбовидными утолщениями, как, напр., при ревматическом пороке сердца (рис. 2), стенокардии и др. Клинические наблюдения, а также экспериментальные исследования на животных показали, что чувствительные нейроны спинномозговых узлов реагируют значительно быстрее интенсивным ростом дополнительных отростков на различные эндогенные и экзогенные вредности, нежели моторные соматические или вегетативные нейроны. Эта способность чувствительных нейронов иногда выражена значительно. В случаях хрон, раздражений вновь образованные отростки могут обвиваться (в виде намотки) вокруг тела собственного или соседнего нейрона, напоминая кокон. Чувствительные нейроны спинномозговых узлов, как и другие типы нервных клеток, имеют ядро, различные органеллы и включения в цитоплазме (см. Нервная клетка). Т. о., отличительным свойством чувствительных нейронов спинномозговых и узлов черепных нервов является их яркая морфол, реактивность, выражающаяся в изменчивости их структурных компонентов. Это обеспечено высоким уровнем синтеза белков и различных активных веществ и свидетельствует об их функциональной подвижности.
Физиология
У беспозвоночных Г. играют ту же роль, что и ц. н. с. у позвоночных, являясь высшими центрами координации соматических и вегетативных функций. В эволюционном ряду от червей до головоногих моллюсков и членистоногих Г., перерабатывающие всю информацию о состоянии окружающей и внутренней среды, достигают высокой степени организации. Это обстоятельство, а также простота анатомической препаровки, относительно большие размеры тел нервных клеток, возможность введения в сому нейронов под прямым визуальным контролем одновременно нескольких микроэлектродов сделали Г. беспозвоночных общераспространенным объектом нейрофизиол, экспериментов. На нейронах круглых червей, октапод, декапод, брюхоногих и головоногих моллюсков методами электрофореза, прямого измерения активности ионов и фиксации напряжения проводят исследования механизмов генерации потенциалов и процесса синаптической передачи возбуждения и торможения, зачастую невыполнимые на большинстве нейронов млекопитающих. Несмотря на эволюционные различия, основные электрофизиол, константы и нейрофизиол, механизмы работы нейронов во многом одинаковы у беспозвоночных и высших позвоночных животных. Поэтому исследования Г., беспозвоночных имеют общефизиол. значение.
У позвоночных соматосенсорные черепные и спинномозговые Г. функционально однотипны. Они содержат тела и проксимальные части отростков афферентных нейронов, передающих импульсацию от периферических рецепторов в ц. н. с. В сомато-сенсорных Г. нет синаптических переключений, эфферентных нейронов и волокон. Так, нейроны спинномозговых Г. у жабы характеризуются следующими основными электрофизиол, параметрами: специфическим сопротивлением — 2,25 кОм/см 2 для деполяризующего и 4,03 кОм/см 2 для гиперполяризующего тока и удельной емкостью 1,07 мкФ/см 2 . Полное входное сопротивление нейронов сомато-сенсорных Г. существенно ниже, чем соответствующий параметр аксонов, поэтому при высокочастотной афферентной импульсации (до 100 импульсов в 1 сек.) проведение возбуждения может блокироваться на уровне тела клетки. В этом случае потенциалы действия, хотя и не регистрируются от тела клетки, продолжают проводиться с периферического нерва в задний корешок и сохраняются даже после экстирпации тел нервных клеток при условии интактности Т-образных ветвлений аксонов. Следовательно, возбуждение сомы нейронов сомато-сенсорных Г. для передачи импульсов от периферических рецепторов к спинному мозгу не обязательно. Эта особенность впервые появляется в эволюционном ряду у бесхвостых амфибий.
Вегетативные Г. позвоночных в функциональном плане принято разделять на симпатические и парасимпатические. Во всех вегетативных Г. происходит синаптическое переключение с преганглионарных волокон на постганглионарные нейроны. В подавляющем большинстве случаев синаптическая передача осуществляется хим. путем с помощью ацетилхолина (см. Медиаторы). В парасимпатическом цилиарном Г. птиц обнаружена электрическая передача импульсов при помощи так наз. потенциалов соединения, или потенциалов связи. Электрическая передача возбуждения через один и тот же синапс возможна в двух направлениях; в процессе онтогенеза она формируется позже химической. Функциональное значение электрической передачи пока не ясно. В симпатических Г. амфибий выявлено небольшое количество синапсов с хим. передачей нехолинергической природы. В ответ на сильное одиночное раздражение преганглионарных волокон симпатического Г. в постганглионарном нерве прежде всего возникает ранняя отрицательная волна (O-волна), обусловленная возбуждающими постсинаптическими потенциалами (ВПСП) при активации н-холинорецепторов постганглионарных нейронов. Тормозной постсинаптический потенциал (ТПСП), возникающий в постганглионарных нейронах под действием катехоламинов, выделяемых хромаффинными клетками в ответ на активацию их м-холинорецепторов, формирует следующую за 0-волной положительную волну (П-волна). Поздняя отрицательная волна (ПО-волна) отражает ВПСП постганглионарных нейронов при активации их м-холинорецепторов. Завершает процесс длительная поздняя отрицательная волна (ДПО-волна), возникающая вследствие суммации ВПСП нехолинергической природы в постганглионарных нейронах. В нормальных условиях на высоте О-волны при достижении ВПСП величины 8—25 мв возникает распространяющийся потенциал возбуждения с амплитудой 55—96 мв, длительностью 1,5—3,0 мсек, сопровождаемый волной следовой гиперполяризации. Последняя существенно маскирует волны П и ПО. На высоте следовой гиперполяризации возбудимость снижается (период рефрактерности), поэтому обычно частота разрядов постганглионарных нейронов не превышает 20—30 импульсов в 1 сек. По основным электрофизиол. характеристикам нейроны вегетативных Г. идентичны большинству нейронов ц. н. с. Нейрофизиол. особенностью нейронов вегетативных Г. является отсутствие истинной спонтанной активности при деафферентации. Среди пре- и постганглионарных нейронов преобладают нейроны групп В и С по классификации Гассера — Эрлангера, основанной на электрофизиол, характеристиках нервных волокон (см.). Преганглионарные волокна обширно ветвятся, поэтому раздражение одной преганглионарной ветви приводит к возникновению ВПСП во многих нейронах нескольких Г. (феномен мультипликации). В свою очередь на каждом постганглионарном нейроне оканчиваются терминали многих преганглионарных нейронов, отличающихся по порогу раздражения и скорости проведения (феномен конвергенции). Условно мерой конвергенции можно считать отношение количества постганглионарных нейронов к количеству преганглионарных нервных волокон. Во всех вегетативных Г. оно больше единицы (за исключением цилиарного ганглия птиц). В эволюционном ряду это отношение возрастает, достигая в симпатических Г. человека величины 100:1. Мультипликация и конвергенция, обеспечивающие пространственную суммации) нервных импульсов, в сочетании с временной суммацией являются основой интегрирующей функции Г. при обработке центробежной и периферической импульсации. Через все вегетативные Г. проходят афферентные пути, тела нейронов которых лежат в спинномозговых Г. Для нижнего брыжеечного Г., чревного сплетения и некоторых интрамуральных парасимпатических Г. доказано существование истинных периферических рефлексов. Афферентные волокна, проводящие возбуждение с малой скоростью (ок. 0,3 м/сек), входят в Г. в составе постганглионарных нервов и оканчиваются на постганглионарных нейронах. В вегетативных Г. обнаружены окончания афферентных волокон. Последние информируют ц. н. с. о происходящих в Г. функционально-хим. изменениях.
Патология
В клин, практике наиболее часто встречается ганглионит (см.), называемый также симпато-ганглионитом, — заболевание, связанное с поражением ганглиев симпатического ствола. Поражение нескольких узлов определяют как полиганглионит, или трунцит (см.).
Спинальные ганглии нередко вовлекаются в патологический процесс при радикулитах (см.).
Читайте также:
