
Где невозможно найти свободные нервные окончания в
Свободные нервные окончания — наиболее распространённый тип сенсорных рецепторов. Расположены в прослойках соединительной ткани внутренних органов, а также в соединительнотканной основе кожи, где они сосредоточены преимущественно в сосочках. Свободные нервные окончания эпидермиса расположены в базальном и шиповатом его слоях. В областях кожи с высокой тактильной чувствительностью (например, пальцы рук) терминали достигают зернистого слоя. Некоторые свободные окончания в соединительной ткани имеют билатеральную организацию и напоминают сэндвич: в центре расположена нервная терминаль эллиптической формы, а снаружи — вспомогательные клетки. Большинство свободных нервных окончаний — механорецепторы. Некоторые окончания в эпидермисе определённо специализированы для регистрации изменений температуры. Имеются также рецепторы, способные определять изменения рН, рО2 и рСО2.
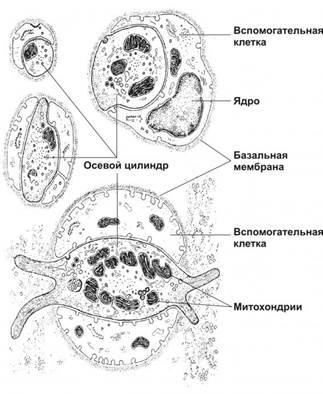
Рисунок 3. Свободные нервные окончания в соединительной ткани. Осевой цилиндр контактирует с вспомогательными клетками на значительном протяжении или полностью окружён ими. Свободное нервное окончание, изображённое в нижней части рисунка, имеет билатеральную организацию: в центре расположена эллиптической формы нервная терминаль, покрытая вспомогательными клетками.
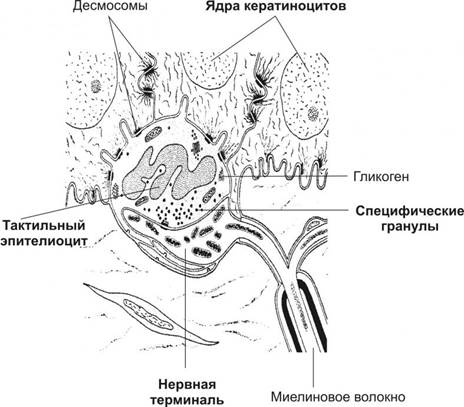
Рисунок 4. Комплекс тактильного эпителиоцита с нервной терминалью. Тактильный эпителиоцит расположен в базальном слое эпидермиса, образует отростки и связывается с кератиноцитами при помощи десмосом. Расширенная нервная терминаль вступает в контакт с тактильным эпителиоцитом. Специфические гранулы расположены в обращённой к нервной терминали части клетки.
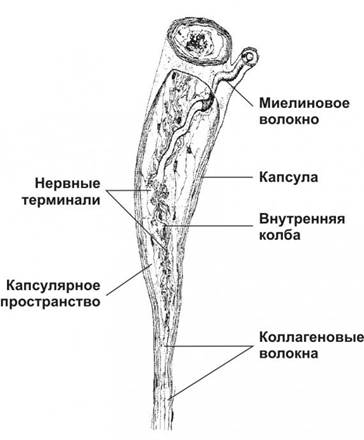
Рисунок 5. Веретеновидное тельце содержит внутреннюю колбу с густой сетью разветвлённых нервных терминалей и вспомогательными пластинчатыми клетками. Снаружи тельце покрыто соединительнотканной капсулой из нескольких слоёв уплощённых фибробластов. Между внутренней колбой и капсулой находится заполненное жидкостью капсулярное пространство.
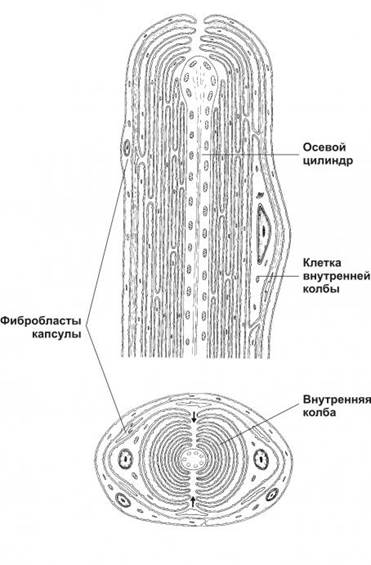
Рисунок 6. Пластинчатое тельце состоит из билатерально организованной внутренней колбы и наружной соединительнотканной капсулы. Клетки внутренней колбы образуют отростки, концентрическими полукольцами окружающие чувствительную нервную терминаль. Стрелками указана щель внутренней колбы, проходящая параллельно короткой оси эллипса нервной терминали.
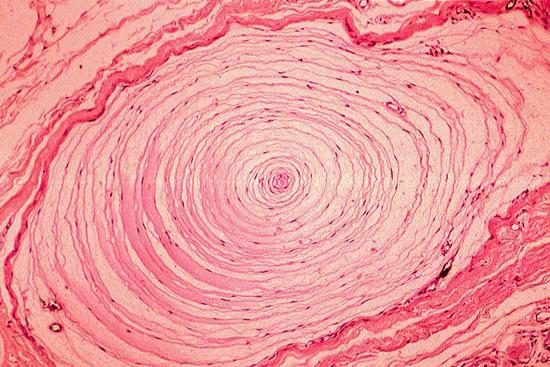
Рисунок 7. Пластинчатое тельце Пачини состоит из внутренней колбы и наружной соединительнотканной капсулы. Клетки внутренней колбы концентрическими полукольцами окружают чувствительную нервную терминаль. Капсулу образуют уплощённые фибробласты и коллагеновые волокна. Окраска гематоксилином и эозином.
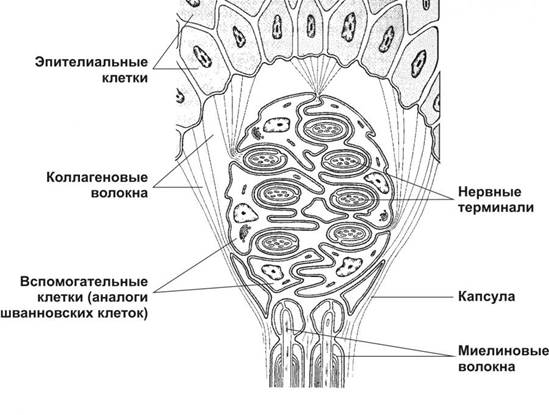
Рисунок 8. Тактильное тельце окружено соединительнотканной капсулой с вплетёнными в неё коллагеновыми волокнами окружающей соединительной ткани. Подходящие к тельцу нервные волокна теряют миелин и заходят внутрь тельца, где формируют многочисленные терминали, окружённые вспомогательными клетками.
В дальнейшем происходит образование более разветвленных окончаний. Некоторые основные нервные волокна, идущие к клеточному телу, позднее покрываются очень тонкой мякотной оболочкой. Остальные волокна остаются безмякотными.
Имеются также и другие типы свободных нервных окончаний, не связанные с восприятием болевых импульсов. Наиболее сложными из них являются сплетения волокон, оканчивающихся в волосяных сумках. Они обладают очень высокой чувствительностью, так как реагируют на самое незначительное давление, достаточное для того, чтобы отклонить волос, с которым они связаны.
Инкапсулированные окончания представляют собой нервные окончания, заключенные в соединительную ткань. Они различаются характером ветвления нервного волокна, количеством и строением покрывающей соединительной ткани, размерами и формой органа как целого. В этой группе есть большие пластинчатые тельца Пачини, связанные с чувством глубокого давления; сходные по строению, но более мелкие тельца Гольджи-Маццони, тактильные тельца Мейснера и половые тельца.
Тактильные тельца формируются в процессе развития относительно поздно, как и большинство инкапсулированных окончаний. Соответственно окрашенные препараты плодов последних трех месяцев развития обнаруживают характерные концевые петли нервных волокон в кожных сосочках. Их соединительнотканная капсула сначала совсем тонкая и слабо заметна на препаратах. Однако развивающаяся соединительная ткань капсулы хорошо заметна на препаратах более поздних плодов, окрашенных гематоксилином и эозином, что указывает на ее дефинитивное строение.
Тельца Пачини образуются примерно так же, как и тактильные тельца. Концевые нервные волокна телец Пачини теряют некоторые веточки и образуют основные пучки, расположенные по оси, с короткими, отходящими в стороны ветвями. Вторичная соединительнотканная капсула становится значительно толще и в конце концов напоминает луковицу с концентрически расположенными слоями.
Очень близкими к чувствительным нервным окончаниям предшествующей группы являются нервно-мышечные и нервно-сухожильные окончания, при помощи которых мы ощущаем положение тела и его частей. Нервно-мышечные веретена представляют собой мышечные волокна, которые обвиты спиральными и кольцеобразными нервными окончаниями. Эти окончания вторично инкапсулированы в соединительную ткань. Они реагируют на растяжение мышечных волокон.
Нервно-сухожильные окончания являются веретенообразными образованиями, концевые нервные разветвления которых расположены между сухожильными волокнами. Сухожильные волокна и связанные с ними нервные окончания покрыты общей соединительнотканной оболочкой. Сильно разветвленные нервные волокна, из которых образуются нервно-мышечные и нервно-сухожильные окончания, могут быть замечены в эмбрионах 4,5 месяцев развития, но их окончательное положение по отношению к отдельным мышечным или сухожильным волокнам устанавливается несколько позднее.
Автор работы: Пользователь скрыл имя, 08 Февраля 2013 в 01:59, реферат
Эти нервые окончания - рецепторы - рассеяны по всему организму и воспринимают различные раздражения как из внешней среды, так и от внутренних органов. В зависимости от специфичности раздражения, воспринимаемого данным видом рецептора, все чувствительные окончания разделяют на свободные нервные окончания, т. е. состоящие только из конечных ветвлений осевого цилиндра. Характерны для эпителия (холодовые рецепторы).
свободные нервные окончания.doc
Рецепторные (чувствительные) нервные окончания
Эти нервые окончания - рецепторы - рассеяны по всему организму и воспринимают различные раздражения как из внешней среды, так и от внутренних органов.
В зависимости от специфичности раздражения, воспринимаемого данным видом рецептора, все чувствительные окончания разделяют на
свободные нервные окончания, т. е. состоящие только из конечных ветвлений осевого цилиндра. Характерны для эпителия (холодовые рецепторы).
В этом случае миелиновые нервные волокна подходят к эпителиальному пласту, теряют миелин, а осевые цилиндры проникают в эпителий и распадаются там между клетками на тонкие терминальные ветви. В многослойном эпителии есть окончания, в состав которых входят, кроме терминалей отростков нервных клеток, специфически измененные эпителиальные клетки - осязательные эпителиоциты. От других клеток эпителия они отличаются светлой цитоплазмой, наличием осмиофильных гранул диаметром 65-180 нм и уплощенным темным ядром. Концевые нервные веточки подходят к таким клеткам и расширяются, образуя дисковидные концевые структуры, связанные с основаниями осязательных эпителиоцитов.
Свободные нервные окончания (рис. 12 и рис. 12а) — наиболее распространённый тип сенсорных рецепторов.
Локализация. Свободные окончания расположены в прослойках соединительной ткани внутренних органов, а также в соединительнотканной основе кожи, где они сосредоточены преимущественно в сосочках. Свободные нервные окончания эпидермиса расположены в базальном и шиповатом его слоях. В областях кожи с высокой тактильной чувствительностью (например, пальцы рук) терминали достигают зернистого слоя.
Строение. Для рассматриваемого вида чувствительных нервных окончаний, локализованных в соединительной ткани, термин "свободное" условен, т.к. ветвления осевого цилиндра, как правило, окружены вспомогательными клетками (аналогичны шванновским). Варианты взаимодействия вспомогательных клеток с осевым цилиндром могут быть различны. Так, вспомогательная клетка может покрывать только часть поверхности осевого цилиндра. Остальная его часть отделена от окружающей соединительной ткани лишь базальной мембраной, обычно расположенной по всей поверхности свободного нервного окончания. Нервная терминаль может формировать пальцевидные выросты, проникающие в окружающую соединительную ткань между вспомогательными клетками.
Билатеральная организация. Некоторые свободные окончания в соединительной ткани имеют билатеральную организацию и напоминают сэндвич: в центре расположена нервная терминаль эллиптической формы, а снаружи — вспомогательные клетки. Полагают, что такая организация необходима для регистрации направления механического смещения.
Модальность. Большинство свободных нервных окончаний — механорецепторы. Некоторые окончания в эпидермисе определённо специализированы для регистрации изменений температуры. По настоящее время не идентифицированы т.н. холодовые и тепловые окончания. Имеются также рецепторы, способные определять изменения рН, рО2 и рСО2.
Среди нервных окончаний различают свободные, лишенные глиальных клеток, и несвободные, у которых нервные окончания имеют оболочку - капсулу, образованную клетками нейроглии или соединительнотканными элементами.
Свободные нервные окончания имеются в коже. Подходя к эпидермису, нервное волокно теряет миелин, проникает через базальную мембрану в эпителиальный слой, где разветвляется между эпителиоцитами вплоть до зернистого слоя. Конечные ветви диаметром менее 0,2 мкм на своих концах колбообразно расширяются. Аналогичные нервные окончания имеются в эпителии слизистых оболочек и в роговице глаза. Концевые свободные рецепторные нервные окончания воспринимают боль, тепло и холод. Другие нервные волокна проникают таким же образом в эпидермис и заканчиваются контактами с осязательными клетками (клетки Меркеля). Нервное окончание расширяется и образует с клеткой Меркеля синапсоподобный контакт. Эти окончания являются механорецепторами, воспринимающими давление.
III. По строению
рецепторов: 1. свободные нервные окончания (конечные ветвления осевого цилиндра лишены оболочки);
1. В эпителии кожи находятся свободные рецепторные окончания.
2. а) Одни из них просто проникают между клетками эпителия.
б) Другие контактируют с основаниями осязательных эпителиоцитов (специфически изменённых эпителиальных клеток).
3. Эти рецепторы способны воспринимать даже очень слабые раздражения, реагируя на давление (прикосновение) и температуру.
Все рецепторы делятся на свободные нервные окончания и инкапсулированные. Свободные нервные окончания вступают в непосредственный контакт с иннервируемой тканью. К ним относятся: диск Меркеля и тельце Гранди. К инкапсулированным нервным окончаниям относятся такие, которые вокруг концевых разветвлений нервного волокна имеют специализированные элементы, образующие различной сложности пластичную капсулу. В зависимости от устройства глиальной капсулы, различают: тельца Месснера, диски Меркеля, тельца Пачини, окончания Руффини.
По особенностям строения чувствительные окончания подразделяют на свободные нервные окончания, т.е. состоящие только из конечных ветвлений осевого цилиндра,
Свободные нервные окончания обычно воспринимают холод, тепло и боль. Такие окончания характерны для эпителия. В этом случае миелиновые нервные волокна подходят к эпителиальному пласту, теряют миелин, а осевые цилиндры проникают в эпителий и распадаются там между клетками на тонкие терминальные ветви. Инкапсулированные рецепторы соединительной ткани при всем их разнообразии всегда состоят из ветвления осевого цилиндра и глиальных клеток. Снаружи такие рецепторы покрыты соединительнотканной капсулой. Примером подобных окончаний могут служить весьма распространенные у человека пластинчатые тельца (или тельца Фатера — Пачини). В центре такого тельца располагается внутренняя луковица, или колба, образованная видоизмененными леммоцитами. Миелиновое чувствительное нервное волокно теряет около пластинчатого тельца миелиновый слой, проникает во внутреннюю луковицу и разветвляется. Снаружи тельце окружено слоистой капсулой, состоящей из фибробластов и спирально ориентированных волокон. Заполненные жидкостью пространства между пластинками содержат коллагеновые микрофибриллы. Давление на капсулу передается через заполненные жидкостью пространства между пластинками на внутреннюю луковицу и воспринимается безмиелиновыми волокнами во внутренней луковице. Пластинчатые тельца воспринимают давление и вибрацию. Они присутствуют в глубоких слоях дермы (особенно в коже пальцев), в брыжейке и внутренних органах. К чувствительным инкапсулированным окончаниям относятся осязательные тельца — тельца Мейснера. Эти структуры имеют овоидную форму, располагаются в верхушках соединительнотканных сосочков кожи. Осязательные тельца состоят из видоизмененных нейролеммоцитов — тактильных клеток, расположенных перпендикулярно длинной оси тельца. Части тактильных клеток, содержащие ядра, расположены на периферии, а уплощенные части, обращенные к центру, формируют пластинчатые отростки, интердигитирующие с отростками противоположной стороны. Тельце окружено тонкой капсулой. Миелиновое нервное волокно входит в основание тельца снизу, теряет миелиновый слой и формирует ветви, извивающиеся между тактильными клетками. Коллагеновые микрофибриллы и волокна связывают тактильные клетки с капсулой, а капсулу с базальным слоем эпидермиса, так что любое смещение эпидермиса передается на осязательное тельце Мейснера. К инкапсулированным нервным окончаниям относятся также рецепторы мышц и сухожилий: это нервно-мышечные веретена и нервно-сухожильные веретена. Нервно-мышечные веретена являются сенсорными органами в скелетных мышцах, которые функционируют как рецептор на растяжение. Веретено состоит из нескольких исчерченных мышечных волокон — интрафузальных волокон, заключенных в растяжимую соединительнотканную капсулу. Между капсулой и интрафузальными волокнами имеется заполненное жидкостью пространство. Интрафузальные волокна имеют актиновые и миозиновые миофиламенты только на концах, которые и сокращаются. Рецепторной частью интрафузального мышечного волокна является центральная, несокращающаяся часть. К интрафузальным мышечным волокнам подходят афферентные нервные волокна. При расслаблении (или растяжении) мышцы увеличивается и длина интрафузальных волокон, что регистрируется рецепторами. Одни окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, другие — реагируют только на изменение длины. При внезапном растяжении в спинной мозг поступает сильный сигнал, вызывающий резкое сокращение мышцы, с которой поступил сигнал, — динамический рефлекс на растяжение. При медленном, длительном растяжен
Рецепторные (чувствительные) нервные окончания воспринимают сигналы из внешней среды (экстерорецепторы) и внутренних органов (интерорецепторы). В зависимости от природы раздражения, регистрируемого рецепторами, они подразделяются в соответствии с физиологической классификацией, на механорецепторы, хеморецепторы, терморецепторы и болевые рецепторы (ноцицепторы). В специализированных органах чувств (орган вкуса, обоняния, зрения, равновесия и слуха) имеются особые рецепторные клетки, которые воспринимают соответствующие раздражения.
Морфологическая классификация чувствительных нервных окончанийоснована на особенностях их структурной организации. В соответствии с этой классификацией различают свободные и несвободные чувствительные нервные окончания; последние включают инкапсулированные и неинкапсулированные окончания (рис. 8.18).

Рис. 8.18. Рецепторные (чувствительные) нервные окончания (по Rohen J.W., Lutjen-Drecoll E. 1982, с изменениями). 1 - свободные нервные окончания (СНО) образованы терминальными ветвлениями дендрита чувствительного нейрона, идущего в составе нервного волокна (НВ). Несвободные инкапсулированные нервные окончания (2-4) образованы ветвлениями дендрита, окруженными леммоцитами, в совокупности с которыми они образуют структуру, называемую внутренней колбой (ВК). Снаружи окончания покрыты соединительнотканной капсулой (СТК). 2 - колба Краузе, 3 - осязательное тельце (Мейснера), 4 - пластинчатое тельце (Фатер-Пачини).
Свободные чувствительные нервные окончаниясостоят только из терминальных ветвлений дендрита чувствительного нейрона, Они встречаются в эпителии, а также в соединительной ткани. Проникая в эпителиальный пласт, нервные волокна утрачивают миелиновую оболочку и нейролемму, а базальная мембрана их леммоцитов сливается с эпителиальной. Свободные нервные окончания обеспечивают восприятие температурных (тепловых и холодовых), механических и болевых сигналов (рис. 8.19).

Рис. 8.19. Свободные нервные окончания в соединительной ткани, представленные концевыми ветвлениями дендритов чувствительных нейронов.
Несвободные чувствительные нервные окончаний содержат все компоненты нервного волокна. Они разделяются на инкапсулированные (имеющие особую соединительнотканную капсулу) и неинкапсулированные.
Несвободные неинкапсулированные нервные окончания состоят из ветвлений дендритов, окруженных леммоцитами. Они встречаются в соединительной ткани кожи (дерме), а также собственной пластинки слизистых оболочек.
Несвободные инкапсулированные нервные окончания весьма разнообразны, но имеют единый общий план строения: их основу составляют ветвления дендрита, которые непосредственно окружены леммоцитами и снаружи покрыты особой соединительнотканной капсулой (см. рис. 8.18.). К этому виду нервных окончаний относят пластинчатые тельца (Фатер-Пачини), осязательные тельца (Мейснера) тельца Руффини, колбы Краузе, нервно-мышечные веретена и нервно-сухожильные веретена (сухожильные органы Гольджи).
Пластинчатые тельца (Фатер-Пачини) встречаются в соединительной ткани внутренних органов и кожи (рис. 8.20.). Они имеют вид округлых образований диаметром 1-5мм, воспринимают давление и вибрацию. Структурными компонентами тельца являются:
1) внутренняя колба (луковица), образованная видоизмененными уплощенными леммоцитами, в которую проникают одно или несколько нервных волокон, имеющих прямой ход;
2) наружная колба - слоистая соединительнотканная капсула, состоящая из фибробластов и коллагеновых волокон, образующих 10-60 концентрических пластин, между которыми имеется жидкость.
При деформации пластин капсулы давление передается на нервное окончание, что вызывает деполяризацию его мембраны.
 А |  Б |
Рис. 8.20. На электронной микрофотографии изображены: Тельца Фатер-Пачини (РС) – инкапсулированный рецептор. Капсула состоит из параллельно расположенной пластинки (по-видимому, производной Шванновской клетки), коллагеновых волокон. В центре капсулы раположены ветвления одного немиелинизированного нервного волокна (х100).
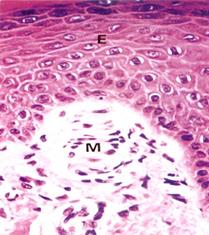
Осязательные тельца (Мейснера) расположены преимущественно в сосочковом слое дермы, имеют эллипсоидную форму и небольшие размеры (около 50-120 мкм) (рис. 8.21.). Их внутренняя колба состоит из плоских глиальных клеток, лежащих перпендикулярно длинной оси тельца, между которыми располагаются веточки дендритов. Между глиальными клетками проникают коллагеновые фибриллы, связанные с базальным слоем эпителия. Капсула тонкая, переходит в периневрий.
 А |  Б |
Рис. 8.21. Тельца Мейснера. Инкапсулированный рецептор (М), находящийся в коже губ, гениталий и т.д. Имеет овальную форму, располагается в коже прямо под эпидермисом (Е). Рецептор состоит из капсулы, образованной коллагеновыми волокнами, Шванновскими клетками (х320). (Б метод импрегнации х150). В капсуле находятся несколько безмякотных нервных окончаний, которые обильно ветвятся.).
Тельца Руффини лежат в соединительнотканной части кожи и капсулах суставов; они воспринимают давление и имеют вид веретеновидных структур длиной до 1-2 мм. Внутреннюю колбу образуют глиальные клетки, между которыми располагаются многочисленные ветвящиеся терминали дендритов с расширениями на концах. Капсула хорошо выражена, образована коллагеновыми волокнами.
Колбы Краузе - мелкие (40-150 мкм) округлые тельца, являющиеся механорецепторами и, возможно, холодовыми рецепторами. Они расположены в соединительной ткани сосочкового слоя дермы и собственной пластинке слизистой оболочки полости рта, надгортанника, в конъюнктиве глаза. Внутренняя колба образована уплощенными глиальными клетками, между которыми тонкие веточки дендрита образуют сплетение в виде клубочка. Капсула состоит из плоских клеток, являющихся продолжением периневрия.
Нервно-мышечные веретена - рецепторы растяжения волокон поперечнополосатых мышц - сложные инкапсулированные нервные окончания, обладающие как чувствительной, так и двигательной иннервацией. Число веретен в мышце зависит от ее функции и тем выше, чем более точными движениями она обладает. Нервно-мышечное веретено (рис. 8.22, 8.23.) имеет длину 0,5-7 мм и располагается параллельно ходу волокон мышцы, называемых экстрафузальными (от лат. extra - вне и fuso - веретено, т.е. расположенными за пределами веретена). Веретено покрыто тонкой соединительнотканной капсулой (продолжением периневрия), внутри которой находятся тонкие поперечнополосатые интрафузальные мышечные волокна двух видов:
- волокна с ядерной сумкой - в расширенной центральной части которых содержатся скопления ядер (1-4 волокна/веретено);
- волокна с ядерной цепочкой - более тонкие с расположением ядер в виде цепочки в центральной части (до 10 волокон/веретено).
Чувствительные нервные волокна образуют кольцеспиральные окончания на центральной части ишрафузальных волокон обоих типов и гроздьевидные окончания у краев волокон с ядерной цепочкой.
Двигательные нервные волокна - тонкие, образуют мелкие нервно-мышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус.
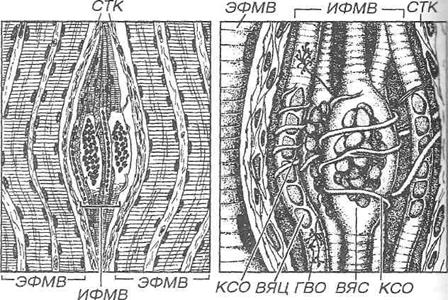
Рис. 8.22. Нервно-мышечное веретено. 1 - общий вид веретена, располагающегося между экстрафузальными мышечными волокнами (ЭФМЗ) и образованного интрафузальными мышечными волокнами (ИФМВ), которые окружены соединительнотканной капсулой (СТК). 2 - детали строения веретена, содержащего два вида ИФМВ: волокна с ядерной сумкой (ВЯС) и волокна с ядерной цепочкой (ВЯЦ). Чувствительные нервные волокна образуют кольцеспиральные окончания (КСО) на центральной части ИФМВ обоих типов и гроздьевидные окончания (ГВО) у краев ВЯЦ. Нервно-мышечное веретено содержит также двигательные нервные волокна и образованные ими нервно-мышечные синапсы по краям ИФМВ (не показаны).
 А |  Б |
Рис. 8.23. Нервно-мышечное веретено N-нервное волокно, С – капсула.А – продольный срез (х320), Б – поперечный.
Нервно-сухожильные веретена (сухожильные органы Гольджи) - рецепторы растяжения - веретеновидные инкапсулированные структуры длиной около 0.5-1мм, располагающиеся в области соединения волокон поперечнополосатых мышц с коллагеновыми волокнами сухожилий. Каждое веретено образовано капсулой из плоских фиброцитов (продолжение периневрия), которая охватывает группу сухожильных пучков, оплетенных многочисленными терминальными веточками нервных волокон, частично покрытых леммоцитами. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.
СПИСОК ЛИТЕРАТУРЫ
ОСНОВНАЯ ЛИТЕРАТУРА
1. Заварзин А.А. Основы сравнительной гистологии. Л. ЛГУ, 1985.- 397 с.
2. Гистология. Под ред. Ю.И. Афанасьева, Н.А.Юриной.- М.: Медицина, 1989.- 670 с.
3. Хэм А., Кормак Д. Гистология. (в 5 томах). М.: Мир. 1982.
4. Антипчук Ю.П. Гистология с основами эмбриологии. - М:Просвещение. 1983.- 240 с.
5. Антипчук Ю.П. Гiстологiя з основами ембрiологii. – K.: Вища школа. 1976.- 141 с.
6. Волкова О.В., Елецкий О.Н. Основы гистологии с гистологической техникой. –М.: Медицина, 1982.- 302 с.
7. Мануилова Н.А. Гистология с основами эмбриологии. М.: Просвещение, 1973.- 214 с.
8. Троценко Б.В., Чирский Н.В. Учебное пособие по курсу гистологии (электронная версия). – Симферополь, 2001.
9. Гистология (введение в гистологию) / под ред. Э.Г. Улумбекова, Ю.А. Челещева. – М.: ГЭОТАР, 1997.
10. Гистология / Ю.И. Афанасьев, Н.А. Юрина, Б.В. Алешин. – М.: Медицина, 1989.
11. Гистология/ Ю.И. Афанасьев, Н.А. Юрина, Е.Ф. Котовский и др. – М.: Медицина, 2002. –
12. Луцик О.Д., Иванова А.И. Кабак К.С. Гистология людини. – Лвів. Мир, 1992.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
1. Елисеев В.Г. Гистология. М.: Медицина, 1983.
2. Куприянов В.В. и др. Сосудистый эндотелий.- К.: Здоровье. – 248 с.
3. Абрамов М.Г. Гематологический атлас. –М.: Медицина. 1985.- 237 с.
4. Шаде Дж., Форд Д. Основы неврологии. М.: Мир, 1976.- 350 с.
5. Гистология (Введение в патологию). Под ред. Э.Г. Улумбекова, Ю.А. Челышева.-М.: ГЭОГАР. 1997.- 947 с.
6. Прохончуков А.А. и др. Гомеостаз костной ткани в норме и при экстремальном воздействии.- М.: Наука, 1984.- 200 с.
7. Соколов В.Е. и др. Адаптивные свойства эпителия и его производные. Атлас микрофотографий.- М.: Наука, 1979.- 110 с.
8. Гурфинкель В.С., Левик Ю.С. Скелетная мышца и функция. –М.: Наука,1985.- 143 с.
9. Кауфман О.Я. Гипертрофия и регенерация гладких мышц. М.:Наука. 1979.- 183 с.
10. Клишов А.В. Гистогенез и регенерация тканей.- Л.: Медицина, 1984.–30 с.
11. Комиссарчик Я.Ю., Миронов А.А. Электронная микроскопия клеток и тканей.- Л.: Наука, 1990.- 140 с.
12. Заварзин А.А. Основы частной цитологии и сравнительной гистологии многоклеточных животных. Л.:Наука, 1976.
13. Гацко Г.Г. Жировая ткань при старении. Минск. Наука и техника, 1985.- 184 с.
14. Гаврилов О.К. Клетки костного мозга и периферической крови –М.: Медицина, 1985.- 288 с.
15. Вельш У., Шторх Ф. Введение в цитологию и гистологию животных. М.: Мир.1976.
16. Виноградов В.В., Воробьёва Н.Ф. Тучные клетки, Новосибирск, Наука, 1973.
17. Занябуш Л. Молекулярная и клеточная биология. М.: Мир. 1982. Т.3.
18. Зенгебуш П. Молекулярная биология. М.: Мир, т.1-3. 1982.
19. Саркисов Д.С. Очерки по структурным основам гемостаза. М.: Медицина, 1977.
21. Серов В.В., Шехтер А.Б. Соединительная ткань. М.: Медицина. 1981.
22. Касавина Б.А., Торбенко В.П. Жизнь костной ткани. М.: Наука. 1979.
23. Кассиль Г.Н. Внутренняя среда организма. М.: Наука. 1983.- 277 с.
24. Пигаревский З.В. Зернистые лейкоциты и их свойства. М.: Мир, 1978.
25. Питерс А., Палей С., Уэбстер С. Ультраструктура нервной системы. М.: Мир. 1972.
26. Шубникова Е.А. Лекции по гистологии. М.: МГУ, 1973.
27. Мозг (пер. с анг. Под ред. Симонова П.В.) М.: Мир. 1982.
28. Фриденштейн А.Я., Чертков И.М. Клеточные основы иммунитета. М.: Медицина. 1981.
Свободные нервные окончания
Свободные нервные окончания – это наиболее многочисленные рецепторы, которые передают информацию в спинной мозг через мельчайшие миелинизированные и немиелинизированные нервные волокна. Свободные нервные окончания иннервируют практически все структуры и ткани тела: брюшные мышцы, различные виды соединительной ткани, крупные кровеносные сосуды и даже мышечные и нервно-сухожильные веретена.
Свободные нервные окончания чувствительны к механическим стимулам, которые связаны с мышечным сокращением, давлением или растяжением (отчего и возникают боли в спине или суставах). Другие свободные нервные окончания реагируют главным образом на изменение температуры или химической среды, например на увеличение концентрации ионов калия К + или молочной кислоты. Некоторые свободные нервные окончания имеют высокий порог и посылают импульсы только в ответ на стимулы, способные вызвать повреждение ткани. Это ноцирецепторные окончания, которые информируют об участке повреждения.
Некоторые немиелинизированные окончания становятся более активными в процессе ишемических сокращений. Они могут отвечать не только за мышечный дискомфорт (увеличение концентрации ионов калия К + или молочной кислоты во время выполнения физической нагрузки), но и за рефлексы сердечно-сосудистой и дыхательной систем (ИБС, ХОБЛ).
Таким образом, если рассматривать отдельно нервы (неврология) и мышцы (анатомия), то такой способ постановки диагноза при болях будет некорректным – в данном случае при болях в спине. Одним словом, болят только мышцы, в которых находятся органы чувств – рецепторы, передающие информацию в ЦНС через аксоны мотонейронов. Разрывая эту связь с помощью обезболивающих наркотических и ненаркотических средств, а тем более оперативно, организм теряет способность к саморегулированию и становится беззащитным перед лицом любой, даже обычной бытовой нагрузки.
Из всего вышесказанного можно сделать следующий очень важный вывод: для назначения лечения при болях в спине необходимо исследовать функцию мышц. Специалисты по современной кинезитерапии используют для этой цели миофасциальную диагностику и миофасциограф (метод Бубновского).
Таким образом, если понимать, что боли исходят из мышц, а интенсивность боли зависит от количества рецепторов, находящихся в мышцах, то отрегулировать интенсивность болевого синдрома можно естественным путем – то есть без обезболивающих лекарств и тем более без хирургического вмешательства. Для этого необходимо устранить спазмы в мышцах, издающих крик о помощи, с помощью упражнений, которые, в свою очередь, устраняют дистрофию. То есть при активизации мышцы снова начинают активно доставлять питание (кровь и воду) в позвоночник и попутно выводить из него уже отработанные клеточные элементы. Это называется метаболизм (обмен веществ).
Боли в мышцах (например, мышцах спины) – это причина болезни. А грыжа или остеохондроз – это следствие плохой работы этих мышц. К тому же если понять, как работает этот анатомо-физиологический и нервный механизм, можно понять и то, что и остеохондроз, и грыжа МПД – это старение позвоночника, а не болезнь. И акцент на грыжи, которые якобы ущемляют нервы, – это миф, созданный невропатологами. Но последним мне хотелось бы напомнить, что существует сосудистый пучок (нерв – вена – артерия), и изолированно повредить нерв можно лишь хирургически или травматически (ножевой или пулевой раной). В таком случае возникает паралич нерва и органов движения, за которые отвечает этот нерв. Но при болях в спине при остеохондрозе паралича не возникает! Поэтому для снятия или уменьшения болевого синдрома необходимо обеспечить возможность активации соответствующей группы мышц.
Причина болезни кроется в болях в мышцах, а грыжа или остеохондроз – это следствие плохой работы этих мышц.
Читайте также:
