
Головные нервы позвоночных животных
Периферический отдел нервной системы. Общие закономерности строения
Закономерности хода и ветвления нервов
Развитие периферического отдела нервной системы
Черепно-мозговые нервы. Общая характеристика
Развитие черепно-мозговых нервов
Список используемой литературы
Периферический отдел нервной системы. Общие закономерности строения
Периферический отдел нервной системы или периферические проводящие нервные пути состоят из спинномозговых нервов, отходящих от спинного мозга, и черепно-мозговых нервов, отходящих от головного мозга в количестве 12 пар.
Нерв- nervus –образован пучками нервных волокон, то есть отростками рецепторных (афферентных) и эфферентных нейронов, и поэтому проводит импульсы в разных направлениях:
а) с периферии к мозговым центрам;
б) из мозговых центров на периферию к исполнительным органам.
Таким образом, все нервы являются смешанными и содержат чувствительные и двигательные нервные волокна. Кроме нервных волокон в нерве имеется соединительно-тканный остов с его сосудами и нервами. В каждом нерве проходят также и симпатические волокна.
Нервы имеют вид белых тяжей разнообразной формы, более округлой или же более плоской. Толщина нервов также колеблется от микроскопических размеров до нескольких сантиметров. Крупные нервы хорошо заметны макроскопически.
Отдельное нервное волокно состоит из осевого цилиндра и неврилеммы, или шванновской оболочки. Осевой цилиндр образован или нейритом или дендритом нейрона. Неврилемма построена из шванновских или немиловских клеток – олигодендроцитов. Она может содержать в своем остове миелин и называется тогда миелиновой или мякотной оболочкой. В соответствии со строением шванновской оболочки и нервы разделяются на мякотные – с миелиновой оболочкой и безмякотные- амиелиновые.
Значение мякотной оболочки заключается в том, что она способствует лучшему проведению нервного возбуждения; в безмякотных нервных волокнах возбуждение проводится со скоростью 1-2 м/с, в то время как в мякотных волокнах скорость достигает 60-120 м/с.
Диаметр отдельных нервных волокон неодинаков: самые толстые из мякотных волокон- это двигательные соматические; средние мякотные волокна- чувствительные соматические; тонкие- чувствительные симпатические; тонкие безмякотные- симпатические двигательные.
В соединительнотканном остове нерва различают эндоневрий, периневрий и эпиневрий. Эндоневрий- endoneurium- упаковывает отдельные нервные волокна, периневрий –perineurium- пучки нервных волокон, а эпиневрий- epineurium- служит наружной оболочкой нерва как целого. Под эндо - и периневрием имеются периневральные лимфатические пространства, сообщающиеся в сторону мозга с субдуральными и субарахноидальными пространствами, а на периферии- с лимфатическими капиллярами.
В остове нерва проходят сосуды нерва – vasa nervorum- обеспечивающие его питание, и нервы сосудов нерва- nervi nervorum. В эпиневрии проходят и лимфатические сосуды, которые, однако, не сообщаются с периневральными пространствами. Число нервных волокон в нерве огромно; в периферическом конце его число волокон, учитывая отходящие коллатерали, может возрасти на 16%.
Закономерности хода и ветвления нервов
1.Спинномозговые нервы отходят метамерно от спинного мозга в каждом сегменте и разделяются на шейные, грудные, поясничные, крестцовые и хвостовые. Черепно-мозговые нервы отходят из продолговатого и среднего мозга. 1 и 2 черепно-мозговые пары нервов занимают в этом отношении особое положение.
2.Каждый спинномозговой нерв имеет два корня- дорсальный и вентральный- radix dorsalis et ventralis.на дорсальном корне находится межпозвоночный спинномозговой узел- ganglion spinale. оба корня у выхода из позвоночного канала соединяются в общий ствол- спинномозговой нерв- n.spinalis- содержащий чувствительные, двигательные, симпатические волокна. Черепно-мозговые нервы отходят только одним корнем, соответствующим дорсальному или вентральному корню спинномозгового нерва.
3.Все эфферентные нервные волокна выходят из вентральных столбов серого мозгового вещества спинного мозга и из соответствующих двигательных ядер продолговатого и среднего мозга. На спинном мозге они формируют вентральные двигательные корни.
4.Все афферентные нервные волокна состоят из нейритов клеток спинномозговых узлов и соответственно ганглиев черепно-мозговых нервов. Следовательно, все тела рецепторных нейронов лежат вне спинного и головного мозга.
5.Каждый грудопоясничный спинномозговой нерв по выходе из позвоночного канала отдает белую соединительную ветвь- ramus communicans albus – в симпатический ствол, ветвь в оболочки спинного мозга-ramus meningeus-затем получает серую соединительную ветвь- ramus communicans grisea- от симпатического ствола и делится на дорсальную и вентральную ветви-ramus dorsalis et ventralis- соответственно разграничению туловищной мускулатуры на дорсальный и вентральный мышечные тяжи с их сосудами. Каждая из упомянутых ветвей, в свою очередь, делится на медиальную и латеральную ветви- ramus medialis et lateralis- для мускулатуры и кожи, что также обуславливается разделением мышечных тяжей на латеральный и медиальный пласты. Совокупность ветвей каждого сегментного нерва вместе с соответствующим участком спинного мозга образует нервный сегмент - невротом- neurotom. Невротомы яснее выражены там, где существует четкая сегментация в скелете и мышцах.
6.При смещении в процессе эволюции миотомов вслед за ними смещаются иннервирующие их ветви соответствующих невротомов. Например, добавочный нерв выходит из позвоночного канала через череп, а иннервирует плечеголовную, трапециевидную и грудино-челюстную мышцы. В области отхождения нервов в конечности образуются плечевое и пояснично-крестцовое нервные сплетения – plexus brachialis et lumbosacralis- а из них уже берут свое начало нервы, направляющиеся в определенные мышечные группы и кости конечностей. Обычно нервы и мышцы конечностей являются многосегментными. Нервные сплетения встречаются и в области шеи. Соединительные ветви между отдельными нервами указывают на происхождение отдельных нервов из нескольких невротомов.
7.Чувствительные нервы хотя в основном и соответствуют кожным сегментам- дерматомам, но иннервируют не только область своего сегмента, а заходят и в смежные дерматомы. Поэтому обезболивание какого-либо кожного сегмента возможно только при выключении трех смежных невротомов.
Развитие периферического отдела нервной системы
У ланцетника спинномозговые узлы еще не обособлены от спинного мозга. Из-за отсутствия позвонков дорсальный и вентральный нервы отходят от мозга в каждом сегменте самостоятельно. Дорсальный нерв идет через миосепты в кожный покров и дает висцеральную ветвь в кишечную стенку, а вентральный нерв - в миотом, то есть между миосептами. В результате дорсальный и вентральный нервы отходят от мозга на разных уровнях, а не в одной сегментальной плоскости. Лишь с появлением скелетной сегментации у вышестоящих животных оба нерва соединяются в общий ствол, который и отходит от мозга двумя корнями. Одновременно наблюдается обособление на дорсальных корнях спинномозговых узлов.
Черепно-мозговые нервы. Общая характеристика
Черепно-мозговых нервов 12 пар. Они выполняют различные функции. Обонятельный, зрительный и равновесно-слуховой нерв - чисто чувствительные нервы и служат проводящими путями анализаторов - обонятельного, зрительного и равновесно-слухового. Глазодвигательный, блоковый и отводящий нервы являются двигательными для глазных мышц. Добавочный и подъязычный нервы двигательные (для трапециевидной, плечеголовной и грудино-челюстной мышц, а второй для мышц языка и подъязычной кости.). Блуждающий нерв или вагус относится к группе парасимпатических нервов. Из остальных пар тройничный нерв:
а) чувствительный для кожи;
б) двигательный для жевательных мышц.
В нем же проходит часть своего пути секреторного волокна в слезную и слюнную железы. Лицевой нерв двигательный для всех мимических мышц, то есть для губных, щечных, носовых, ушных мышц и мышц век. В нем же проходит часть вкусовых волокон с языка и секреторные волокна для слезной, подчелюстной и подъязычной желез. Языкоглоточный нерв содержит чувствительные волокна со слизистой оболочки корня языка и глотки, вкусовые волокна с корня языка и двигательные волокна для расширителя глотки. В нем же часть своего пути проходят секреторные волокна для околоушной слюнной железы.
От головного мозга акулы отходит 10 пар черепно-мозговых нервов (рис. 26), 11 и 12 пара развита слабо.
I пара – Обонятельный нерв – чувствующий. Иннервирует обонятельную луковицу. Отходит от обонятельных долей переднего мозга.
II пара – Зрительный нерв – чувствующий. Отходит от зрительных бугров промежуточного мозга. Иннервирует сетчатку глаза.
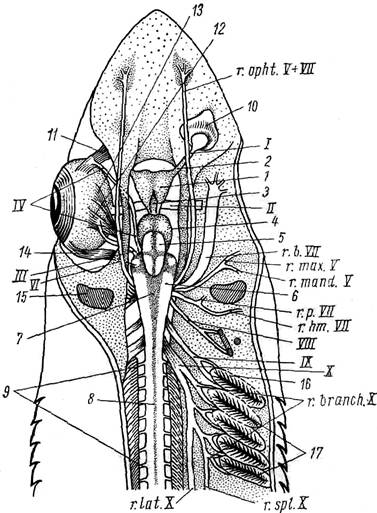
Рисунок 26 – Головной мозг и головные нервы колючей акулы (со спинной стороны, правый глаз удален)
1 – передний мозг; 2 – обонятельная доля переднего мозга;
3 – промежуточный мозг; 4 – средний мозг; 5 – мозжечок;
6 – продолговатый мозг; 7 – ромбовидная ямка; 8 – спинной мозг;
9 – спинномозговые нервы; 10 – обонятельный мешок; 11 – верхняя косая мышца глаза; 12 – верхняя прямая мышца глаза; 13 – внутренняя прямая мышца глаза; 14 – наружная прямая мышца глаза; 15 – брызгальце; 16 – первая жаберная щель; 17 – вторая–пятая жаберные щели;
I – обонятельный нерв; II – зрительный нерв; III – глазодвигательный нерв; IV – блоковый нерв; VI – отводящий нерв; V – тройничный нерв:
г. орht. V – глазничная ветвь, r. mах V – верхнечелюстная ветвь,
r. mаnd. V – нижнечелюстная ветвь; VII – лицевой нерв:
r. орht. VII – глазничная ветвь, r. b. VII – щечная ветвь,
r. р. VII – небная ветвь, r. hm. VII – подъязычная ветвь;
VIII – слуховой нерв; IX – языкоглоточный нерв; Х – блуждающий нерв: r. branch. Х – жаберные ветви, r. lat. X – боковая ветвь,
r. spl. X – внутренностная ветвь.
III пара – Глазодвигательный нерв – двигательный. От дна среднего мозга. Иннервирует мышцы глаза.
IV пара – Блоковый нерв – двигательный. От дна среднего мозга. Иннервирует мышцы глаза.
V пара – Тройничный нерв – смешанный. От боковой поверхности продолговатого мозга. Делится на 3 ветви:
а) глазничная – иннервирует кожу передней части головы;
б) верхнечелюстная – иннервирует зубы и мускулатуру челюстной дуги, слизистую рта;
в) нижнечелюстная – иннервирует зубы и мускулатуру челюстной дуги, слизистую рта.
VI пара – Отводящий нерв – двигательный. От дна продолговатого мозга. Иннервирует наружную прямую мышцу глаза.
VII пара – Лицевой нерв – смешанный. От боковой поверхности продолговатого мозга за тройничным нервом. Делится на 4 ветви:
а) глазничная – идет с аналогичной ветвью тройничного нерва;
б) щечная – кожа головы, слизистая ротовой полости;
в) небная – кожа головы, слизистая ротовой полости, мускулатура подъязычной дуги;
г) подъязычная – кожа головы, слизистая ротовой полости, мускулатура подъязычной дуги.
VIII пара – Слуховой нерв – чувствующий. От продолговатого мозга за лицевым нервом. Иннервирует чувствующий эпителий внутреннего уха.
IX пара – Языкоглоточный нерв – смешанный. Отходит позади слухового нерва. Иннервирует слизистую глотки и ее мускулатуру, 1-ю жаберную дугу.
X пара – Блуждающий нерв – смешанный. От заднебоковой поверхности продолговатого мозга. Делится на ряд ветвей:
а) 4 жаберных – слизистая глотки и мускулатура 2-5 жаберных дуг;
б) внутренностная – сердце, кишечник, легкие, плавательный пузырь;
в) боковая – орган боковой линии (у наземных отсутствует).
XI пара (только у млекопитающих) – Добавочный нерв – двигательный. Образуется из задних корешков блуждающего нерва. Иннервирует мускулатуру плечевого пояса.
XII пара (только у амниот) – Подъязычный нерв – двигательный. От задней части продолговатого мозга. Иннервирует мускулатуру языка и подъязычного аппарата.
У позвоночных имеются 12 пар черепно-мозговых нервов, которые связаны с определенными отделами головного мозга. У низших позвоночных последние нервы могут отходить еще от спинного мозга.
- Обонятельный нерв. Отходит от переднего мозга. Имеет двойную природу. Как правило, образован отростками специализированных нервных клеток, которые тянутся к обонятельной луковице, где происходит переключение на второй нейрон. Если обонятельная луковица далеко отстоит от обонятельной капсулы, то тяж можно считать обонятельным нервом, если луковица за счет выроста головного мозга, поэтому этот нерв по сути представляет собой тракт. Обонятельный нерв представляет собой комплекс коротких чувствующих волокон, идущих от эпителия обонятельного мешка к обонятельной луковице.
- Зрительный нерв. Фактически это зрительный тракт, образующийся как вырост стенки мозга. Он отходит от нижнебоковой поверхности промежуточного мозга и, пройдя через отверстие в стенке глазницы, входит в глазное яблоко, распространяясь по внутренней поверхности сетчатки. Чисто чувствующий нерв. У всех позвоночных животных движения глазного яблока осуществляются при помощи 6 глазных мышц, каждая из которых одним концом прикрепляется к стенке глазницы, а другим к определенному участку поверхности глазного яблока. От передней стенки глазницы отходят 2 косые мышцы глаза: верхняя и нижняя. От задней стенки глазницы пучком отходят 4 прямые мышцы глаза: нижняя прямая, внутренняя прямая, верхняя прямая и наружная прямая.
III. Глазодвигательный нерв. Отходит от дна среднего мозга, пронизывает стенку черепа и делится у основания прямых мышц глаза на ветви, идущие к 4 мышцам: к нижней косой и к нижней, внутренней и верхней прямым.
- Блоковый нерв. В виде тонкой нити отходит от задневерхней части среднего мозга, проходит вперед по крыше среднего моста и, пройдя через переднюю часть стенки глазницы, разветвляется в верхней косой мышце глаза.
- Тройничный нерв. Отходит толстым корнем от переднебоковой поверхности продолговатого мозга и сразу же делится на три ветви: глазничную, верхнечелюстную и нижнечелюстную. Глазничная ветвь вместе с глазничной ветвь лицевого нерва проходит через глазницу и ветвится в переднее части рыла (у акулы), иннервируя кожные органы чувств. Верхнечелюстная и нижнечелюстная ветви начинаются общим стволом и обособляются позади глаза, они иннервируют мышцы челюстной дуги, зубы, кожу головы. По функции тройничный нерв, смешанный: он имеет чисто чувствующие ветви – глазничную и верхнечелюстную, а также смешанную – нижнечелюстную.
- Отводящий нерв. От дна продолговатого мозга сразу уходит в дно черепной коробки. Иннервирует наружную прямую мышцу глаза.
VII. Лицевой нерв. Отходит сразу же за тройничным, тоже распадается на несколько ветвей. Наиболее крупные из них: подъязычная, небная, глазничная и щечная ветви. Подъязычная ветвь проходит позади брызгальца и иннервирует висцеральную дугу с ее слизистой и мускулатурой. Небная отходит вперед от основания предыдущей и ветвится в слизистой оболочке крыши ротовой полости. Глазничная ветвь идет вместе с одноименной тройничного, иннервируя кожные органы боковой линии на спинной поверхности головы. Щечная ветвь аналогичным образом иннервирует брюшную поверхность головы. Отходя крупным стволом позади глазничной ветви, она виде широкого плоского тяжа проходит по дну глазницы. Лицевой нерв – смешанный.
VIII. Слуховой нерв. Идет от боковой поверхности продолговатого мозга и почти сразу же входит в стенку черепной коробки, иннервируя внутреннее ухо. Чисто чувствующий нерв.
- Языкоглоточный нерв. Подходит к первой жаберной щели и здесь делится на две ветви, иннервирующие соответственно переднюю и заднюю поверхности. Задняя ветвь иннервирует всю мускулатуру первой жаберной дуги. Смешанный.
- Блуждающий нерв. Отходит от заднебоковой поверхности продолговатого мозга несколькими корешками, почти сразу же сливающимися в толстый нервный тяж. Иннервирует обширную область тела, основные ветви: четыре жаберные, внутренностная и боковая. 2 жаберные ветви иннервируют жаберные щели от 2 до 5й. Каждая ветвь подойдя к жаберной щели делится на две: переднюю и заднежаберную. Переднежаберная – чувствующая, заднежаберная – смешанная и иннервирует все соответствующую мускулатуру. Внутренностная ветвь идет как продолжение основного ствола блуждающего нерва, входит в брюшную полость и иннервирует внутренние органы. Более поверхностно лежит боковая ветвь, иннервирующая органы боковой линии туловища и хвоста. Блуждающий смешанный.
- Добавочный нерв. Только у млекопитающих приобретает статус отдельного нерва. До млекопитающих ветвь блуждающего нерва, иннервирующая висцеральную трапецевидную мышцу, а затем ее производные.
XII. Подъязычный нерв. Является головным только у амниот. Особого развития достигает у млекопитающих, у которых иннервирует подъязычную мускулатуру и мускулатуру языка. У низших представлен 1-м и 2-м спинномозговыми нервами.
ТЕМА 5. НЕРВНАЯ СИСТЕМА АКУЛЫ
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ОБЪЕКТА
Подтип Позвоночные, Vertebrata
Класс Хрящевые рыбы, Chondrichthyes
Подкласс Пластиножаберные, Elasmobranchii
Отряд Катранообразные, или Колючие акулы, Squaliformes
Представитель — Колючая акула, Squalus acanthias L.
МАТЕРИАЛ И ОБОРУДОВАНИЕ
На одного-двух студентов необходимы:
1. Хранящаяся в 70%-ном спирте голова акулы с отпрепарированным головным мозгом и черепными нервами.
2. Ванночка.
3. Пинцет.
4. Препарировальные иглы — 2.
5. Лупа 4—6 X.
ЗАДАНИЕ
Рассмотреть отделы мозга. Проследить отхождение от мозга черепных нервов и их наиболее крупные ветви. Найти глазные мышцы и подходящие к ним нервы. Сделать следующие рисунки:
1. Мозг акулы с отходящими от него нервами.
2. Глаз с его мышцами и иннервирующими их нервами.
Дополнительное задание
По демонстрационному препарату или модели познакомьтесь со строением стато-акустического органа (внутреннее ухо) акулы.
ОПИСАНИЕ НЕРВНОЙ СИСТЕМЫ
Головной мозг. У акулы головной мозг состоит из пяти отделов. Передний мозг (telencephalon; рис. 26, 1) крупный, нечетко разделен на правую и левую половины (полушария). В передней части полушарий хорошо заметны сильно развитые обонятельные доли (рис. 26, 2).
Рис. 26. Головной мозг и головные нервы колючей акулы
(со спинной стороны, правый глаз удален):
1 — передний мозг, 2 — обонятельная доля переднего мозга, 3 —промежуточный мозг, 4 — средний мозг, 5 — мозжечок, 6 — продолговатый мозг, 7 — ромбовидная ямка, 8 — спинной мозг, 9 — спинномозговые нервы, 10 — обонятельный мешок, 11 — верхняя косая мышца глаза, 12 — верхняя прямая мышца глаза, 13 — внутренняя прямая мышца глаза, 14 — наружная прямая мышца глаза, 15 — брызгальце, 16 — первая жаберная щель, 17 — вторая — пятая жаберные щели; I — обонятельный тракт; II — зрительный нерв; III — глазодвигательный нерв; IV — блоковый нерв; VI — отводящий нерв; V —тройничный нерв: г. opht. V — глазничная ветвь, г. max. V — верхнечелюстная ветвь, г. mand. V — нижнечелюстная ветвь; VII — лицевой нерв: г. opht. VII — глазничная ветвь, г. b. VII — щечная ветвь, r. р. VII — нёбная ветвь, r. hm. VII — подъязычная ветвь; VIII — слуховой нерв; IX — языкоглоточный нерв; X — блуждающий нерв; r. branch. X — жаберные ветви, r. spl. X — внутренностная ветвь, r. lat. X — боковая ветвь
Задняя часть переднего мозга незаметно переходит в промежуточный мозг (diencephalon; рис. 26, 3), который на препарате виден как углубление между вздутиями переднего и среднего мозга.
Средний мозг (mesencephalon; рис. 26, 4) имеет вид крупных парных вздутий (зрительные доли), которые прикрывают собой остальную массу этого отдела мозга.
Четвертый отдел мозга — мозжечок (cerebellum, metencephalon; рис. 26, 5) — у акуловых рыб сильно развит и спереди налегает на средний, а сзади — на продолговатый мозг.
Последний отдел головного мозга — продолговатый мозг (myelencephalon; рис. 26, 6). Сверху в центральной части продолговатого мозга отчетливо видна ромбовидная ямка (рис. 26, 7), представляющая собой полость этого отдела мозга (четвертый желудочек). Она затянута сосудистой оболочкой, которая при препарировке обычно разрушается.
Головные нервы. Характер отхождения головных нервов у акуловых рыб типичен для всех позвоночных животных. От головного мозга у акуловых рыб отходят десять пар головных (черепных) нервов, каждая пара — симметрично с обеих сторон головного мозга. Головные нервы обычно имеют двойное обозначение: порядковый номер и название.
I. Обонятельный нерв (nervus olphactorius). У акуловых рыб, как и у всех позвоночных, от переднего мозга отходят выросты его лобных долей — обонятельные тракты (рис. 26, I). Они образуют у обонятельных капсул расширения — обонятельные луковицы. Обонятельный нерв представляет комплекс коротких чувствующих волокон, идущих от эпителия обонятельного мешка к обонятельной луковице.
II. Зрительный нерв (n. opticus; рис. 26, II). Фактически это зрительный тракт, образующийся как вырост стенки мозга — он отходит от нижне-боковой поверхности промежуточного мозга и, пройдя через отверстие в стенке глазницы, входит в глазное яблоко, распространяясь по внутренней поверхности сетчатки. Нерв чисто чувствующий.
Рис. 27. Глазные мышцы акулы и иннервирующие их нервы :
1 — верхняя косая мышца глаза, 2 — нижняя косая мышца, 3 — нижняя прямая мышца глаза, 4 — внутренняя прямая мышца, 5 — верхняя прямая мышца, 6 — наружная прямая мышца; II — зрительный нерв, III — глазодвигательный нерв, IV — блоковый нерв, VI — отводящий нерв
У всех позвоночных животных движения глазного яблока осуществляются при помощи 6 глазных мышц, каждая из которых одним концом прикрепляется к стенке глазницы, а другим — к определенному участку поверхности глазного яблока. От передней стенки глазницы отходят 2 косые мышцы глаза: верхняя (m. obliquus superior; рис. 27, 1) и нижняя (m. obliquus inferior; рис. 27, 2). От задней стенки глазницы пучком отходят 4 прямые мышцы глаза: прямая нижняя (m. rectus inferior; рис. 27, 3), прямая внутренняя (m. rectus internus; рис. 27, 4), прямая верхняя (m. rectus superior; рис. 27, 5) и прямая наружная (т. rectus externus; рис. 27, 6). Эти мышцы иннервируются тремя парами головных нервов — III, IV и VI. Все эти нервы чисто двигательные.
III. Глазодвигательный нерв (n. oculornotorius; рис. 26, III). Отходит от дна среднего мозга (его видно, если слегка отодвинуть средний мозг от стенки черепа), пронизывает стенку черепа и делится у основания прямых мышц глаза на ветви, идущие к четырем мышцам: к нижней косой и к нижней, внутренней и верхней прямым (рис. 27, III).
IV. Блоковый нерв (n. trochlearis; рис. 26, IV). В виде тонкой нити отходит от задне-верхней части среднего мозга (на препарате он виден выходящим из-под мозжечка), проходит вперед по крыше среднего моста и, пройдя через переднюю стенку глазницы, разветвляется в верхней косой мышце глаза (рис. 27, IV).
Все остальные головные нервы отходят от продолговатого мозга.
VI. Отводящий нерв (n. abducens; рис. 26, VI). От дна продолговатого мозга сразу уходит в дно черепной коробки. На препарате отхождение этого нерва не видно. Он иннервирует наружную прямую мышцу глаза, на внутренней поверхности которой хорошо видны его ветви (рис. 27, VI).
V. Тройничный нерв (n. trigeminus; pис. 26, V). Это нерв сложный. Он отходит толстым корнем от переднебоковой поверхности продолговатого мозга и сразу же делится на три ветви: глазничную, верхнечелюстную и нижнечелюстную.
Глазничная ветвь (ramus ophthalmicus; рис. 26, r. opht. V) вместе с глазничной ветвью лицевого нерва (см. ниже — VII) проходит через глазницу и ветвится в передней части рыла, иннервируя кожные органы чувств.
Верхнечелюстная (r. maxillaris; рис. 26, г. max. V) и нижнечелюстная (r. mandibularis; рис. 26, r. mand. V) ветви начинаются общим стволом и обособляются позади глаза; они иннервируют мышцы челюстной дуги, зубы, слизистую оболочку ротовой полости и т.д.
По функции тройничный нерв смешанный: он имеет чисто чувствующую ветвь — глазничную и смешанные (чувствующие и двигательные) — верхнечелюстную и нижнечелюстную.
VII. Лицевой нерв (n. facialis; рис. 26, VII). Отходит сразу же за тройничным (на препарате их основания трудно разграничить). Этот нерв также распадается на несколько ветвей. Наиболее крупные и них — глазничная, щечная, нёбная и подъязычная ветви.
Глазничная ветвь (r. ophtalmicus; рис. 26, r. opht. VII) идет вместе с одноименной ветвью тройничного нерва.
Позади глазничной ветви отходит крупный ствол — щечная ветвь (r. buccalis; рис. 26, r. b. VII). Она в виде широкого тяжа проходит по дну глазницы, иннервируя кожные органы чувств брюшной поверхности головы.
Далее общим стволом отходят две ветви: нёбная и подъязычная.
Нёбная ветвь (r. palatinus; рис. 26, r. р. VII) сразу же после обособления уходит в дно черепной коробки и затем ветвится в слизистой оболочке ротовой полости.
Подъязычная ветвь (r. hyomandibularis; рис. 26, r. hm. VII) проходит позади брызгальца (рис. 26, 15) и иннервирует мускулатуру подъязычной дуги и кожные органы чувств боков головы.
Лицевой нерв, как и тройничный, смешанный: его глазничная, щечная и нёбная ветви чувствующие, а подъязычная — смешанная (чувствующая и двигательная).
VIII. Слуховой (n. acusticus; рис. 26, VIII). Идет от боковой поверхности продолговатого мозга и почти сразу же входит в стенку черепной коробки, иннервируя внутреннее ухо. Это чисто чувствующий нерв.
IX. Языкоглоточный нерв (n. glossopharyngeus; рис. 26, IX). Подходит к первой жаберной щели и здесь делится на две ветви, иннервирующие соответственно ее переднюю и заднюю поверхности. Задняя ветвь иннервирует всю мускулатуру первой жаберной дуги. Этот нерв по своей функции смешанный: он имеет и чувствующие, и двигательные волокна.
X. Блуждающий нерв (n. vagus, рис. 26, X). Отходит от задне-боковой поверхности продолговатого мозга несколькими корешками, почти сразу же сливающимися в толстый нервный тяж. В отличие от других головных нервов иннервирует обширную область тела, его основные ветви — четыре жаберные, внутренностная и боковая.
Четыре жаберные ветви (r. branchialis; рис. 26, r. branch. X) иннервируют жаберные щели от второй до пятой. Каждая ветвь, подойдя к жаберной щели, делится на две: передне- и заднежаберную. Переднежаберная ветвь чувствующая, а заднежаберная — смешанная и иннервирует всю мускулатуру соответствующей жаберной дуги (от второй до пятой).
Внутренностная ветвь (r. splanchnicus; рис. 26, r. spl. X) идет как продолжение основного ствола блуждающего нерва. Она входит в брюшную полость, иннервируя внутренние органы.
Более поверхностно лежит боковая ветвь (r. lateralis; рис. 26, r. lat. X), иннервирующая органы боковой линии туловища и хвоста.
Блуждающий нерв смешанный (включает и чувствующие, и двигательные волокна).
У высших позвоночных животных обособляются еще две пары головных нервов.
XI. Добавочный нерв (n. accessorius) имеется только у млекопитающих.
XII. Подъязычный нерв (n. hypoglossus) впервые четко обособляется у пресмыкающихся.
Зачатки этих нервов есть у акуловых рыб в виде нервных корешков, отходящих позади блуждающего нерва в области затылочной стенки мозгового черепа.
Спинномозговые нервы (n. spinalis; рис. 26, 9) видны на препарате — они посегментно отходят от спинного мозга (medulla spinalis). Иннервируют соответствующие сегменты тела и по своей функции представляют собой смешанные нервы, состоящие из чувствующих и двигательных волокон.
Внутреннее ухо (стато-акустический орган). У хрящевых рыб состоит из перепончатого лабиринта, заключенного в хрящевую слуховую капсулу — боковое расширение хрящевого черепа позади глазницы. Три изогнутые трубочки — полукружные каналы — лежат в трех взаимно перпендикулярных плоскостях (рис. 28) и концами открываются в преддверье, или перепончатый мешок.
Рис. 28. Схема перепончатого лабиринта акулы :
1 — полукружные каналы, 2 — овальный мешочек, 3 — круглый мешочек, 4 — лагена
Он подразделяется на две части: верхнюю — овальный мешочек и более крупную нижнюю — круглый мешочек, от которого отходит небольшой вырост — лагена. Полость перепончатого лабиринта заполнена эндолимфой, в которой взвешены мелкие кристаллики — отоконии. В полости круглого мешочка обычно находятся более крупные известковые образования — отолиты. Окончания слухового нерва подходят к отдельным участкам перепончатого лабиринта, покрытым чувствующим эпителием — слуховым пятнам и слуховым гребням. Перепончатый лабиринт — орган чувства равновесия и орган слуха.
ЗАКЛЮЧЕНИЕ
Хрящевые рыбы — новый этап эволюции позвоночных животных. По сравнению с круглоротыми характерен более высокий уровень организации и более активные формы жизнедеятельности.
Наиболее существенное отличие этого класса (и всех челюстноротых - Gnathostomata) от бесчелюстных (Agnatha) в том, что висцеральный отдел черепа челюстноротых состоит из нескольких пар расчленных подвижных висцеральных дуг: челюстной (хватательная функция), подъязычной (функция подвижного прикрипления челюстной дуги к осевому черепу) и нескольких (у хрящевых рыб пяти) жаберных дуг (функция опоры дыхательного аппарата и обеспечения дыхательных движений). Такое устройство висцерального скелета обеспечивает интенсификацию дыхания и значительно облегчает схватывание добычи, тем самым способствуя повышению активности питания и в конечном результате повышению общего уровня обмена веществ.
У хрящевых рыб имеются и другие прогрессивные по сравнению с круглоротыми особенности строения. Появление парных конечностей способствовало увеличению скорости и особенно устойчивости и маневренности плавания. Замена (хотя и неполная) хорды хрящевыми позвонками привела к усилению опорной функции осевого скелета при сохранении его подвижности (способность к изгибам), что также способствовало активизации передвижения, поскольку основным локомоторным органом становится хвостовой отдел тела. Возрастанию скорости плавания способствует и увеличение размеров хвостового плавника.
Резко возрастают у хрящевых рыб относительные размеры головного мозга, все отделы которого хорошо развиты. Органы чувств (глаза, органы обоняния и слуха, сейсмосенсорные органы боковой линии) развиты значительно лучше, чем у круглоротых. Все это также обеспечивает более активный образ жизни. Черепная коробка хрящевых рыб уже закрывает головной мозг со всех сторон; капсулы органов чувств срастаются со стенками мозговой коробки, образуя цельный прочный осевой череп.
Челюстноротым (в том числе и хрящевым рыбам) свойствен иной, нежели у круглоротых, тип строения органов дыхания. Эктодермальные по происхождению жабры прикрепляются к подвижным жаберным дугам и состоят из большого количества тонких жаберных лепестков. Вода непрерывно омывает поверхность жаберных лепестков, что облегчает насыщение крови кислородом. Более отчетливая дифферен-цировка пищеварительного тракта на отделы (как морфологическая, так и функциональная), увеличение поверхности всасывания благодаря имеющемуся в толстой кишке хорошо развитому спиральному клапану, сильное развитие желез пищеварительной системы (печень, поджелудочная железа), вырабатывающих различные пищеварительные ферменты, — все это способствует повышению скорости и эффективности пищеварения.
Вся совокупность прогрессивных изменений, охватывающих различные системы органов, обеспечивает более высокий уровень обмена веществ, общую активизацию жизнедеятельности хрящевых рыб по сравнению с низшими позвоночными (круглоротыми) и появление в этой группе большего разнообразия жизненных форм.
Однако по сравнению с эволюционно бэлее молодым классом костных рыб многие особенности строения хрящевых рыб выглядят примитивными и ограничивают возможности широкой адаптивной радиации. Массивный хрящевой скелет увеличивает массу тела. Пояса парных плавников лежат в толще мускулатуры и не укреплены на осевом скелете. Все это ограничивает возрастание подвижности. Массивные хрящевые челюсти способны лишь к однообразным простым хватательным движениям. Приспособительные изменения, связанные с различным характером питания акул и скатов, ограничиваются различиями в строении зубов.
По сравнению с костистыми рыбами для хрящевых рыб характерен более низкий уровень обмена веществ, что определяется рядом морфологических и физиологических особенностей: еще довольно простым строением дыхательного аппарата (отсутстви жабе рной крышки), меньшей дифференцировкой и длиной кишечника, меньшей скоростью кровотока и кислородной емкостью крови и т. д. Все это находит свое выражение и в ограниченном распространении хрящевых рыб (только обитатели моря), и в малом разнообразии их жизненных форм при ограниченном числе видов (около 600 против почти 20 тыс. видов костных рыб).
Но в море и сейчас хрящевые рыбы могут считаться вполне процветающей группой, успешно отстаивающей свои жизненные ниши и конкурирующей с костными рыбами. Этому способствуют некоторые своеобразные особенности хрящевых рыб. Прежде всего для них характерна большая живучесть, чем у костных рыб. Она проявляется в способности к длительному голоданию, в возможности переносить очень большие повреждения и ранения, в большей мощи мускулатуры (в эксперименте акула заметно не снижает скорости движения, если к ней привязан дополнительный груз, составляющий до 25% массы ее тела; многие костистые рыбы почти полностью теряют способность двигаться при дополнительной нагрузке в 5—10% от массы тела). Эти особенности в той или иной степени связаны с относительно большим размером головного мозга и большей (по сравнению с костными рыбами) его дифференцировкой (сильное развитие переднего мозга и мозжечка и др.), с повышенной автономностью периферической нервной системы, тем что в мышечных сегментах имеется значительное количество дыхательного пигмента — миоглобина, и другими физиологическими и биохимическими особенностями.
Специфические черты в строении половой системы, обеспечивающие возможность внутреннего оплодотворения, относительно крупные размеры яиц с большим запасом питательных веществ, их прочная рогоподобная скорлупа резко снижают эмбриональную смертность. Еще более повышает эффективность размножения свойственное многим видам хрящевых рыб яйцеживорождение. Эти особенности снижают расход энергии при размножении и повышают выживаемость акуловых рыб в борьбе за существование.
Очень своеобразен у хрящевых рыб механизм осморегуляции. Осмотическое давление в тканях и крови хрящевых рыб примерно равно осмотическому давлению морской воды. Этот изотонизм обеспечивается высоким содержанием мочевины в крови и тканях. При изменениях солености среды меняется уровень мочевины в крови и тканях, приводя осмотическое давление жидкостей тела в соответствие с таковым среды. Концентрация же биологически важных солей в организме остается при этом неизменной. Такой тип осморегуляции свойствен только хрящевым рыбам.
Хрящевые рыбы — наиболее древняя группа рыб, для которых характерны многие примитивные черты организации. Они сохранились до наших дней благодаря выработке своеобразных черт организации, частично описанных выше, помогающих им успешно выдерживать конкуренцию с костными рыбами — эволюционно более молодой и прогрессивной группой, имеющей более высокий сбщий уровень организации. Хрящевые и костные рыбы — это два самостоятельных конвергентных типа развития водных позвоночных животных.
Дополнительная литература
Гуртовой Н. Н., Матвеев Б. С, Дзержинский Ф. Я. Практическая зоотомия позвоночных. Низшие хордовые, бесчелюстные, рыбы. М., 1976.
Никольский Г. В. Частная ихтиология. М., 1971.
Никольский Г. В. Экология рыб. М., 1974.
Суворов Е. К. Основы ихтиологии М., 1948.
Шмальгаузен И. И. Основы сравнительной анатомии позвоночных. М., 1947.
Читайте также:
