
Характеристика торможения как одного из нервных процессов
Общая характеристика функций нервной системы.
Деятельность нервной системы складывается из двух процессов: возбуждения и торможения.
Характеристика процессов возбуждения в ЦНС.
Возбуждение в ЦНС осуществляется благодаря активности возбуждающих синапсов. Возбуждающий медиатор взаимодействует с хеморецепторами постсинаптической мембраны. В результате на постсинаптической мембране возникает ВПСП, связанный с увеличением проницаемости главным образом ионов натрия (возможно кальция). При этом выходящий ток калия несколько снижает деполяризацию, вызванную натрием.
Последовательность процессов в постсинаптической мембране.
Нейромедиатор взаимодействует с рецептором, активирует мембранные ферменты аденилатциклазу и гуанилатциклазу. Это приводит к увеличению образования циклической АМФ и ГМФ, которые вызывают фосфорилирование мембранных белков, что приводит к открытию хемовозбудимых каналов для натрия, калия и кальция. Под влиянием ВПСП в аксонном холмике развивается потенциал действия.
Проявление активности нейронов. Различают спонтанную и вызванную активность.
Спонтанная активность – это самопроизвольное возбуждение нейронов (автоматия). Различают нерегулярную активность, когда импульсы в нейроне возникают через различные промежутки времени; взрывную – возникает серия частых импульсов; Регулярную – с высокой частотой.
Вызванная активность возникает в ответ на поступление информации от барорецепторов (при изменении давления, от проприорецепторов при изменении тонуса мышц, от хеморецепторов при изменении состава микросреды, от осморецепторов при изменении осмотического давления). В результате возникает активность ранее молчавших нейронов или изменение активности уже работающих нейронов.
Торможение в ЦНС.
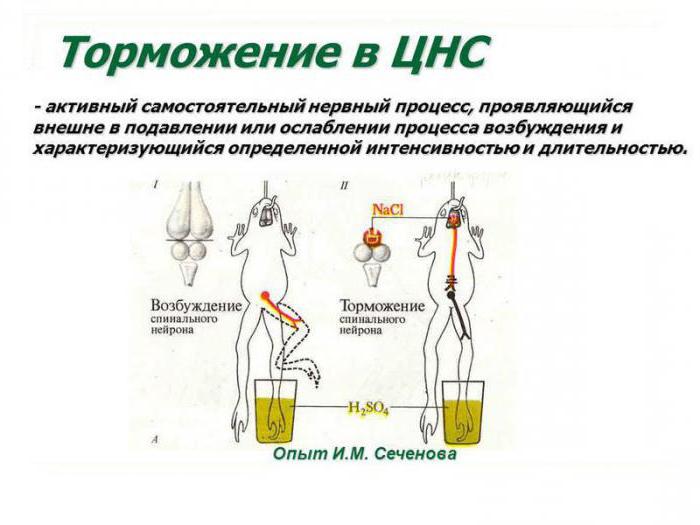
Явление центрального торможения было открыто в 1863 году Иваном Михайловичем Сеченовым. Он обнаружил торможение рефлексов спинного мозга при раздражении зрительного бугра. Было установлено, что есть специальные структуры, вызывающие торможение рефлексов. В дальнейшем было показано, что торможение рефлекторной деятельности одной стороны тела может возникнуть при возбуждении противоположной. Например, возбуждение сгибателя правой ноги вызывает торможение сгибателя левой ноги (работы Введенского, Шеррингтона).
Торможение это не утомление и не перевозбуждение. Это самостоятельный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения.
Торможение проявляется в форме локального процесса и поэтому всегда связано с наличием тормозных синапсов. Такие синапсы образуются аксонами специальных тормозных нейронов, угнетающих активность всех нервных клеток, с которыми они связаны.
Различают пресинаптическое и постсинаптическое торможение. Постсинаптическое подразделяется на прямое, возвратное и латеральное.
Характеристика торможения в ЦНС
Пресинаптическое торможение – это вид торможения развивается за счет процессов, приводящих к угнетению освобождению медиатора из пресинаптических окончаний и снижению эффективности синаптической передачи или полному ее прекращению. Пресинаптическое торможение обнаружено в мозговом стволе и, особенно в спинном мозге. Оно обусловлено наличием вставочных тормозных нейронов, к которым подходят коллатерали афферентных волокон. Аксон тормозного нейрона образует синапс на пресинаптических терминалах возбуждающей клетки (аксо-аксональный синапс).
Механизм пресинаптического торможения. В случае избыточного притока сенсорной информации с рецепторов происходит активация тормозных интернейронов. Тормозной синапс высвобождает ГАМК (гамма-аминомасленную кислоту), которая вызывает стойкую деполяризацию пресинаптической терминали. Это приводит к снижению амплитуды ПД, поступающего на пресинаптическую терминаль. Следствие этого – снижение квантовой секреции медиатора из нервных терминалей возбуждающего нейрона. При этом на мотонейроне регистрируется снижение амплитуды ВПСП, что снижает вероятность возникновения потенциала действия.
Постсинаптическое торможение осуществляется за счет процессов, происходящих на постсинаптической мембране.
Механизм постсинаптического торможения. В тормозном синапсе выделяются тормозные медиаторы (например – глицин), которые взаимодействуют с рецепторами постсинаптической мембраны. Это приводит к увеличению проницаемости для калия и хлора и к гиперполяризации постсинаптической мембраны. При этом возбудимость нейрона снижается и снижается вероятность ответа на приходящий сигнал.
Воздействия на процесс торможения. Процесс торможения можно блокировать, воздействуя на различные этапы передачи в тормозном синапсе.
1)Столбнячный токсин нарушает высвобождение тормозных медиаторов (пресинаптический уровень воздействия).
2) Стрихнин конкурирует с тормозным медиатором за рецептор на постсинаптической мембране (постсинаптический уровень воздействия).
Функционирование условно рефлекторного механизма базируется на двух основных нервных процессах: возбуждения и торможения. Достаточно сильное раздражение органа приводит его в активное деятельное состояние - возбуждение.
Возбуждение - свойство живых организмов, активный ответ возбудимой ткани на раздражение. Основная функция нервной системы, направленная на реализацию того или иного способа активации организма. Оно проявляется в мгновенных и существенных сдвигах в процессах обмена веществ, то есть может происходить только в живых клетках. Первый и притом обязательный признак возникшего возбуждения - электрическая реакция на результат изменений электрического заряда поверхностной мембраны клеток. Затем наступает специфическая для каждого органа реакция, чаще всего выражающаяся во внешней работе: мышца сокращается, железа выделяет сок, в нервной клетке возникает импульс.
Возбудимость, то есть способность в ответ на раздражение приходить в состояние возбуждения, - одно из основных свойств живой клетки. Исчезновение возбудимости означает прекращение рабочих функций, а в конечном счете, и жизни.
Вызвать состояние возбуждения можно различными раздражителями, например механическими (укол булавкой, удар), химическими (кислота, щелочь), электрическими. Наименьшая сила раздражения, достаточная для того, чтобы вызвать минимальное возбуждение, называется порогом раздражения.
По мере укрепления условного рефлекса происходит усиление тормозного процесса.
Торможение - активный, неразрывно связанный с возбуждением процесс, приводящий к задержке деятельности нервных центров или рабочих органов. В первом случае торможение называется центральным, во втором -периферическим.
В зависимости от природы физиологического механизма, лежащего в основе тормозного эффекта на условнорефлекторную деятельность организма, различают безусловное (внешнее и запредельное) и условное (внутреннее) торможение условных рефлексов.
Торможение безусловное - разновидность коркового торможения. В отличие от условного торможения наступает без предварительной выработки. Включает в себя: 1) индукционное (внешнее) торможение; 2) запредельное (охранительное) торможение.
Внешнее торможение условного рефлекса возникает под действием другого постороннего условного или безусловного раздражителя. Когда под влиянием какого-нибудь изменения внешней или внутренней среды в коре больших полушарий возникает достаточно сильный очаг возбуждения, то вследствие отрицательной индукции, возбудимость других ее пунктов оказывается пониженной - в той или иной степени в них развивается тормозное состояние.
Индукционное (внешнее) торможение - экстренное прекращение условнорефлекторной деятельности под воздействием посторонних стимулов, биологическое значение его - преимущественное обеспечение ориентировочной реакции на неожиданно возникший раздражитель. Примером такого торможения может служить следующий опыт.
У собаки выработан прочный условный рефлекс на свет электрической лампочки. Величина реакции - 10 капель слюны за 30 секунд изолированного действия раздражителя. Включение одновременно с зажиганием лампочки нового раздражителя (звонка) привело к уменьшению условного рефлекса до 1-2 капель. На повторное действие света (без включения звонка) выделилось 7 капель слюны. Испробованный еще через несколько минут условный рефлекс на свет лампочки полностью восстановился. Таким образом, под влиянием нового постороннего раздражителя произошло торможение условного рефлекса, продолжавшееся в течение некоторого времени. Источником индукционного торможения могут оказаться и раздражения, возникающие в самом организме, например сдвиги в работе пищеварительного аппарата.
При повторном действии одного и того же постороннего раздражителя вызываемый им очаг возбуждения постепенно слабеет, явление индукции исчезает и в результате прекращается тормозящее влияние на условные рефлексы.
Если изолированное действие условного пищевого раздражителя, обычно подкрепляемого через 20 секунд, продолжать 2-3 минуты, выделение слюны прекратится. То же произойдет при чрезмерном усилении раздражения. Секреция прекращается в результате развившегося торможения. Это можно доказать, пробуя также другие условные раздражители. Примененные сразу же после удлиненного во времени или чрезмерно сильного раздражения, они вызывают слабую рефлекторную реакцию вследствие иррадиации тормозного процесса на другие клетки коры.
Торможение, развивающееся в корковой клетке под влиянием длительного или сверхсильного раздражения, Павлов назвал запредельным.
Запредельное (охранительное) торможение - торможение, возникающее при действии стимулов, возбуждающих соответствующие корковые структуры выше присущего им предела работоспособности, и обеспечивающее тем самым реальную возможность ее сохранения или восстановления.
Как индукционное, так и запредельное торможение свойственно не только коре больших полушарий, но и всем другим отделам нервной системы. Существует, однако, вид торможения, встречающийся только в высшем отделе головного мозга. Такое специфическое корковое торможение Павлов назвал условным или внутренним.
Условное (внутреннее) торможение условного рефлекса носит условный характер и требует специальной выработки. Биологический смысл его в том, что изменившиеся условия внешней среды требуют соответствующего адаптивного приспособительного изменения в условнорефлекторном поведении.
При выработке обычного условного рефлекса устанавливается связь раздражаемого пункта с другим возбужденным пунктом коры. При выработке условного торможения действие раздражителя связывается с тормозным состоянием корковых клеток. Один и тот же раздражитель в зависимости от того, с каким состоянием коры связывается его действие, может привести к образованию либо условного рефлекса, либо условного торможения. В первом случае он станет положительным условным раздражителем, а во втором -отрицательным.
Развитие торможения легко обнаружить в эксперименте. Так, у собаки предварительно были выработаны прочный условный рефлекс на удары метронома с подкреплением через 3 минуты и условные рефлексы на другие раздражители с подкреплением через 30 секунд. Затем метроном пускался в ход на 1 минуту и тотчас заменялся другим раздражителем, подкрепляемым через 30 секунд. При такой постановке опыта эффект действия второго раздражителя оказывался резко сниженным, то есть заторможенным. Очевидно, торможение, развившееся под влиянием, одноминутного действия метронома, захватило и другие участки коры.
Различают четыре вида внутреннего торможения: угасание, дифференци-ровка, условный стимул, запаздывание.
Если условный раздражитель предъявляется без подкрепления безусловным, то через некоторое время после изолированного применения условного стимула реакция на него угасает. Такое торможение условного рефлекса называется угасательным (угасание). Угасание условного рефлекса - это временное торможение, угнетение рефлекторной реакции. Спустя некоторое время новое предъявление условного стимула без подкрепления его безусловным вначале вновь приводит к проявлению условнорефлекторной реакции.
Если у животного или человека с выработанным условным рефлексом на определенную частоту звукового стимула (например, звука метронома с частотой 50 в секунду) близкие по смыслу раздражители (звук метронома с частотой 45 или 55 в секунду) не подкреплять безусловным стимулом, то условно рефлекторная реакция на последние угнетается, подавляется. Такой вид внутреннего (условного) торможения называется дифференцировочным торможением (дифференцировка). Дифференцировочное торможение лежит в основе многих форм обучения, связанных с выработкой тонких навыков.
Если условный стимул, на который образован условный рефлекс, применяется в комбинации с некоторым другим стимулом и их комбинация не подкрепляется безусловным стимулом, наступает торможение условного рефлекса, вызываемого этим стимулом. Этот вид условного торможения называется условным тормозом.
Запаздывательное торможение - торможение, наступающее тогда, когда подкрепление условного сигнала безусловным раздражителем осуществляется с большим опозданием (2-3 мин.) по отношению к моменту предъявления условного раздражителя.
В лечебных целях, а также в физиологическом эксперименте чаще всего применяют в качестве раздражителя электрическую энергию. Преимущество ее заключается в том, что, правильно дозированная, она не вызывает в ткани необратимых изменений и может применяться повторно. Нетрудно точно отградуировать как интенсивность и длительность раздражения, так и скорость его возникновения и исчезновения.
Виды торможения
Дата добавления: 2018-05-12 ; просмотров: 734 ;
С точки зрения физиологических механизмов, лежащих в основе торможения безусловных и условных рефлексов в ЦНС, в настоящее время принято различать первичное и вторичное торможение. Первичное торможение является результатом активации особых тормозящих структур (тормозных, или тормозящих нейронов), действующих на тормозимую клетку и вызывающих в ней торможение как первичный процесс без предварительного возбуждения. К первичному торможению относятся пресинаптическое и постсинаптическое торможение (разновидностью постсинаптического являются реципрокное торможение, возвратное, или антидромное, торможение и латеральное). Вторичное торможение возникает в клетке без действия на нее специфических тормозящих структур, а как следствие ее возбуждения, т.е. вторично. Ко вторичному торможению относят пессимальное торможение и торможение вслед за возбуждением. Среди процессов торможения в коре больших полушарий, в результате которых временно прекращается реализация условных рефлексов, принято выделять внешнее торможение, пессимальное торможение и внутреннее, или условное, торможение (дифференцировочное, угасательное, запаздыавающее, условно-тормозное). О процессах торможения в коре больших полушарий более подробно излагается в главе 34 (Торможение условных рефлексов).
Рассмотрим тормозные нейроны в ЦНС. Любой вид первичного торможения осуществляется с участием специализированных вставочных нейронов, или интернейронов, которые получили название тормозных нейронов. Часть этих нейронов называются по имени автора, описавшего соответствующий тип (например, клетки Реншоу, клетки Уилкинсона, грушевидные клетки Пуркинье в мозжечке, звездачатые клетки коры и другие). Их аксоны образуют контакты (аксо-дендритные, аксо-соматические) непосредственно с тормозимой клеткой, либо аксоны тормозных клеток взаимодействуют с аксоном возбуждающего нейрона, который направляется к тормозимому нейрону - в этом случае образуется аксо-аксональный синапс, который блокирует проведение возбуждения по аксону, и возбуждающий сигнал не доходит до тормозимого нейрона. Тормозные нейроны имеются во всех отделах ЦНС, но особенно их много в спинном мозге, в мозжечке, в базальных ядрах и в коре больших полушарий. Отметим еще раз, что на долю вставочных нейронов приходится до 97-99% всех нейронов мозга. Даже если только часть этих нейронов является тормозными, то и в этом случае можно утверждать, что тормозные нейроны составляют достаточно большую нейронную популяцию. Трагические последствия нарушения процессов торможения в ЦНС (как это, например, имеет место при столбняке) еще раз доказывают важность тормозных процессов в деятельности ЦНС, в том числе для выполнения координационной ее деятельности.
Постсинаптическое, или координационное, торможение как вариант первичного торможения. Этот вид торможения развивается в основном в аксо-дендритных синапсах. Наличие в нервной системе тормозящих синапсов было показано Ллойдом, Реншоу, Уилкинсоном. Основой постсинаптического торможения является гиперполяризация постсинаптической мембраны тормозимого нейрона, представляющая собой тормозный постсинаптический потенциал (ТПСП). В качестве медиатора постсинаптического торможения могут выступать гамма-аминомасляная кислота (ГАМК), глицин, норадреналин, дофамин, серотонин, эндогенные бензодиазепины (эндозепины), эндогенные опиоиды (эндорфин, энекефалин и др.), фактор J (Флори), некоторые аминокислоты и пептиды. Все они приводят к повышению проницаемости постсинаптической мембраны тормозимого нейрона к ионам К + и Сl - , в результате чего возникает ТПСП. Важным условием эффективности тормозного процесса является наличие на тормозимом нейроне соответствующих рецепторов, способных воспринять тормозной медиатор. Исследование механизмов генерации ТПСП первоначально было подробно проанализировано в опытах на альфа-мотонейронах спинного мозга, а в последующем изучено в отношении нейронов ретикулярной формации ствола мозга, нейронов коры больших полушарий, мозжечка и таламических ядер теплокровных животных.
Основными тормозными медиаторами являются ГАМК и глицин.
ГАМК-ергические тормозные нейроны очень широко представлены в ЦНС, особенно их много в коре большого мозга. Эффект ГАМК-ергических тормозных нейронов может быть заблокирован бикукулином (специфический блокатор ГАМКА-рецепторов) и факлофеном (блокатор FAMKБ - рецепторов). В тоже время ряд факторов повышают эффективность тормозного действия ГАМК. Среди них - селективный агонист ГАМКБ - рецепторов баклофен, а также позитивные модуляторы ГАМКА-рецепторов - барбитураты, бензодиазепины и этанол. Вот почему применение указанных веществ, в том числе барбитуратов приводит к развитию сна.
Глицинергические тормозные нейроны в основном представлены в стволе мозга и в спинном мозге. Глицин вызывает гиперполяризацию за счет повышения проницаемости хлорных каналов. Специфическим блокатором глициновых рецепторов является стрихнин, а также столбнячный токсин. Вот почему при столбняке происходит выраженная иррадиация возбуждения по ЦНС, наличию которой препятствует глицинергические нейроны.
Поскольку на постсинаптической мембране одного и того же нейрона взаимодействуют от многих синаптических входов множественные ВПСП и ТПСП, обеспечивая интегративную деятельность нейрона, постсинаптическое торможение получило название координационного.
Структурной основной пресинаптического торможения являются аксо-аксональные синапсы, образованные терминалями аксонов тормозных интернейронов и аксонными окончаниями возбуждающих нейронов. При этом окончание аксона тормозного нейрона является пресинаптическим по отношению к терминали возбуждающего нейрона, которая оказывается постсинаптической по отношению к тормозному окончанию и пресинаптической по отношению к активируемой им нервной клетки. В этих аксо-аксональных синапсах под влиянием медиатора тормозного нейрона развивается деполяризация, которая вызывает (подобно катодической депрессии Вериго) уменьшение амплитуды потенциала действия, приходящего в возбуждающее окончание аксона. В результате происходит угнетение процесса высвобождения медиатора возбуждающими нервными окончаниями и снижение амплитуды возбуждающего постсинаптического потенциала. Благодаря этому возникает частичная или полная блокада проведения возбуждения к нервным окончаниям, образующим синапсы на данном (тормозимом) нейроне. Кроме того, пресинаптическое торможение может реализоваться за счет гиперполяризации пресинаптических терминалей под влиянием специфического тормозного медиатора, например, ГАМК, что также препятствует проведению импульса от возбуждающего нейрона к тормозимому нейрону. Известно, что пресинаптическое торможение, вызываемое с участием ГАМК, блокируется с помощью бикуку-лина, специфического блокатора ГАМКА-рецепторов. Это означает, что свое тормозное гиперполяризующее действие ГАМК оказывает за счет повышения проницаемости плазмолеммы аксона для ионов С1 - .
Считается, что пресинаптическое торможение чаще всего наблюдается в стволе мозга и в спинном мозге.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
В начале образования положительного условного рефлекса происходит распространение возбуждения из непосредственного пункта раздражения в коре мозга на другие отделы. Такое распространение И.П.Павлов назвал иррадиацией возбудительного процесса. При иррадиации в процесс возбуждения вовлекаются соседние нервные клетки по отношению к группе клеток, непосредственно возбужденных пришедшими сигналами. Распространение происходит по ассоциативным нервным волокнам коры, которые соединяют рядом расположенные клетки. В иррадиации возбуждения могут участвовать также подкорковые образования и ретикулярная формация.
По мере замедления условного рефлекса возбуждение сосредоточивается все в более ограниченной зоне коры, к которой адресовано раздражение. Это явление носит название концентрации возбудительного процесса. В случае выработки дифференцировоч-ного торможения, оно и ограничивает иррадиацию возбуждения.
И.П.Павлов считал, что торможение также способно к иррадиации и концентрации. Торможение, возникшее в анализаторе при использовании отрицательного условного раздражителя, иррадиирует по коре головного мозга, но в 4-5 раз медленнее (от 20 сек до 5 мин), чем возбуждение. Еще медленнее происходит концентрация торможения. По мере повторения и закрепления отрицательного условного рефлекса время концентрации торможения укорачивается и торможение сосредоточивается в ограниченной зоне коры.
При положительной индукции в клетках, смежных с теми, где только что вызывалось торможение, после прекращения действия тормозного сигнала возникает состояние повышенной возбудимости. Вследствие этого импульсы, поступающие к нейронам при действии положительного раздражителя, вызывают повышенный эффект. При отрицательной индукции в клетках коры, окружающих возбужденные нейроны, возникает процесс торможения.
Отрицательная индукция ограничивает иррадиацию процесса возбуждения в коре мозга. Отрицательной индукцией можно объяснить торможение условных рефлексов более сильными посторонними раздражениями (внешнее безусловное торможение). Такое сильное раздражение вызывает в коре мозга интенсивное возбуждение нейронов, вокруг которых появляется широкая зона торможения нейронов, захватывающая клетки, возбужденные условным раздражителем.
Явления отрицательной и положительной индукции в коре головного мозга подвижны, постоянно сменяют друг друга. В разных пунктах коры мозга одновременно могут возникать очаги возбуждения и торможения, положительной и отрицательной индукции.
У человека синтетическая деятельность коры не ограничивается лишь формированием временных связей между корковыми представительствами безусловных рефлексов и центрами органов чувств. Существенное значение имеет образование временных связей между центрами, участвующими в восприятии комплексных и последовательных раздражении.
Аналитическая деятельность коры головного мозга заключается в дифференцировании по характеру и интенсивности массы раздражении, доходящих в форме сигналов до мозговой коры, что достигается с помощью внутреннего торможения, позволяющего точно дифференцировать раздражители по их биологической значимости. Анализ внешних и внутренних воздействий в организме начинается с момента их действия на рецепторы. По пути к корковым нейронам афферентные сигналы проходят ряд образований центральной нервной системы, где происходит их элементарный анализ. Высший же анализ осуществляется в коре головного мозга.
Под свойствами нервных процессов понимают такие характеристики возбуждения и торможения, как сила, уравновешенность и подвижность этих процессов.
Сила нервных процессов. При измерении силы процесса возбуждения обычно пользуются кривой зависимости величины условной реакции от силы раздражителя. Условная реакция перестает увеличиваться при определенной интенсивности условного сигнала. Эта граница и характеризует силу процесса возбуждения. Показателем силы тормозного процесса является стойкость тормозных условных рефлексов, а также скорость и прочность выработки дифференцировочного и запаздывающего вида торможения.
Уравновешенность нервных процессов. Для определения уравновешенности нервных процессов сравниваются силы процессов возбуждения и торможения у данного животного. Если оба процесса взаимно компенсируют друг друга, то они уравновешены, а если нет, то, например, при выработке дифференцировок может наблюдаться срыв тормозного процесса, если он оказывается слабым. Если же доминирует тормозной процесс в силу недостаточности возбуждения, то в трудных условиях дифференци-ровка сохраняется, но резко уменьшается величина реакции на положительный условный сигнал.
Подвижность нервных процессов. О ней можно судить по скорости переделки положительных условных рефлексов в тормозные и обратно. Часто для определения подвижности нервных процессов применяется переделка динамического стереотипа. Если переход от положительной реакции к тормозной и от тормозной к положительной осуществляется быстро, то это свидетельствует о высокой подвижности нервных процессов.
Регуляция нервной деятельности представляет собой процессы возбуждения и торможения в ЦНС. Вначале она возникает как элементарная реакция на раздражение. В процессе эволюции произошло усложнение нейрогуморальных функций, приводящее к образованию основных отделов нервной и эндокринной систем. В данной статье мы изучим один из главных процессов – торможение в ЦНС, виды и механизмы его осуществления.
Нервная ткань, её строение и функции
Одна из разновидностей животных тканей, названная нервной, имеет особое строение, обеспечивающее как процесс возбуждения, так и приводящее в действие функции торможения в ЦНС. Нервные клетки состоят из тела и отростков: коротких (дендритов) и длинного (аксона), который обеспечивает передачу нервных импульсов от одного нейроцита к другому. Окончание аксона нервной клетки контактирует с дендритами следующего нейроцита в местах, называемых синапсами. Они обеспечивают передачу биоэлектрических импульсов по нервной ткани. Причем возбуждение всегда движется в одном направлении – с аксона на тело или дендриты другого нейроцита.
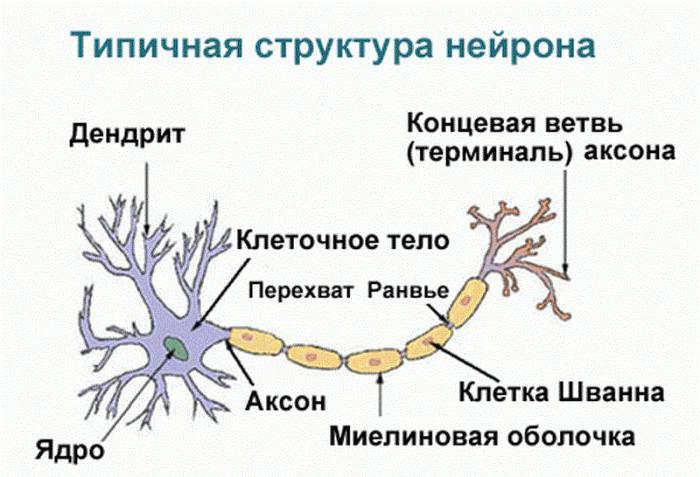
Еще одно свойство, кроме возбуждения, протекающее в нервной ткани, – торможение в ЦНС. Оно является ответной реакцией организма на действие раздражителя, ведущей к снижению или полному прекращению двигательной или секреторной активности, в которой участвуют центробежные нейроны. Торможение в нервной ткани может возникать и без предварительного возбуждения, а только лишь под воздействием тормозного медиатора, например ГАМК. Он является одним из главных трансмиттеров торможения. Здесь же можно назвать такое вещество, как глицин. Эта аминокислота участвует в усилении тормозных процессов и стимулирует в синапсах выработку молекул гаммааминомаслянной кислоты.
И. М. Сеченов и его работы в нейрофизиологии
Выдающийся российский ученый, создатель теории рефлекторной деятельности головного мозга доказал наличие в центральных отделах нервной системы особых комплексов клеток, способных к инактивации биоэлектрических процессов. Открытие центров торможения в ЦНС стало возможным благодаря применению И. Сеченовым трех видов экспериментов. К ним относятся: перерезание участков коры в различных зонах головного мозга, стимуляция отдельных локусов серого вещества физическими или химическими факторами (электрическим током, раствором хлорида натрия), а также метод физиологического возбуждения мозговых центров. И. М. Сеченов был прекрасным экспериментатором, проводя сверхточные разрезы в зоне между зрительными буграми и непосредственно в самом таламусе лягушки. Он наблюдал уменьшение и полное прекращение двигательной активности конечностей животного.
Взаимодействуют ли нервные процессы?
Исследования выдающихся российских физиологов И. П. Павлова и И. М. Сеченова доказали, что работа центральной нервной системы характеризуется координацией рефлекторных реакций организма. Взаимодействие процессов возбуждения и торможения в ЦНС приводит к согласованной регуляции функций организма: двигательной активности, дыхания, пищеварения, выделения. Биоэлектрические процессы одновременно происходят в нервных центрах и могут последовательно меняться во времени. Это обеспечивает корреляцию и своевременное прохождение ответных рефлексов на сигналы внутренней и внешней среды. Многочисленные опыты, проведенные нейрофизиологами, подтвердили тот факт, что возбуждение и торможение в ЦНС – это ключевые нервные явления, в основе которых лежат некоторые закономерности. Остановимся на них подробнее.
Нервные центры коры головного мозга способны распространять оба вида процессов по всей нервной системе. Это свойство называется иррадиацией возбуждения или торможения. Противоположное явление – уменьшение или ограничение участка мозга, распространяющего биоимпульсы. Оно названо концентрацией. Оба вида взаимодействий ученые наблюдают в течение образования условных двигательных рефлексов. Во время начальной стадии формирования двигательных навыков, вследствие иррадиации возбуждения одновременно сокращаются сразу несколько групп мышц, не обязательно участвующих в выполнении формируемого двигательного акта. Только после многократных повторений формируемого комплекса физических движений (катания на коньках, лыжах, велосипеде), в результате концентрации процессов возбуждения в конкретных нервных очагах коры, все движения человека становятся высококоординированными.

Переключения в работе нервных центров могут происходить также вследствие индукции. Она проявляется при выполнении следующего условия: сначала происходит концентрация торможения или возбуждения, причем эти процессы должны быть достаточной силы. В науке известны два вида индукции: S-фаза (центральное торможение в ЦНС усиливает возбуждение) и отрицательная форма (возбуждение вызывает процесс торможения). Встречается также последовательная индукция. В этом случае нервный процесс меняется на противоположный в самом нервном центре. Исследования нейрофизиологов доказали тот факт, что поведение высших млекопитающих и человека определяется явлениями индукции, иррадиации и концентрации нервных процессов возбуждения и торможения.
Безусловное торможение
Рассмотрим более подробно виды торможения в ЦНС и остановимся на такой его форме, которая присуща как животным, так и человеку. Сам термин был предложен И. Павловым. Ученый считал этот процесс одним из врождённых свойств нервной системы и выделил два его вида: гаснущее и постоянное. Остановимся на них детальнее.
Допустим, в коре существует очаг возбуждения, генерирующий импульсы к рабочему органу (к мышцам, секреторным клеткам желез). Вследствие изменения условий внешней или внутренней среды возникает еще один возбужденный участок коры головного мозга. Он вырабатывает биоэлектрические сигналы большей интенсивности, что тормозит возбуждение в ранее активном нервном центре и его рефлекторной дуге. Гаснущее торможение в ЦНС приводит к тому, что интенсивность ориентировочного рефлекса постепенно уменьшается. Объяснение этому следующее: первичный раздражитель уже не вызывает процесса возбуждения в рецепторах афферентного нейрона.
Другой вид торможения, наблюдаемого как у человека, так и у животных, демонстрирует опыт, проведенный лауреатом нобелевской премии в 1904 году И. П. Павловым. Во время кормления собаки (с выведенной из щеки фистулой) экспериментаторы включали резкий звуковой сигнал – выделение слюны из фистулы прекращалось. Такой вид торможения ученый назвал запредельным.
Являясь врождённым свойством, торможение в ЦНС протекает по безусловно-рефлекторному механизму. Оно достаточно пассивно и не вызывает расхода большого количества энергии, приводя к прекращению условных рефлексов. Постоянное безусловное торможение сопровождает многие психосоматические заболевания: дискинезии, спастический и вялый параличи.
Что такое гаснущий тормоз
Продолжая изучать механизмы торможения в ЦНС, рассмотрим, что представляет собой один из его видов, названный гаснущим тормозом. Хорошо известно, что ориентировочный рефлекс представляет собой реакцию организма на воздействие нового постороннего сигнала. В этом случае в коре мозга образуется нервный центр, находящийся в состоянии возбуждения. Он и формирует рефлекторную дугу, отвечающую за реакцию организма и называемую ориентировочным рефлексом. Этот рефлекторный акт вызывает торможение условного рефлекса, происходящего в данный момент. После многоразового повторения постороннего раздражителя рефлекс, называемый ориентировочным, постепенно снижается и наконец исчезает. А значит, не вызывает больше торможения условного рефлекса. Такой сигнал и получил название гаснущего тормоза.
Таким образом, внешнее торможение условных рефлексов связано с влиянием на организм постороннего сигнала и является врождённым свойством центральной и периферической нервной системы. Внезапный или новый раздражитель, например, болевое ощущение, посторонний звук, изменение освещенности, не только вызывает ориентировочный рефлекс, но также способствует ослаблению или даже полному прекращению условно-рефлекторной дуги, активной в данный момент. Если посторонний сигнал (кроме болевого) действует повторно, торможение условного рефлекса проявляется меньше. Биологическая роль безусловной формы нервного процесса заключается в проведении ответной реакции организма на раздражитель, наиболее важный в данный момент.
Внутреннее торможение
Его другое название, используемое в физиологии высшей нервной деятельности, – условное торможение. Главная предпосылка возникновения такого процесса – отсутствие подкрепления сигналов, поступающих из внешнего мира, врождёнными рефлексами: пищеварительным, слюноотделительным. Возникшие в этих условиях процессы торможения в ЦНС требуют определенного временного интервала. Рассмотрим их виды более подробно.
Например, дифференцировочное торможение возникает как ответ на сигналы окружающей среды, совпадающие по амплитуде, интенсивности и силе к условному раздражителю. Эта форма взаимодействия нервной системы и окружающего мира позволяет организму более тонко различать раздражители и вычленять из их совокупности тот, который получает подкрепление врожденным рефлексом. Например, на звук звонка с силой 15 Гц, подкрепленный кормушкой с пищей, у собаки выработали условную слюноотделительную реакцию. Если к животному применить еще один звуковой сигнал, силой 25 Гц, не подкрепляя его пищей, в первой серии опытов у собаки из фистулы слюна будет выделяться на оба условных раздражителя. Через некоторое время у животного произойдет дифференциация этих сигналов, и на звук, силой 25 Гц слюна из фистулы перестанет выделяться, то есть разовьется дифференцировочное торможение.
Освободить мозг от информации, потерявшей жизненно значимую роль для организма, – эту функцию как раз и выполняет торможение в ЦНС. Физиология опытным путем доказала, что условные двигательные реакции, хорошо закрепленные выработанными навыками, могут сохраняться на протяжении всей жизни человека, например, катание на коньках, езда на велосипеде.
Подводя итог, можно сказать, что процессы торможения в ЦНС – это ослабление или прекращение определенных реакций организма. Они имеют очень большое значение, так как все рефлексы организма корригируются в соответствии с измененными условиями, а если условный сигнал потерял свое значение, то даже полностью могут исчезать. Различные виды торможения в ЦНС являются базовыми для таких способностей психики человека, как сохранение самообладания, различение раздражителей, ожидание.
Запаздывающий вид нервного процесса
Опытным путем можно создать ситуацию, при которой ответ организма на условный сигнал из внешней среды проявляется еще до воздействия безусловного раздражителя, например пищи. При увеличении промежутка времени между началом воздействия условного сигнала (свет, звук, например, удары метронома) и моментом подкрепления до трех минут выделение слюны на вышеназванные условные раздражители все более запаздывает и проявляется только в момент, когда перед животным появляется кормушка с пищей. Отставание ответа на условный сигнал характеризует процессы торможения в ЦНС, названные запаздывающим видом, при котором его время протекания соответствует интервалу запаздывания безусловного раздражителя, например пищи.
Значение торможения в ЦНС
Биологические основы поведения человека
Оба вида активности нервной ткани, как возбуждение, так и торможение в ЦНС, являются основой высшей нервной деятельности. Она обуславливает физиологические механизмы психической деятельности человека. Учение высшей нервной деятельности было сформулировано И. П. Павловым. Современная его трактовка звучит следующим образом:
- Возбуждение и торможение в ЦНС, происходящие во взаимодействии, обеспечивают сложные психические процессы: память, мышление, речь, сознание, а также формируют сложные поведенческие реакции человека.
Чтобы составить научно обоснованный режим учебы, труда, отдыха, ученые применяют знания закономерностей высшей нервной деятельности.
Биологическое значение такого активного нервного процесса, как торможение, можно определить следующим образом. Изменение условий внешней и внутренней среды (отсутствие подкрепления условного сигнала врождённым рефлексом) влечет за собой адекватные изменения приспособительных механизмов в организме человека. Поэтому приобретенный рефлекторный акт угнетается (гаснет) или вовсе исчезает, так как становится для организма нецелесообразным.
Что такое сон?
И. П. Павлов в своих работах экспериментально доказал тот факт, что процессы торможения в ЦНС и сон имеют единую природу. В период бодрствования организма на фоне общей активности коры головного мозга все же диагностируются отдельные её участки, охваченные внутренним торможением. Во время сна оно иррадиирует по всей поверхности больших полушарий, достигая подкорковых образований: зрительных бугров (таламуса), гипоталамуса, ретикулярной формации и лимбической системы. Как указывал выдающийся нейрофизиолог П. К. Анохин, все вышеперечисленные части центральной нервной системы, ответственные за поведенческую сферу, эмоции и инстинкты, во время сна снижают свою активность. Это влечет за собой снижение генерирования нервных импульсов, поступающих из-под корки. Таким образом, активизация коры снижается. Это обеспечивает возможность покоя и восстановления обмена веществ как в нейроцитах большого мозга, так и во всем организме в целом.

Опытами других ученых (Гесса, Экономо) были установлены особые комплексы нервных клеток, входящие в неспецифические ядра зрительных бугров. Процессы возбуждения, диагностируемые в них, вызывают снижение частоты биоритмов коры, которые можно расценивать как переход от активного состояния (бодрствования) ко сну. Исследования таких участков головного мозга, как Сильвиев водопровод и ІІІ желудочек, подтолкнули ученых к идее о наличии центра регуляции сна. Он анатомически связан с участком мозга, ответственным за бодрствование. Поражение этого локуса коры вследствие травмы или в результате наследственных нарушений у человека приводит к патологическим состояниям бессонницы. Также отметим тот факт, что регуляция такого жизненно важного для организма процесса торможения, как сон, осуществляется нервными центрами промежуточного мозга и подкорковых ядер: хвостатого, миндалевидного, ограды и чечевицеобразного.
Читайте также:
