
Интрамуральный ганглий что это такое
Ганглии (иначе - нервные узлы) – это совокупность особых клеток. Она состоит из тел, дендритов и аксонов. Они, в свою очередь, относятся к нервным клеткам. Также нервные узлы включают в себя вспомогательные глиальные клетки. Их задачей является создание опоры для нейронов. Как правило, нервные ганглии покрыты соединительными тканями. Эти скопления встречаются не только у позвоночных, но и у некоторых беспозвоночных животных. Соединяясь между собой, нервные узлы создают сложные структурные системы. Примером могут являться структуры цепочки или сплетения. Далее в статье подробнее будет описано, что такое нервные узлы, как происходит взаимодействие между ними. Кроме того, будет приведена классификация и описание основных видов.
Позвоночные животные
Ганглии, существующие у этих особей, имеют некоторые особенности. Так, они не входят в пределы центральной нервной системы. Некоторые называют их базальными ганглиями. Однако наиболее правильным считается термин "ядро". Нервные узлы и система, которую они формируют, являются связующими элементами между компонентами нервной системы. Они пропускают импульсы и осуществляют управление работой определенных внутренних органов.
Классификация
Все ганглии подразделяются на несколько типов. Рассмотрим основные. Понятие "спинальный ганглий" объединяет сенсорные (афферентные) элементы. Второй тип - автономные элементы. Они располагаются в соответствующей (автономной) нервной системе. Основной вид - базальный. Их составляющими являются нейронные узлы, которые находятся в белом веществе. Оно содержится в головном мозге. Работа нейронов заключается в регулировании некоторых функций организма, а также в содействии в выполнении нервных процессов. Существует также вегетативный тип. Он представляет собой один узел нервов. Относится этот элемент к вегетативной нервной системе. Эти узлы проходят вдоль позвоночника. Вегетативные ганглии являются очень маленькими. Их размер может быть меньше миллиметра, а самые большие соизмеримы с горошинами. Задачей вегетативных ганглиев является регуляция функционирования внутренних органов и распределение импульсов.
Сравнение с термином "сплетение"
В книгах часто встречается понятие "сплетение". Его можно принять за синоним к слову "ганглии". Однако сплетением называют конкретные нервные узлы. Они присутствуют в определенном количестве на замкнутом участке. А ганглий – это область соединения синаптических контактов.
Нервная система
С точки зрения анатомии выделяются два ее типа. Первый называют центральной нервной системой. Сюда можно отнести головной мозг и спинной. Второй тип представляет собой совокупность узлов, нервных окончаний и самих нервов. Этот комплекс носит название периферической нервной системы.
Нервную систему формируют нервная трубка и ганглиозная пластина. К черепной части первой относятся головной мозг с органами чувств, к туловищному отделу – спинной мозг. Ганглиозная пластина формирует спинномозговые, вегетативные узлы и хромаффинную ткань. Нервная ткань существует как слагаемое системы, регулирующее соответствующие процессы организма.

Общие сведения
Нервные узлы – это объединение нервных клеток, выходящее за границы центральной нервной системы. Существуют вегетативные и чувствительные виды. Последние располагаются рядом с корешками спинного мозга и черепно-мозговыми нервами. По форме спинномозговой узел напоминает веретено. Окружен он оболочкой из соединительной ткани. Она также проникает в сам узел, при этом удерживая в себе кровеносные сосуды. Нервные клетки, находящиеся в спинномозговом узле, светлые, крупного размера, ядра их легко различимы. Нейроны формируют группы. Составляющими центра спинномозгового узла являются отростки нервных клеток и прослойки эндоневрия. Отростки-дендриты начинаются в чувствительной зоне спинномозговых нервов, а заканчиваются в периферийной части, где находятся их рецепторы. Нередким случаем является превращение биполярных нейронов в псевдоуниполярные. Это происходит во время их созревания. Из псевдоуниполярного нейрона выходит отросток, обвивающий клетку. Он разграничивается на афферентную, другое название "дендритная", и эфферентную, иначе - аксональную, части.
Дендриты и аксоны
Эти структуры покрывают миелиновые оболочки, составляющими которых являются нейролеммоциты. Нервные клетки спинномозгового узла окружают клетки олигодендроглий, имеющие такие названия, как мантийные глиоциты, глиоциты натрия, а также клетки-сателлиты. У этих элементов очень маленькие круглые ядра. Кроме того, оболочку этих клеток окружает капсула из соединительных тканей. Компоненты ее отличаются от прочих ядрами овальной формы. Биологически активными веществами, содержащимися в нервных клетках спинномозгового узла, являются ацетилхолин, глутаминовая кислота, субстанция P.
Вегетативные, или автономные, структуры
Автономные нервные узлы располагаются в нескольких местах. Во-первых, близ позвоночника (там находятся паравертебральные структуры). Во-вторых, перед позвоночником (превертебральные). Помимо этого, автономные узлы иногда находятся и в стенках органов. Например, в сердце, бронхах и мочевом пузыре. Такие ганглии называют интрамуральными. Еще один вид находится неподалеку от поверхности органов. С автономными структурами соединяются преганглионарные нервные волокна. Они обладают отростками нейронов из ЦНС. Вегетативные скопления делятся на два типа: симпатические и парасимпатические. Почти для всех органов получение постганглионарных волокон осуществляется от клеток, которые могут находиться в обоих типах вегетативных структур. Но воздействие, которое оказывают нейроны, отличается в зависимости от типа скоплений. Так, симпатическое действие может усиливать работу сердца, тогда как парасимпатическое ее замедляет.
Строение
Независимо от типа автономного узла их строение практически полностью совпадает. Каждую структуру покрывает оболочка соединительной ткани. В вегетативных узлах существуют особенные нейроны под названием "мультиполярные". Их отличает необычная форма, а также местоположение ядра. Существуют нейроны с несколькими ядрами и клетки с увеличенным количеством хромосом. Нейронные элементы и их отростки заключены в капсулу, составляющими которой являются глиальные клетки-сателлиты. Их называют мантийными глиоцитами. На верхнем слое этой оболочки находится мембрана, окруженная соединительной тканью.

Интрамуральные структуры
Эти нейроны вместе с проводящими путями могут составлять собой метасимпатический участок автономной нервной системы. По мнению гистолога Догеля, среди интрамуральных типов структур выделяются клетки трех разновидностей. К первым относятся длинноаксонные эфферентные элементы I типа. Эти клетки имеют нейроны больших размеров, у которых дендриты длинные, а аксон короткий. Для равноотросчатых афферентных нервных компонентов характерны длинные и дендриты, и аксон. А ассоциативные нейроны соединяют между собой клетки двух первых типов.
Периферическая система
Задача нервов обеспечивать связью нервные центры спинного, головного мозга и нервные структуры. Элементы системы взаимодействуют посредством соединительной ткани. Нервные центры – это области, отвечающие за обработку информации. Почти все рассматриваемые структуры состоят и из афферентных волокон, и из эфферентных. Набор волокон, являющийся, собственно, нервом, может содержать в себе не только структуры, защищенные электроизолирующей миелиновой оболочкой. В них присутствуют и те, что такого "покрытия" не имеют. Кроме того, нервные волокна разделены прослойкой соединительной ткани. Ее отличает рыхлость и волокнистость. Называется эта прослойка эндоневрием. Он содержит малое количество клеток, основную его часть составляют коллагеновые ретикулярные волокна. В этой ткани находятся небольшие кровеносные сосудики. Некоторые пучки с нервными волокнами окружает слой другой соединительной ткани – периневрий. Его составляющими являются последовательно расположенные клетки и волокна коллагена. Капсула, обволакивающая весь нервный ствол (она называется эпиневрий), образуется из соединительной ткани. Она, в свою очередь, обогащена клетками-фибробластами, макрофагами и компонентами жира. В ней находятся кровеносные сосуды с нервными окончаниями.
Интрамуральные нервные узлы внутренних органов и связанные с ними проводящие пути ввиду их высокой автономии, сложности организации и особенностей медиаторного обмена иногда выделяются в самостоятельный метасимпатический отдел вегетативной нервной системы.
В интрамуральных узлах русским гистологом Догелем А.С. описаны нейроны трех типов:
длинноаксонные эфферентные клетки I типа;
равноотросчатые афферентные клетки II типа;
ассоциативные клетки III типа.
Длинноаксонные эфферентные нейроны (клетки Догеля I типа) - многочисленные и крупные нейроны с короткими дендритами и длинным аксоном, который направляется за пределы узла к рабочему органу, где образует двигательные или секреторные окончания.
Равноотросчатые афферентные нейроны (клетки Догеля II типа) имеют длинные дендриты и аксон, уходящий за пределы данного узла в соседние. Эти клетки входят в качестве рецепторного звена в состав местных рефлекторных дуг, которые замыкаются без захода нервного импульса в ЦНС.
Ассоциативные нейроны (клетки Догеля III типа) - это местные вставочные нейроны, соединяющие своими отростками несколько клеток I и II типа.
Висцеровисцеральный рефлекс включает пути, в которых возбуждение возникает и заканчивается во внутренних органах. В этом случае эффектор способен отвечать либо усилением, либо торможением функций. Основой для осуществления этих процессов являются местные рефлекторные дуги, замыкающиеся в узлах автономной нервной системы. Эти дуги могут быть разного уровня: одни из них замыкаются в интрамуральных ганглиях, т.е. обеспечиваются метасимпатической нервной системой, другие — в предпозвоночных ганглиях, третьи — в спинальных ганглиях и структурах более высокого уровня.
3.Аномалии развития зародыша. Их причины. Понятие о критических периодах эмбриогенеза.
Отдельные ткани и органы формируются в различные периоды роста эмбриона и плода. При этом ткани организма в момент максимальной интенсивности процессов дифференцировки становятся высоко чувствительными к повреждающим воздействием внешней среды (ионизирующая радиация, инфекции, химические агенты и пр.).
Такие периоды, для которых характерна повышенная чувствительность к воздействию повреждающих факторов, называют критическими периодами эмбриогенеза. Вероятность формирования отклонений в развитии в критические периоды наиболее высока.
В онтогенезе человека можно выделить несколько критических периодов развития: в прогенезе, эмбриогенезе и постнатальной жизни. К ним относятся:
1) развитие половых клеток – овогенез и сперматогенез;
3) имплантация (7 – 8-е сутки эмбриогенеза);
4) развитие осевых зачатков органов и формирование плаценты (3–8-я неделя развития);
5) стадия усиленного роста головного мозга (15–20-я неделя);
6) формирование основных функциональных систем организма и дифференцировка полового аппарата (20–24-я неделя);
8) период новорожденности (до 1 года);
9) половое созревание (11– 16 лет)
Влияние тератогенных факторов чаще всего реализуется в виде развития множественных пороков и аномалий развития, формирование которых зависит от дозы повреждающего агента, продолжительности его воздействия и срока беременности, на котором произошло неблагоприятное воздействие.
14.1. Компоненты нервной системы
14.1.1. Подразделение нервной системы
на центральную и периферическую
2. Последняя включает:
1. а) Нервные узлы (или ганглии) - это скопления нервных клеток (точнее, их тел) вне центральной нервной системы.
б) Скопления же нейронов в головном или спинном мозгу называются ядрами .
2. а) А нервные стволы - это совокупность идущих параллельно нервных волокон.
б) Среди последних в нерве могут одновременно присутствовать
а) два типа образований периферической нервной системы -
б) а также один из двух компонентов центральной нервной системы -
б) Они различаются в строении и локализации нервных ядер, узлов и стволов.
14.1.2. Соматическая нервная система
и её рефлекторная дуга
14.1.2.1. Введение
2. Её рефлекторная дуга обычно включает три части :

14.1.2.2. Чувствительные нейроны
б) Узлы же располагаются по ходу
в) А. В первом случае узел называется спинномозговым (1) .
а) В одних случаях они (как показано на схеме)
б) В других случаях аксоны чувствительных нейронов
14.1.2.3. Ассоциативные нейроны
б) Тела последующих ассоциативных нейронов (если они вовлечены в дугу) могут находиться также в других отделах ЦНС: в т.ч. в
б) Аксоны же могут
14.1.2.4. Двигательные нейроны
Б. Импульсы от ассоциативных нейронов обычно поступают по многочисленным аксо соматическим синапсам (п. 13.3.4).
б) Тела (8.А) двигательных нейронов находятся
б) А б лагодаря восходящим и нисходящим связям спинного мозга с корой больших полушарий (создающим более сложные дуги), человек может
14.1.3. Вегетативная нервная система
14.1.3.1. Функция
I. Общие сведения
а) В свою очередь, вегетативная нервная система подразделяется на два отдела, или две системы:
II. Действия симпатической и парасимпатической систем
процессы восстановления:
спазма сосудов и
усиления сердцебиения,
14.1.3.2. Общие особенности вегетативной нервной системы
Вегетативная нервная система имеет ряд особенностей.-
б) По отношению к этим ганглиям, различают 2 вида вегетативных нервных волокон:
б) Медиаторы же в окончаниях пост ганглионарных волокон различны:
14.1.3.3. Рефлекторные дуги
симпатической нервной системы
В случае симпатической нервной системы вышеперечисленные особенности проявляются следующим образом.
а) Локализация остальных частей данных нейронов обычна:

б) Постганглионарные волокна (10) , отходящие от экстрамуральных узлов, во-первых,
в) Кроме того, от узлов симпатического ствола часть постганглионарных волокон (в составе т.н. серых соединительных ветвей)
14.1.3.4. Рефлекторные дуги
парасимпатической нервной системы
б) Аксоны этих нейронов образуют преганглионарные парасимпатические волокна и идут в составе
б) Поэтому постганглионарные волокна парасимпатической системы (образованные аксонами эффекторных нейронов) -
14.1.3.5. Периферические вегетативные рефлекторные дуги
но также других нейронов:
б) Тогда последовательность прохождения сигнала и ответа такова.-
Теперь рассмотрим подробней морфологию упоминавшихся выше компонентов нервной системы -
14.2. Нервные стволы и нервные узлы
14.2.1. Нервные стволы
14.2.1.1. Соединительнотканные элементы
а) Малое увеличение

прослойки соединительной ткани между пучками называются периневрием (5 ).
б) Большое увеличение

14.2.1.2. Нервные волокна
б ) В данном препарате нерв состоит, главным образом, из миелиновых нервных волокон (п. 12.4.3.1) :
б) Большое увеличение
14.2.2. Нервные узлы
Здесь будет рассмотрено строение трёх типов узлов -
14.2.2.1. Чувствительные узлы
а) В качестве пример а обратимся к спинномозговы м узл ам .
б) Строение чувствительных узлов, лежащих по ходу некоторых черепномозговых нервов, аналогично.
I. Корешки спинного мозга и местоположение спинномозговых узлов

II. Строение узла
б ) А. В центре узла ( между группами нейронов ) проходят нервные волокна ( 5 ).
б) В соответствии с п. 12.2.2.2 , эти нейроны имеют

14.2.2.2. Симпатические узлы
Согласно п. 14.1.3.4, такие узлы находятся
I. Общий вид

II. Эффекторные нейроны

б) Видимо, с одним нейроном контактируют сразу несколько преганглионарных волокон (чем и объясняется наличие нескольких дендритов).
2. Аксоны эффекторных нейронов , покидая узел и подходя к органу, образуют , как мы уже говорили (п. 14.1.3. 2) ,
III. МИФ-клетки
2. Их на данном препарате различить не удаётся.
3. а) Считают, что, возбуждаясь преганглионарными волокнами, МИФ-клетки частично
14.2.2.3. Интрамуральные ганглии
I. Общий вид
1. а) Интрамуральный ганглий (1) выявляется как скопление нервных клеток в толще органа.
б) В данном случае видна окружающая мышечная ткань (2) стенки мочевого пузыря.
а) 
б) Вокруг них, как обычно, находятся глиальные клетки-сателлиты (4) и соединительнотканные элементы.
б) (Другое поле зрения) .jpg)
II. Типы нейронов
14.3. Спинной мозг
2. Как видно, схема устанавливает связь между
14.3.2. План строения
14.3.2.1. Оболочки мозга
2. Первые две образованы рыхлой волокнистой соединительной тканью,
4. а) Паутинная оболочка не заходит в углубления мозга.
б) Поэтому между ней и мягкой оболочкой образуется
В самом спинном мозгу можно различить
14.3.2.2. Серое вещество: общие сведения

14.3.2.3. Белое вещество: общие сведения
б) Между передними канатиками - глубокая срединная вырезка (1 0 ), просвет которой на препарате не всегда виден.
Теперь дадим более детальную характеристику серого и белого вещества.
14.3.3. Серое вещество спинного мозга
14.3.3.1. Функциональные структуры серого вещества
А. ЗАДНИЕ РОГА
Как мы знаем, в задних рогах содержатся вставочные (ассоциативные) нейроны, которые получают сигналы от чувствительных нейронов спинномозговых узлов.
Нейроны задних рогов образуют следующие структуры.
Аксоны этих нейронов идут к мотонейронам передних рогов того же сегмента спинного мозга -
Б. ПРОМЕЖУТОЧНАЯ ЗОНА И БОКОВЫЕ РОГА
Здесь нейроны сгруппированы в два или одно ядро (в зависимости от уровня спинного мозга).
В. ПЕРЕДНИЕ РОГА
Г. НЕЙРОНЫ, СОДЕРЖАЩИЕСЯ ВО ВСЕХ РОГАХ
14.3.3.2. Классификация нейронов спинного мозга
I. Классификация всех нейронов по местонахождению их аксонов
II. Классификация клеток передних рогов
2. Поэтому данные клетки, иннервируя в скелетных мышцах экстрафузальные мышечные волокна, участвуют
2. Таким образом, они стимулируют цепочку событий. -
14.3.3.3. Просмотр препарата
Рассмотрим отдельные фрагменты предыдущего препарата.
Мы видим глиальную строму, в которой располагаются нейроны.

а) Нейроны (1), лежащие ближе к центральному каналу,

а) Видны крупные мотонейроны (1) с телами неправильной формы и отходящими от них отростками.

14.3.4. Белое вещество спинного мозга
1. а) Микроскопическое строение белого вещества спинного мозга однообразно:
на поперечном срезе это сечения миелиновых (как правило) нервных волокон.
б) В центре каждого волокна - осевой цилиндр, который окружён толстой миелиновой оболочкой.
2. Но в функциональном отношении проводящие пути белого вещества различны. –
1. Восходящие пучки
(Голля и Бурдаха) -
2. Восходящие пучки:
3. Нисходящие пучки:
а) от коры больших полушарий
(боковой пирамидный тракт );
а) Содержат аксоны нейронов головного мозга.
4. Нисходящие пучки:
а) от коры больших полушарий
( передний пирамидный тракт) ,
б) от среднего мозга
(от подкорковых центров зрения и слуха),
В вегетативной нервной системе различают центральные и периферические отделы. Центральные отделы симпатической нервной системы представлены ядрами боковых рогов тораколюмбального отдела спинного мозга. В парасимпатической нервной системе центральные отделы включают ядра среднего и продолговатого мозга, а также ядра боковых рогов сакрального отдела спинного мозга. Парасимпатические волокна краниобульбарного отдела выходят в составе III-й, VII-й, IX-й и Х-й пар черепных нервов.
Периферические отделы вегетативной нервной системы образованы нервными стволами, ганглиями и сплетениями.
Вегетативные рефлекторные дуги начинаются чувствительным нейроном, тело которого лежит в спинномозговом узле (ганглии), как и в соматической рефлекторной дуг. Ассоциативные нейроны находятся в боковых рогах спинного мозга. Здесь нервные импульсы переключаются на промежуточные преганглионарные нейроны, отростки которых покидают центральные ядра и достигают вегетативных ганглиев, где передают импульсы на двигательный нейрон. В связи с этим различают нервные волокна преганглионарные и постганглионарные. Первые из них покидают центральную нервную систему в составе вентральных корешков спинномозговых нервов и черепных нервов. Как в симпатической, так и в парасимпатической системах преганглионарные нервные волокна принадлежат холинергическим нейронам. Аксоны нейронов, расположенных в вегетативных ганглиях, называются постганглионарными. Они не образуют прямых контактов с эффекторными клетками. Их терминальные отделы по своему ходу формируют расширения — варикозности, в составе которых находяся пузырьки медиатора. В области варикозности нет глиальной оболочки и нейромедиатор, выделяясь в окружающую среду, влияет на эффекторные клетки (например, на клетки желез, гладкие миоциты и др.).
В периферических ганглиях симпатической нервной системы, как правило, находятся адренергические эфферентные нейроны (за исключением нейронов, имеющих синаптические связи с потовыми железами, где симпатические нейроны являются холинергическими). В парасимпатических ганглиях эфферентные нейроны всегда холинергические.
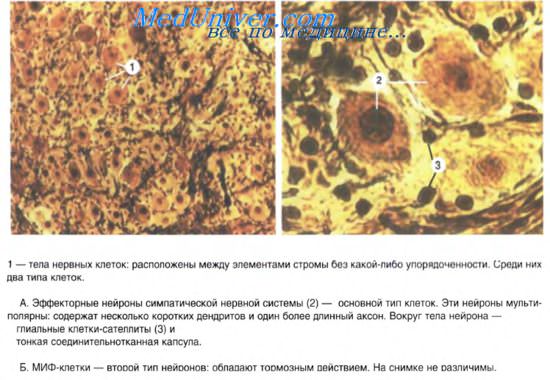
Ганглии представляют собой скопления мультиполярных нейронов (от нескольких клеток до десятков тысяч). Экстраорганные (симпатические) ганглии имеют хорошо выраженную соединительнотканную капсулу, как продолжение периневрия. Парасимпатические ганглии находятся, как правило, в интрамуральных нервных сплетениях. Ганглии интрамуральных сплетений, как и другие вегетативные узлы, содержат вегетативные нейроны местных рефлекторных дуг. Мультиполярные нейроны диаметром 20-35 мкм расположены диффузно, каждый нейрон окружен глиоцитами ганглия. Кроме того, описаны нейроэндокринные, хеморецепторные, биполярные, а у некоторых позвоночных и униполярные нейроны. В симпатических ганглиях имеются мелкие интенсивно флюоресцирующие клетки (МИФ-клетки) с короткими отростками и большим количеством гранулярных пузырьков в цитоплазме. Они выделяют катехоламины и оказывают тормозящее влияние на передачу импульсов с преганглионарных нервных волокон на эфферентный симпатический нейрон. Эти клетки называют интернейронами.
Среди крупных мультиполярных нейронов вегетативных ганглиев различают: двигательные (клетки Догеля I-го типа), чувствительные (клетки Догеля II-го типа) и ассоциативные (клетки Догеля III-го типа). Двигательные нейроны имеют короткие дендриты с пластинчатыми расширениями ("рецептивные площадки"). Аксон этих клеток очень длинный, уходит за пределы ганглия в составе постганглионарных тонких безмиелиновых нервных волокон и оканчивается на гладких миоцитах внутренних органов. Клетки I-го типа называют длинноаксонными нейронами. Нейроны II-го типа — равноотростчатые нервные клетки. От их тела отходят 2-4 отростка, среди которых различить аксон трудно. Не разветвляясь, отростки уходят далеко от тела нейрона. Их дендриты имеют чувствительные нервные окончания, а аксон оканчивается на телах двигательных нейронов в соседних ганглиях. Клетки II-го типа являются чувствительными нейронами местных вегетативных рефлекторных дуг. Клетки Догеля III-го типа по форме тела похожи на вегетативные нейроны П-го типа, но их дендриты не выходят за пределы ганглия, а нейрит направляется в другие ганглии. Многие исследователи считают эти клетки разновидностями чувствительных нейронов.
Таким образом, в периферических вегетативных ганглиях имеются местные рефлекторные дуги, состоящие из чувствительных, двигательных и, возможно, ассоциативных вегетативных нейронов.
Интрамуральные вегетативные ганглии в стенке пищеварительного тракта отличаются тем, что в их составе, кроме двигательных холинергических нейронов, имеются тормозные нейроны. Они представлены адренергическими и пуринергическими нервными клетками. В последних медиатором является пуриновый нуклеотид. В интрамуральных вегетативных ганглиях встречаются также пептидергические нейроны, выделяющие вазоинтестинальный пептид, соматостатин и ряд других пептидов, с помощью которых осуществляются нейроэндокринная регуляция и модуляция деятельности тканей и органов пищеварительной системы.
ГАНГЛИИ (ganglia нервные узлы)— скопления нервных клеток, окруженные соединительной тканью и клетками глии, расположенные по ходу периферических нервов.
Различают Г. вегетативной и соматической нервной системы. Г. вегетативной нервной системы делятся на симпатические и парасимпатические и содержат тела постганглионарных нейронов. Г. соматической нервной системы представлены спинномозговыми узлами и Г. чувствительных и смешаных черепных нервов, содержащих тела чувствительных нейронов и дающих начало чувствительным порциям спинномозговых и черепных нервов.
Содержание
- 1 Эмбриология
- 2 Анатомия
- 3 Гистология
- 4 Физиология
- 5 Патология
- 6 Краткая анатомическая характеристика нервных ганглиев (узлов)
Эмбриология
Зачатком спинномозговых и вегетативных узлов является ганглиозная пластинка. Она образуется у эмбриона в тех отделах нервной трубки, которые граничат с эктодермой. У зародыша человека на 14—16-й день развития ганглиозная пластинка располагается по дорсальной поверхности замкнувшейся нервной трубки. Затем она расщепляется по всей длине, обе ее половины перемещаются вентрально и в виде нервных валиков лежат между нервной трубкой и поверхностной эктодермой. В дальнейшем соответственно сегментам дорсальной стороны зародыша в нервных валиках возникают очаги пролиферации клеточных элементов; эти участки утолщаются, обособляются и превращаются в спинномозговые узлы. Из ганглиозной пластинки развиваются также чувствительные ганглии У, VII—X пар черепных нервов, аналогичные спинномозговым ганглиям. Зародышевые нервные клетки, нейробласты, формирующие спинномозговые ганглии, представляют собой биполярные клетки, т. е. имеют два отростка, отходящих от противоположных полюсов клетки. Биполярная форма чувствительных нейронов у взрослых млекопитающих и человека сохраняется только в чувствительных клетках преддверно-улит кового нерва, преддверного и спирального ганглиев. В остальных, как спинномозговых, так и черепных чувствительных узлах, отростки биполярных нервных клеток в процессе их роста и развития сближаются и сливаются в большинстве случаев в один общий отросток (processus communis). По этому признаку чувствительные нейроциты (нейроны) называют псевдоуниполярными (neurocytus pseudounipolaris), реже протонейронами, подчеркивая древность их происхождения. Спинномозговые узлы и узлы в. н. с. отличаются характером развития и строения нейронов. Развитие и морфология вегетативных ганглиев — см. Вегетативная нервная система.
Анатомия
Основные сведения об анатомии Г. приведены в таблице.
Гистология


Возраст и перенесенные заболевания сказываются на структуре спинномозговых ганглиев — в них появляется значительно большее, чем у здоровых, количество различных атипичных нейронов, в особенности с дополнительными отростками, снабженными колбовидными утолщениями, как, напр., при ревматическом пороке сердца (рис. 2), стенокардии и др. Клинические наблюдения, а также экспериментальные исследования на животных показали, что чувствительные нейроны спинномозговых узлов реагируют значительно быстрее интенсивным ростом дополнительных отростков на различные эндогенные и экзогенные вредности, нежели моторные соматические или вегетативные нейроны. Эта способность чувствительных нейронов иногда выражена значительно. В случаях хрон, раздражений вновь образованные отростки могут обвиваться (в виде намотки) вокруг тела собственного или соседнего нейрона, напоминая кокон. Чувствительные нейроны спинномозговых узлов, как и другие типы нервных клеток, имеют ядро, различные органеллы и включения в цитоплазме (см. Нервная клетка). Т. о., отличительным свойством чувствительных нейронов спинномозговых и узлов черепных нервов является их яркая морфол, реактивность, выражающаяся в изменчивости их структурных компонентов. Это обеспечено высоким уровнем синтеза белков и различных активных веществ и свидетельствует об их функциональной подвижности.
Физиология
У беспозвоночных Г. играют ту же роль, что и ц. н. с. у позвоночных, являясь высшими центрами координации соматических и вегетативных функций. В эволюционном ряду от червей до головоногих моллюсков и членистоногих Г., перерабатывающие всю информацию о состоянии окружающей и внутренней среды, достигают высокой степени организации. Это обстоятельство, а также простота анатомической препаровки, относительно большие размеры тел нервных клеток, возможность введения в сому нейронов под прямым визуальным контролем одновременно нескольких микроэлектродов сделали Г. беспозвоночных общераспространенным объектом нейрофизиол, экспериментов. На нейронах круглых червей, октапод, декапод, брюхоногих и головоногих моллюсков методами электрофореза, прямого измерения активности ионов и фиксации напряжения проводят исследования механизмов генерации потенциалов и процесса синаптической передачи возбуждения и торможения, зачастую невыполнимые на большинстве нейронов млекопитающих. Несмотря на эволюционные различия, основные электрофизиол, константы и нейрофизиол, механизмы работы нейронов во многом одинаковы у беспозвоночных и высших позвоночных животных. Поэтому исследования Г., беспозвоночных имеют общефизиол. значение.
У позвоночных соматосенсорные черепные и спинномозговые Г. функционально однотипны. Они содержат тела и проксимальные части отростков афферентных нейронов, передающих импульсацию от периферических рецепторов в ц. н. с. В сомато-сенсорных Г. нет синаптических переключений, эфферентных нейронов и волокон. Так, нейроны спинномозговых Г. у жабы характеризуются следующими основными электрофизиол, параметрами: специфическим сопротивлением — 2,25 кОм/см 2 для деполяризующего и 4,03 кОм/см 2 для гиперполяризующего тока и удельной емкостью 1,07 мкФ/см 2 . Полное входное сопротивление нейронов сомато-сенсорных Г. существенно ниже, чем соответствующий параметр аксонов, поэтому при высокочастотной афферентной импульсации (до 100 импульсов в 1 сек.) проведение возбуждения может блокироваться на уровне тела клетки. В этом случае потенциалы действия, хотя и не регистрируются от тела клетки, продолжают проводиться с периферического нерва в задний корешок и сохраняются даже после экстирпации тел нервных клеток при условии интактности Т-образных ветвлений аксонов. Следовательно, возбуждение сомы нейронов сомато-сенсорных Г. для передачи импульсов от периферических рецепторов к спинному мозгу не обязательно. Эта особенность впервые появляется в эволюционном ряду у бесхвостых амфибий.
Вегетативные Г. позвоночных в функциональном плане принято разделять на симпатические и парасимпатические. Во всех вегетативных Г. происходит синаптическое переключение с преганглионарных волокон на постганглионарные нейроны. В подавляющем большинстве случаев синаптическая передача осуществляется хим. путем с помощью ацетилхолина (см. Медиаторы). В парасимпатическом цилиарном Г. птиц обнаружена электрическая передача импульсов при помощи так наз. потенциалов соединения, или потенциалов связи. Электрическая передача возбуждения через один и тот же синапс возможна в двух направлениях; в процессе онтогенеза она формируется позже химической. Функциональное значение электрической передачи пока не ясно. В симпатических Г. амфибий выявлено небольшое количество синапсов с хим. передачей нехолинергической природы. В ответ на сильное одиночное раздражение преганглионарных волокон симпатического Г. в постганглионарном нерве прежде всего возникает ранняя отрицательная волна (O-волна), обусловленная возбуждающими постсинаптическими потенциалами (ВПСП) при активации н-холинорецепторов постганглионарных нейронов. Тормозной постсинаптический потенциал (ТПСП), возникающий в постганглионарных нейронах под действием катехоламинов, выделяемых хромаффинными клетками в ответ на активацию их м-холинорецепторов, формирует следующую за 0-волной положительную волну (П-волна). Поздняя отрицательная волна (ПО-волна) отражает ВПСП постганглионарных нейронов при активации их м-холинорецепторов. Завершает процесс длительная поздняя отрицательная волна (ДПО-волна), возникающая вследствие суммации ВПСП нехолинергической природы в постганглионарных нейронах. В нормальных условиях на высоте О-волны при достижении ВПСП величины 8—25 мв возникает распространяющийся потенциал возбуждения с амплитудой 55—96 мв, длительностью 1,5—3,0 мсек, сопровождаемый волной следовой гиперполяризации. Последняя существенно маскирует волны П и ПО. На высоте следовой гиперполяризации возбудимость снижается (период рефрактерности), поэтому обычно частота разрядов постганглионарных нейронов не превышает 20—30 импульсов в 1 сек. По основным электрофизиол. характеристикам нейроны вегетативных Г. идентичны большинству нейронов ц. н. с. Нейрофизиол. особенностью нейронов вегетативных Г. является отсутствие истинной спонтанной активности при деафферентации. Среди пре- и постганглионарных нейронов преобладают нейроны групп В и С по классификации Гассера — Эрлангера, основанной на электрофизиол, характеристиках нервных волокон (см.). Преганглионарные волокна обширно ветвятся, поэтому раздражение одной преганглионарной ветви приводит к возникновению ВПСП во многих нейронах нескольких Г. (феномен мультипликации). В свою очередь на каждом постганглионарном нейроне оканчиваются терминали многих преганглионарных нейронов, отличающихся по порогу раздражения и скорости проведения (феномен конвергенции). Условно мерой конвергенции можно считать отношение количества постганглионарных нейронов к количеству преганглионарных нервных волокон. Во всех вегетативных Г. оно больше единицы (за исключением цилиарного ганглия птиц). В эволюционном ряду это отношение возрастает, достигая в симпатических Г. человека величины 100:1. Мультипликация и конвергенция, обеспечивающие пространственную суммации) нервных импульсов, в сочетании с временной суммацией являются основой интегрирующей функции Г. при обработке центробежной и периферической импульсации. Через все вегетативные Г. проходят афферентные пути, тела нейронов которых лежат в спинномозговых Г. Для нижнего брыжеечного Г., чревного сплетения и некоторых интрамуральных парасимпатических Г. доказано существование истинных периферических рефлексов. Афферентные волокна, проводящие возбуждение с малой скоростью (ок. 0,3 м/сек), входят в Г. в составе постганглионарных нервов и оканчиваются на постганглионарных нейронах. В вегетативных Г. обнаружены окончания афферентных волокон. Последние информируют ц. н. с. о происходящих в Г. функционально-хим. изменениях.
Патология
В клин, практике наиболее часто встречается ганглионит (см.), называемый также симпато-ганглионитом, — заболевание, связанное с поражением ганглиев симпатического ствола. Поражение нескольких узлов определяют как полиганглионит, или трунцит (см.).
Спинальные ганглии нередко вовлекаются в патологический процесс при радикулитах (см.).
Читайте также:
