
Нервно мышечное соединение его функция
Синапс представляет собой место функционального, а не физического контакта между нейронами; в нем происходит передача информации от одной клетки к другой. Обычно встречаются синапсы между концевыми веточками аксона одного нейрона и дендритами (аксодендритные синапсы) или телом (аксосоматические синапсы) другого нейрона. Число синапсов, как правило, очень велико, что обеспечивает большую площадь для передачи информации. Например, на дендритах и телах отдельных мотонейронов спинного мозга находится свыше 1000 синапсов. Некоторые клетки головного мозга могут иметь до 10000 синапсов (рис. 16.8).
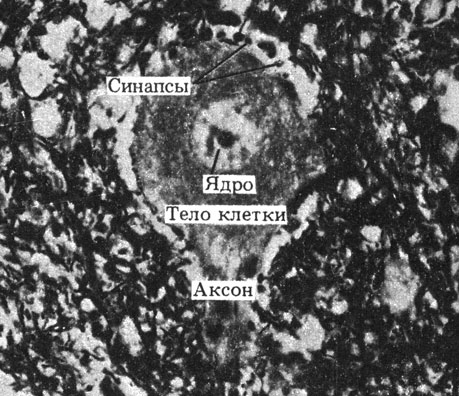
Рис. 16.8. Синапсы на двигательном нейроне (электронная микрофотография)
Существуют два типа синапсов - электрические и химические - в зависимости от природы проходящих через них сигналов. Между окончаниями двигательного нейрона и поверхностью мышечного волокна существует нервно-мышечное соединение, отличающееся по строению от межнейронных синапсов, но сходное с ними в функциональном отношении. Структурные и физиологические различия между обычным синапсом и нервно-мышечным соединением будут описаны несколько позже.
Химические синапсы - наиболее распространенный тип синапса у позвоночных. Это луковицеобразные утолщения нервных окончаний, называемые синаптическими бляшками и расположенные в непосредственной близости от окончания дендрита. Цитоплазма синаптической бляшки содержит митохондрии, гладкий эндоплазматический ретикулум, микрофиламенты и многочисленные синаптические пузырьки. Каждый пузырек имеет в диаметре около 50 нм и содержит медиатор - вещество, с помощью которого нервный сигнал передается через синапс. Мембрана синаптической бляшки в области самого синапса утолщена в результате уплотнения цитоплазмы и образует пресинаптическую мембрану. Мембрана дендрита в области синапса также утолщена и образует постсинаптическую мембрану. Эти мембраны разделены промежутком - синаптической щелью шириной около 20 нм. Пресинаптическая мембрана устроена таким образом, что к ней могут прикрепляться синаптические пузырьки и выделяться в синаптическую щель медиаторы. Постсинаптическая мембрана содержит крупные белковые молекулы, действующие как рецепторы медиаторов, и многочисленные каналы и поры (обычно закрытые), через которые в постсинаптический нейрон могут поступать ионы (см. рис. 16.10, А).
Синаптические пузырьки содержат медиатор, который образуется либо в теле нейрона (и попадает в синаптическую бляшку, пройдя через весь аксон), либо непосредственно в синаптической бляшке. В обоих случаях для синтеза медиатора нужны ферменты, образующиеся в теле клетки на рибосомах. В синаптической бляшке молекулы медиатора "упаковываются" в пузырьки, в которых они хранятся до момента высвобождения. Основные медиаторы нервной системы позвоночных - ацетилхолин и норадреналин, но существуют и другие медиаторы, которые будут рассмотрены позже.
Ацетилхолин - аммонийное производное, формула которого приведена на рис. 16.9. Это первый из известных медиаторов; в 1920 г. Отто Леви выделил его из окончаний парасимпатических нейронов блуждающего нерва в сердце лягушки (разд. 16.2). Структура норадреналина подробно рассматривается в разд. 16.6.6. Нейроны, высвобождающие ацетилхолин, называются холинэргическими, а высвобождающие норадреналин - адренэргическими.
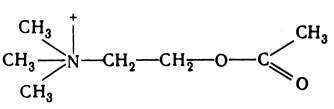
Рис. 16.9. Структурная формула ацетилхолина
Как полагают, прибытие нервного импульса в синаптическую бляшку вызывает деполяризацию пресинаптической мембраны и повышение ее проницаемости для ионов Са 2+ . Входящие в синаптическую бляшку ионы Са 2+ вызывают слияние синаптических пузырьков с пресинаптической мембраной и выход их содержимого из клетки (экзоцитоз), в результате чего оно попадает в синаптическую щель. Весь этот процесс называют электросекреторным сопряжением. После высвобождения медиатора материал пузырьков используется для образования новых пузырьков, заполняемых молекулами медиатора. Каждый пузырек содержит около 3000 молекул ацетилхолина.
Молекулы медиатора диффундируют через синаптическую щель (этот процесс занимает около 0,5 мс) и связываются с находящимися на постсинаптической мембране рецепторами, способными узнавать молекулярную структуру ацетилхолина. При связывании молекулы рецептора с медиатором ее конфигурация меняется, что приводит к открытию ионных каналов и поступлению в постсинаптическую клетку ионов, вызывающих деполяризацию или гиперполяризацию (рис. 16.4,А) ее мембраны в зависимости от природы высвобождаемого медиатора и строения молекулы рецептора. Молекулы медиатора, вызвавшие изменение проницаемости постсинаптической мембраны, сразу же удаляются из синаптической щели либо путем их реабсорбции пресинаптической мембраной, либо путем диффузии из щели или ферментативного гидролиза. В случае холинэргических синапсов находящийся в синаптической щели ацетилхолин гидролизуется ферментом ацетилхолинэстеразой, локализованным на постсинаптической мембране. В результате гидролиза образуется холин, он всасывается обратно в синаптическую бляшку и вновь превращается там в ацетилхолин, который хранится в пузырьках (рис. 16.10).
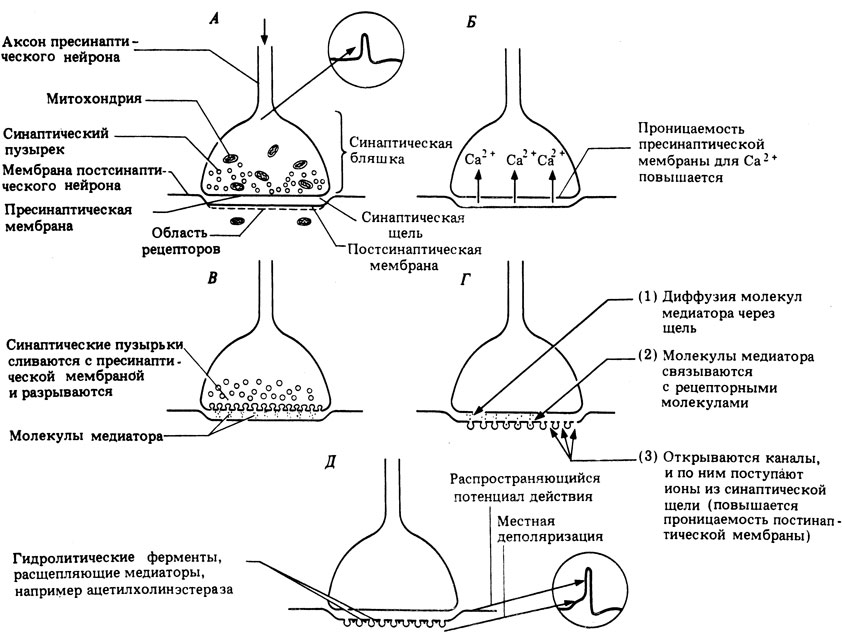
Рис. 16.10. Механизм химической передачи импульсов в нервном синапсе; от А до Д - последовательные этапы процесса
В возбуждающих синапсах под действием ацетилхолина открываются специфические натриевые и калиевые каналы, и ионы Na + входят в клетку, а ионы К + выходят из нее в соответствии с их концентрационными градиентами. В результате происходит деполяризация постсинаптической мембраны. Эту деполяризацию называют возбудительным постсинаптическим потенциалом (ВПСП). Амплитуда ВПСП обычно невелика, но продолжительность его больше, чем у потенциала действия. Амплитуда ВПСП меняется ступенчатым образом, и это позволяет предполагать, что медиатор освобождается порциями, или "квантами", а не в виде отдельных молекул. По-видимому, каждый квант соответствует освобождению медиатора из одного синаптического пузырька. Одиночный ВПСП не способен, как правило, вызвать деполяризацию пороговой величины, необходимой для возникновения потенциала действия. Но деполяризующие эффекты нескольких ВПСП складываются, и это явление носит название суммации. Два или больше ВПСП, возникших одновременно в разных синапсах одного и того же нейрона, могут сообща вызвать деполяризацию, достаточную для возбуждения потенциала действия в постсинаптическом нейроне. Это называют пространственной суммацией. Быстро повторяющееся высвобождение медиатора из пузырьков одной и той же синаптической бляшки под действием интенсивного стимула вызывает отдельные ВПСП, которые следуют так часто один за другим во времени, что их эффекты тоже суммируются и вызывают в постсинаптическом нейроне потенциал действия. Это называется временной суммацией. Таким образом, импульсы могут возникать в одиночном постсинаптическом нейроне либо как результат слабой стимуляции нескольких связанных с ним пресинаптических нейронов, либо как результат повторной стимуляции одного из его пресинаптических нейронов. В тормозных синапсах высвобождение медиатора повышает проницаемость постсинаптической мембраны за счет открытия специфических каналов для ионов К + и Сl - . Перемещаясь по концентрационным градиентам, эти ионы вызывают гиперполяризацию мембраны, называемую тормозным постсинаптическим потенциалом (ТПСП).
Медиаторы сами по себе не обладают возбуждающими или тормозящими свойствами. Например, ацетилхолин оказывает возбуждающее действие в большинстве нервно-мышечных соединений и других синапсов, но вызывает торможение в нервно-мышечных соединениях сердца и висцеральной мускулатуры. Эти противоположные эффекты обусловлены теми событиями, которые развертываются на постсинаптической мембране. От молекулярных свойств рецептора зависит, какие ионы будут входить в постсинаптический нейрон, а эти ионы в свою очередь определяют характер изменения постсинаптических потенциалов, как описано выше.
У многих животных, в том числе у кишечнополостных и позвоночных, передача импульсов через некоторые синапсы осуществляется путем прохождения электрического тока между пре- и постсинаптическими нейронами. Ширина щели между этими нейронами составляет всего лишь 2 нм, и суммарное сопротивление току со стороны мембран и жидкости, заполняющей щель, очень мало. Импульсы проходят через синапсы без задержки, и на их передачу не действуют лекарственные вещества или другие химические препараты.
Нервно-мышечное соединение представляет собой специализированный вид синапса между окончаниями двигательного нейрона (мотонейрона) и эндомизием мышечных волокон (разд. 17.4.2). Каждое мышечное волокно имеет специализированный участок - двигательную концевую пластинку, где аксон моторного нейрона (мотонейрона) разветвляется, образуя немиелинизированные веточки толщиной около 100 нм, проходящие в неглубоких желобках по поверхности мышечной мембраны. Мембрана мышечной клетки - сарколемма - образует множество глубоких складок, называемых постсинаптическими складками (рис. 16.11). Цитоплазма окончаний мотонейрона сходна с содержимым синаптической бляшки и во время стимуляции освобождает ацетилхолин с помощью того же механизма, о котором говорилось выше. Изменения конфигурации молекул - рецепторов, находящихся на поверхности сарколеммы, ведут к изменению ее проницаемости для Na + и К + , и в результате происходит местная деполяризация, называемая потенциалом концевой пластинки (ПКП). Эта деполяризация по величине вполне достаточна для возникновения потенциала действия, который распространяется по сарколемме в глубь волокна по системе поперечных трубочек (Т-системе) (разд. 17.4.7) и вызывает сокращение мышцы.

Рис. 16.11. Схема строения двигательной концевой пластинки и нервно-мышечного соединения. В верхней части рисунка (над линией Х-Х) изображено окончание аксона моторного нейрона на поверхности мышцы - двигательная концевая пластинка. В нижней части (под линией XX) показана ультраструктура нервно-мышечного соединения
Основная функция межнейронных синапсов и нервно-мышечных соединений состоит в передаче сигнала от рецепторов к эффекторам. Кроме того, строение и организация этих участков химической секреции обусловливают ряд важных особенностей проведения нервного импульса, которые можно резюмировать следующим образом:
1. Однонаправленность передачи. Высвобождение медиатора из пресинаптической мембраны и локализация рецепторов на постсинаптической мембране допускают передачу нервных сигналов по данному пути только в одном направлении, что обеспечивает надежность работы нервной системы.
2. Усиление. Каждый нервный импульс вызывает освобождение в нервно-мышечном синапсе достаточного количества ацетилхолина, чтобы вызвать распространяющийся ответ в мышечном волокне. Благодаря этому нервные импульсы, приходящие к нервно-мышечному соединению, как бы они ни были слабы, могут вызвать реакцию эффектора, и это повышает чувствительность системы.
3. Адаптация, или аккомодация. При непрерывной стимуляции количество освобождающегося в синапсе медиатора постепенно уменьшается до тех пор, пока запасы медиатора не будут истощены; тогда говорят, что синапс утомлен, и дальнейшая передача им сигналов тормозится. Адаптивное значение утомления состоит в том, что оно предотвращает повреждение эффектора вследствие перевозбуждения. Адаптация имеет место также на уровне рецепторов. (См. описание в разд. 16.4.2.)
4. Интеграция. Постсинаптический нейрон может получать сигналы от большого числа возбуждающих и тормозных пресинаптических нейронов (синаптическая конвергенция); при этом постсинаптический нейрон способен суммировать сигналы от всех пресинаптических нейронов. Благодаря пространственной суммации нейрон интегрирует сигналы, поступающие из многих источников, и выдает координированный ответ. В некоторых синапсах имеет место облегчение, состоящее в том, что после каждого стимула синапс становится более чувствительным к следующему стимулу. Поэтому следующие друг за другом слабые стимулы могут вызывать ответ, и это явление используется для повышения чувствительности определенных синапсов. Облегчение нельзя рассматривать как временную суммацию: здесь происходит химическое изменение постсинаптической мембраны, а не электрическая суммация постсинаптических мембранных потенциалов.
5. Дискриминация. Временная суммация в синапсе позволяет отфильтровывать слабые фоновые импульсы, прежде чем они достигнут мозга. Например, экстероцепторы кожи, глаз и ушей постоянно получают из окружающей среды сигналы, не имеющие особого значения для нервной системы: для нее важны лишь изменения интенсивности стимулов, приводящие к увеличению частоты импульсов, которое обеспечивает их передачу через синапс и надлежащую реакцию.
6. Торможение. Передача сигналов через синапсы и нервно-мышечные соединения может затормаживаться определенными блокирующими агентами, воздействующими на постсинаптическую мембрану (см. ниже). Возможно и пресинаптическое торможение, если на окончании аксона чуть выше данного синапса оканчивается другой аксон, образующий здесь тормозный синапс. При стимуляции такого тормозного синапса уменьшается число синаптических пузырьков, разряжающихся в первом, возбуждающем синапсе. Такое устройство позволяет изменять воздействие данного пресинаптического нейрона с помощью сигналов, приходящих от другого нейрона.
Химические вещества выполняют в нервной системе множество различных функций. Воздействия одних веществ широко распространены и хорошо изучены (как, например, возбуждающее действие ацетилхолина и адреналина), тогда как эффекты других носят локальный характер и пока еще недостаточно ясны. Некоторые вещества и их функции приведены в табл. 16.2.

Таблица 16.2. Вещества, влияющие на синапсы и нервно- мышечные соединения млекопитающих
Полагают, что некоторые лекарственные препараты, используемые при таких психических нарушениях, как тревожность и депрессия, воздействуют на химическую передачу в синапсах. Многие транквилизаторы и седативные средства (трициклический антидепрессант имипрамин, резерпин, ингибиторы моноаминоксидазы и др.) оказывают свой лечебный эффект, взаимодействуя с медиаторами, их рецепторами или отдельными ферментами. Так, например, ингибиторы моноаминоксидазы подавляют фермент, участвующий в расщеплении адреналина и норадреналина, и скорее всего оказывают свой лечебный эффект при депрессии, увеличивая продолжительность действия этих медиаторов. Галлюциногены типа диэтиламида лизерговой кислоты и мескалина, воспроизводят действие каких-то природных медиаторов мозга или же подавляют действие других медиаторов.
Проводившееся недавно изучение действия некоторых болеутоляющих веществ - опиатов героина и морфина - показало, что в мозгу млекопитающих присутствуют природные (эндогенные) вещества, вызывающие сходный эффект. Все эти вещества, взаимодействующие с опиатными рецепторами, получили общее название эндорфинов. К настоящему времени открыто много таких соединений; из них лучше всего изучена группа относительно небольших пептидов, называемых энкефалинами (мет-энкефалин, β-эндорфин и др.). Считается, что они подавляют болевые ощущения, влияют на эмоции и имеют отношение к некоторым психическим заболеваниям.
Все это открыло новые пути для изучения функций мозга и биохимических механизмов, лежащих в основе воздействия на боль и лечения с помощью таких различных методов, как внушение, гипно? и акупунктура. Предстоит выделить еще много других веществ типа эндорфинов, установить их строение и функции. С их помощью можно будет получить более полное представление о работе мозга, и это лишь вопрос времени, так как методы выделения и анализа веществ, присутствующих в столь малых количествах, непрерывно совершенствуются.
Волокна скелетных мышц иннервируются толстыми миелинизированными нервными волокнами, исходящими от крупных мотонейронов передних рогов спинного мозга. Как отмечалось в главе 6, каждое нервное волокно после вхождения в брюшко мышцы ветвится и иннервирует от 3 до нескольких сотен волокон скелетной мышцы. Каждое нервное окончание в области средней части мышечного волокна образует синапс, называемый нервно-мышечным соединением. Потенциал действия, возникающий под влиянием нервного сигнала в мышечном волокне, распространяется в обе стороны к его концам. На каждом мышечном волокне, за исключением примерно 2%, формируется только один синапс.
Физиологическая анатомия нервно-мышечного соединения. Двигательная концевая пластинка. На рисунке показано нервно-мышечное соединение между толстым миелинизированным нервным волокном и мышечной клеткой. Нервное волокно формирует совокупность разветвляющихся нервных окончаний, которые погружаются в поверхность мышечного волокна, оставаясь, однако, снаружи его плазматической мембраны. Всю структуру называют двигательной концевой пластинкой. Она покрыта одной или несколькими шванновскими клетками, которые изолируют пластинку от окружающей жидкости.
На рисунке показана схема электронной микрофотографии синапса между одиночным окончанием аксона и мембраной мышечного волокна. Углубление мышечной мембраны называют синаптичсским желобком, или синоптической впадиной, а пространство между мембранами нервного окончания и мышечного волокна — синоптическим пространством, или синоптической щелью. Ширина этого пространства — 20-30 нм. На дне желобка имеется множество более мелких складок мембраны мышечного волокна, называемых субневральными углублениями. Их наличие существенно увеличивает площадь поверхности, на которую может действовать синаптический медиатор.

В окончании аксона содержится большое количество митохондрий, поставляющих АТФ — источник энергии, необходимой для синтеза возбуждающего медиатора ацетилхолино, который, в свою очередь, возбуждает мембрану мышечного волокна. Ацетилхолин синтезируется в цитоплазме окончания и быстро абсорбируется в многочисленные мелкие синаптические везикулы (пузырьки), примерно 300000 которых в норме находятся в нервных окончаниях одной концевой пластинки. В синаптической щели много ацетилхолинэстеразы — фермента, разрушающего ацетилхолин в течение нескольких миллисекунд после его выделения из синаптических пузырьков.
Когда нервный импульс достигает нервно-мышечного соединения, около 125 везикул ацетилхолина раскрываются из терминалей в синаптическое пространство. Некоторые детали этого механизма можно увидеть на рисунке, где представлено увеличенное изображение синаптического пространства с мембраной нервного окончания над ним и мышечной мембраной с ее субневральными углублениями под ним.
На внутренней поверхности нервной мембраны расположены линейные плотные полоски, поперечный срез которых виден на рисунке. По сторонам каждой плотной полоски расположены белковые частицы, пронизывающие мембрану нервного волокна, — электроуправляемые кальциевые каналы. Когда потенциал действия распространяется по терминали, каналы открываются, позволяя ионам кальция диффундировать из синаптического пространства внутрь нервного окончания. Ионы кальция, в свою очередь, как полагают, оказывают особое влияние на ацетилхолиновые везикулы, в результате они притягиваются к мембране нервного волокна, прилежащей к плотным полоскам. Затем везикулы сливаются с мембраной и выделяют ацетилхолин в синаптическое пространство путем экзоцитоза.
Некоторые из деталей этого процесса гипотетичны, однако точно известно, что действенным стимулом, вызывающим выделение ацетилхолина из везикул, является вход ионов кальция и что ацетилхолин из везикул выделяется через мембрану нервного волокна, прилежащую к плотным полоскам.
Соединение Нервно-Мышечное (Neuromuscular Junction), Синапс Нервно-Мышечный (Myoneural Junction)
зона контакта двигательного окончания и мышечного волокна, которое иннервируется им. Каждое мышечное волокно иннервируется веточкой аксона двигательного нейрона, который, оканчиваясь на волокне, образует двигательную концевую пластинку. Структура, соединяющая нервное окончание и мышечное волокно - аксо-мышечный синапс - состоит из пресинаптической мембраны (плазматическая мембрана нервного окончания) и постсинаптической мембраны (плазматическая мембрана мышечного волокна), разделенных синаптической щелью, куда из нервного окончания выделяется нейромедиатор, вызывая сокращение мышцы.
15. Нейромедиаторы, их строение и функции, образование и метаболизм. Классификация нейромедиаторов.
Нейромедиа́торы (нейротрансмиттеры, посредники) — биологически активные химические вещества, посредством которых осуществляется передача электрического импульса с нервной клетки через синаптическое пространство между нейронами. Нервный импульс, поступающий в пресинаптическое окончание, вызывает освобождение в синаптическую щель медиатора. Молекулы медиаторов реагируют со специфическими рецепторными белками клеточной мембраны, инициируя цепь биохимических реакций, вызывающих изменение трансмембранного тока ионов, что приводит к деполяризации мембраны и возникновению потенциала действия. Нейромедиаторы являются, как и гормоны, первичными мессенджерами, но их высвобождение и механизм действия в химических синапсах сильно отличается от такового гормонов. В пресинаптической клетке везикулы, содержащие нейромедиатор, высвобождают его локально в очень маленький объём синаптической щели. Высвобожденный нейромедиатор затем диффундирует через щель и связывается с рецепторами на постсинаптической мембране. Диффузия является медленным процессом, но пересечение такой короткой дистанции, которая разделяет пре- и постсинаптические мембраны (0,1 мкм или меньше), происходит достаточно быстро и позволяет осуществлять быструю передачу сигнала между нейронами или между нейроном и мышцей.Недостаток какого-либо из нейромедиаторов может вызывать разнообразные нарушения, например, различные виды депрессии. Также считается, что формирование зависимости от наркотиков и табака связано с тем, что при употреблении этих веществ задействуются механизмы производства нейромедиатора серотонина, а также других нейромедиаторов, блокирующие (вытесняющие) аналогичные естественные механизмы.
Аминокислоты (и их производные). К ним относят таурин, норадреналин, ДОФАминГАМК, глицин, ацетилхолин, гомоцистеин и некоторые другие (адреналин, серотонин, гистамин, серотонин).
Таурин. Таурин образуется из аминокислоты цистеина. Сначала происходит окисление серы в SH-группе до остатка серной кислоты (процесс идет в несколько стадий), а затем происходит декарбоксилирование. Таурин - это необычная кислота, в которой нет карбоксильной группы, а имеется остаток серной кислоты.
Таурин принимает участие в проведении нервного импульса в процессе зрительного восприятия.
Ацетилхолин. Для синтеза холина требуются аминокислоты серин, метионин. Этаноламин может быть использован и в готовом виде. Но, как правило, из крови в нервную ткань поступает уже готовый холин. Второй же предшественник этого нейромедиатора - Ацетил-КоА, синтезируется в нервных окончаниях.
Продукт этой реакции ацетилхолин участвует в синаптической передаче нервного импульса. Он накапливается в синаптических пузырьках, образуя комплексы с отрицательно заряженным белком везикулином. Передача возбуждения с одной клетки на другую осуществляется с помощью специального синаптического механизма.
Синапс – это функциональный контакт специализированных участков плазматических мембран двух возбудимых клеток. Синапс состоит из пресинаптической мембраны, синаптической щели и постинаптической мембраны. Мембраны клеток в месте контакта имеют утолщения в виде бляшек – нервных окончаний. Нервный импульс, достигший нервного окончания, не в состоянии преодолеть возникшее перед ним препятствие - синаптическую щель. После этого электрический сигнал преобразуется в химический. Пресинаптическая мембрана содержит специальные канальные белки, подобные белкам, формирующим натриевый канал в мембране аксона. Они тоже реагируют на мембранный потенциал, изменяя свою конформацию и формируют канал. В результате ионы Са2+ проходят через пресинаптическую мембрану по градиенту концентраций в нервное окончание. Градиент концентраций Са2+ создается работой Са2+-зависимой.
АТФазы – кальциевым насосом. Повышение концентрации Са2+ внутри нервного окончания вызывает слияние 200-300 имеющихся там везикул, заполненных ацетилхолином, с плазматической мембраной. Далее ацетилхолин секретируется в синаптическую щель путем экзоцитоза, и присоединяется к рецепторным белкам, расположенным на поверхности постсинаптической мембраны.
Ацетилхолиновый рецептор представляет собой трансмембранный олигомерный гликопротеиновый комплекс, состоящий из 6 субъединиц: 2-бета, 1-гамма и 1-дельта. Плотность расположения белков-рецепторов в постсинаптической мембране очень велика - около 20000 молекул на 1 мкм2. Пространственная структура рецептора строго сооответствует конформации медиатора.
Во время гидролиза ацетилхолина образуется промежуточный фермент-субстратный комплекс, в котором ацетилхолин связан с активным центром фермента через серин.
Необратимое ингибирование холинэстеразы вызывает смерть. Ингибиторами холинэстеразы являются фосфорорганические соединения (хлорофос, дихлофос, табун, зарин, зоман, бинарные яды). Эти вещества связываются ковалентно с серином в активном центре фермента. Некоторые из них синтезированы в качестве инсектицидов, а некоторые – в качестве боевых отравляющих веществ (нервно-паралитические яды). Смерть наступает в результате остановки дыхания.
Обратимые ингибиторы холинэстеразы используются как лечебные препараты. Например, при лечении глаукомы и атонии кишечника.
Катехоламины: норадреналин и дофамин. Адренэргические синапсы встречаются в постганглионарных волокнах, в волокнах симпатической нервной системы, в различных отделах головного мозга. Катехоламины в нервной ткани синтезируются по общему механизму из тирозина. Ключевой фермент синтеза – тирозингидроксилаза, ингибируемая конечными продуктами.
Норадреналин – медиатор в постганглионарных волокнах симпатической и в различных отделах ЦНС.
Дофамин – медиатор проводящих путей, тела нейронов которого расположены в отделе мозга, который отвечает за контроль произвольных движений. Поэтому при нарушении дофаминэргической передачи возникает заболевание паркинсонизм.
Катехоламины, как и ацетилхолин, накапливаются в синаптических пузырьках и тоже выделяется в синаптическую щель при поступлении нервного импульса. Но регуляция в адренэргическом рецепторе происходит иначе. В пресинаптической мембране здесь имеется специальный регуляторный белок - ахромогранин (Мм = 77 кДа), который в ответ на повышение концентрации медиатора в синаптической щели связывает уже выделившийся медиатор и прекращает его дальнейший экзоцитоз. Фермента, разрушающего медиатор, в адренэргических синапсах нет. После передачи импульса молекулы медиатора перекачивается специальной транспортной системой путем активного транспорта с участием АТФ обратно через пресинаптическую мембрану и включается вновь в везикулы. В пресинаптическом нервном окончании излишек медиатора может быть инактивирован моноаминоксидазой, а также катехоламин-О-метилтрансферазой путем метилирования по оксигруппе. Кокаин тормозит активный транспорт катехоламинов.
ГАМК – тормозной медиатор. Повышает проницаемость постсинаптических мембран для ионов калия. Это ведет к изменению мембранного потенциала.
Глицин – тормозной медиатор, по вызываемым эффектам подобен гамк.
Пептиды. Имеют в своем составе от трех до нескольких десятков аминокислотных остатков. Функционируют только в высших отделах нервной системы.
Эти пептиды, как и катехоламины, выполняют функцию не только нейромедиаторов, но и гормонов. Передают информацию от клетки к клетке по системе циркуляции.
1. нейрогипофизарные гормоны (вазопрессин, либерины, статины). Эти вещества одновременно и гормоны и медиаторы;
2. гастроинтестинальные пептиды (гастрин, холецистокинин). Гастрин вызывает чувство голода, холецистокинин вызывает чувство насыщения, а также стимулирует сокращение желчного пузыря и функцию поджелудочной железы;
3. опиатоподобные пептиды (или пептиды обезболивания). Образуются путем реакций ограниченного протеолиза белка-предшественника проопиокортина. Взаимодействуют с теми же рецепторами, что и опиаты (например, морфин), тем самым имитируют их действие. Общее название - эндорфины - вызывают обезболивание. Они легко разрушаются протеиназами, поэтому их фармакологический эффект незначителен;
4. пептиды сна. Их молекулярная природа не установлена. Известно лишь, что их введение животным вызывает сон;
5. пептиды памяти (скотофобин). Накапливается в мозге крыс при тренировке на избегание темноты;
6. пептиды - компоненты ренин-ангиотензиновой системы. Показано, что введение ангиотензина-II в центр жажды головного мозга вызывает появление этого ощущения и стимулирует секрецию антидиуретического гормона.
16. Нейроглия. Источники онтогенетического развития нейроглии. Нейроглия (от нейро. и греч. gl?a — клей), глия, клетки в мозге, своими телами и отростками заполняющие пространства между нервными клетками — нейронами — и мозговыми капиллярами. Каждый нейрон окружен несколькими клетками Н., которая равномерно распределена по всему мозгу и составляет около 40% его объёма. Клетки Н. — число их в центральной нервной системе (ЦНС) млекопитающих около 140 млрд. — мельче нейронов в 3—4 раза и отличаются от них по морфологическим и биохимическим признакам. С возрастом количество нейронов в ЦНС уменьшается, а клеток Н. — увеличивается, т.к. последние, в отличие от нейронов, сохраняют способность к делению. Основные функции Н.: создание между кровью и нейронами гемато-энцефалического барьера, необходимого как для защиты нейронов, так и главным образом для регуляции поступления веществ в ЦНС и их выведения в кровь; обеспечение реактивных свойств нервной ткани (образование рубцов после травмы, участие в реакциях воспаления, в образовании опухолей и др.). Различают астроглию, олигоглию, или олигодендроглию, и эпендиму, которые вместе составляют макроглию, а также микроглию, занимающую особое положение среди клеток Н.
18.Строение и функции эпендимы. Эпендима – выстилает все внутренние полости в головном и спинном мозге. Выполняет роль барьера между веществом мозга и омывающей его спинномозговой жидкостью; регулирует секрецию и состав спинномозговой жидкости. Эпендима - клетки в мозге животных и человека, выполняющие в центральной нервной системе разграничительную, опорную и секреторную функции; форма нейроглии. Э. дифференцируется из клеток нервной трубки в раннем эмбриогенезе. Клетки Э. (эпендимоциты) выстилают стенки спинномозгового канала и желудочков головного мозга. Их тела вытянуты, на свободном конце — реснички (теряемые во многих отделах мозга после рождения особи), биение которых способствует циркуляции спинномозговой жидкости. От противоположного конца эпендимоцита в мозг отходит длинный, ветвящийся отросток. Э. стенок 3-го желудочка мозга (ее клетки называются таницитами), возможно, осуществляет обмен биологически активными веществами между нейронами прилегающих областей мозга, спинномозговой жидкостью и сосудами воротной системы гипофиза.
19. Строение, функции и происхождение микроглии. Микроглиоциты, или микроглия, представляют собой мелкие клетки, рассеянные в центральной нервной системе. При травмах или дегенерации нервной ткани они способны мигрировать к очагу повреждения, где превращаются в крупные макрофаги, поглощающие путем фагоцитоза продукты распада. Таким образом, микроглиоциты препятствуют развитию воспалительных процессов и распространению инфекции в нервной ткани. Микроглия – белое в-во головного и спинного мозга преимущественно около кровеносных сосудов. Выполняет защитную роль, сходную с ролью макрофагов; предотвращает попаданию в нервную систему чужеродных субстанций. Клетки микроглии происходят из мезодермы. Как видно из названия, они отличаются небольшими размерами. Эти клетки могут активно передвигаться и выполнять фагоцитарные функции. Благодаря способности к активной миграции микроглия распределена по всей центральной нервной системе.
Читайте также:
