
Какие химические элементы участвуют в проведении нервного импульса

Кандидат биологических наук Л. Чайлахян, научный сотрудник Института биофизики АН СССР
Мозг человека, без сомнения, высшее достижение природы.
Велика и заманчива цель, но неимоверно сложен объект исследования. Шутка сказать, этот килограмм ткани представляет собой сложнейшую систему связи десятков миллиардов нервных клеток.
Однако первый существенный шаг к познанию работы мозга уже сделан. Может быть, он один из самых легких, но он чрезвычайно важен для всего дальнейшего.
Я имею в виду исследование механизма передачи нервных импульсов — сигналов, бегущих по нервам, как по проводам. Именно эти сигналы являются той азбукой мозга, с помощью которой органы чувств посылают в центральную нервную систему сведения-депеши о событиях во внешнем мире. Нервными импульсами зашифровывает мозг свои приказы мышцам и различным внутренним органам. Наконец, на языке этих сигналов говорят между собой отдельные нервные клетки и нервные центры.
В проблеме изучения механизма нервного импульса и его распространения можно выделить два основных вопроса: природа проведения нервного импульса или возбуждения в пределах одной клетки — по волокну и механизм передачи нервного импульса от клетки к клетке — через синапсы.
Какова природа сигналов, передающихся от клетки к клетка по нервным волокнам?
Этой проблемой человек интересовался уже давно, Декарт предполагал, что распространение сигнала связано с переливанием жидкости по нервам, как по трубкам. Ньютон думал, что это чисто механический процесс. Когда появилась электромагнитная теория, ученые решили, что нервный импульс аналогичен движению тока по проводнику со скоростью, близкой к скорости распространения электромагнитных колебаний. Наконец, с развитием биохимии появилась точка зрения, что движение нервного импульса — это распространение вдоль по нервному волокну особой биохимической реакции.
И всё же ни одно из этих представлений не оправдалось.
В настоящее время природа нервного импульса раскрыта: это удивительно тонкий электрохимический процесс, в основе которого лежит перемещение ионов через оболочку клетки.
Большой вклад в раскрытие этой природы внесли работы трех ученых: Алана Ходжкина, профессора биофизики Кембриджского университета; Эндрью Хаксли, профессора физиологии Лондонского университета, и Джона Экклса, профессора физиологии австралийского университета в Канберре. Им присуждена Нобелевская премия в области медицины за 1963 год,
Впервые предположение об электрохимической природе нервного импульса высказал известный немецкий физиолог Бернштейн в начале нашего столетия.
К началу двадцатого века было довольно многое известно о нервном возбуждении. Ученые уже знали, что нервное волокно можно возбудить электрическим током, причем возбуждение всегда возникает под катодом — под минусом. Было известно, что возбужденная область нерва заряжается отрицательно по отношению к невозбужденному участку. Было установлено, что нервный импульс в каждой точке длится всего 0,001—0,002 секунды, что величина возбуждения не зависит от силы раздражения, как громкость звонка в нашей квартире не зависит от того, как сильно мы нажимаем на кнопку. Наконец, ученые установили, что носителями электрического тока в живых тканях являются ионы; причем внутри клетки основной электролит — соли калия, а в тканевой жидкости — соли натрия. Внутри большинства клеток концентрация ионов калия в 30—50 раз больше, чем в крови и в межклеточной жидкости, омывающей клетки.
И вот на основании всех этих данных Бернштейн предположил, что оболочка нервных и мышечных клеток представляет собой особую полупроницаемую мембрану. Она проницаема только для ионов К + ; для всех остальных ионов, в том числе и для находящихся внутри клетки отрицательно заряженных анионов, путь закрыт. Ясно, что калий по законам диффузии будет стремиться выйти из клетки, в клетке возникает избыток анионов, и по обе стороны мембраны появится разность потенциалов: снаружи — плюс (избыток катионов), внутри — минус (избыток анионов). Эта разность потенциалов получила название потенциала покоя. Таким образом, в покое, в невозбужденном состоянии внутренняя часть клетки всегда заряжена отрицательно по сравнению с наружным раствором.
Бернштейн предположил, что в момент возбуждения нервного волокна происходят структурные изменения поверхностной мембраны, ее поры как бы увеличиваются, и она становится проницаемой для всех ионов. При этом, естественно, разность потенциалов исчезает. Это и вызывает нервный сигнал.
Мембранная теория Бернштейма быстро завоевала признание и просуществовала свыше 40 лет, вплоть до середины нашего столетия.
Но уже в конце 30-х годов теория Бернштейна встретилась с непреодолимыми противоречиями. Сильный удар ей был нанесен в 1939 году тонкими экспериментами Ходжкина и Хаксли. Эти ученые впервые измерили абсолютные величины мембранного потенциала нервного волокна в покое и при возбуждении. Оказалось, что при возбуждении мембранный потенциал не просто уменьшался до нуля, а переходил через ноль на несколько десятков милливольт. То есть внутренняя часть волокна из отрицательной становилась положительной.
Но мало ниспровергнуть теорию, надо заменить ее другой: наука не терпит вакуума. И Ходжкин, Хаксли, Катц в 1949—1953 годах предлагают новую теорию. Она получает название натриевой.
Здесь читатель вправе удивиться: до сих пор о натрии не было речи. В этом все и дело. Ученые установили с помощью меченых атомов, что в передаче нервного импульса замешаны не только ионы калия и анионы, но и ионы натрия и хлора.
В организме достаточно ионов натрия и хлора, все знают, что кровь соленая на вкус. Причем натрия в межклеточной жидкости в 5—10 раз больше, чем внутри нервного волокна.
Что же это может означать? Ученые предположили, что при возбуждении в первый момент резко увеличивается проницаемость мембраны только для натрия. Проницаемость становится в десятки раз больше, чем для ионов калия. А так как натрия снаружи в 5—10 рез больше, чем внутри, то он будет стремиться войти в нервное волокно. И тогда внутренняя часть волокна станет положительной.
А через какое-то время — после возбуждения — равновесие восстанавливается: мембрана начинает пропускать и ионы калия. И они выходят наружу. Тем самым они компенсируют тот положительный заряд, который был внесен внутрь волокна ионами натрия.
Совсем нелегко было прийти к таким представлениям. И вот почему: диаметр иона натрия в растворе раза в полтора больше диаметра ионов калия и хлора. И совершенно непонятно, каким образом больший по размеру ион проходит там, где не может пройти меньший.
Нужно было решительно изменить взгляд на механизм перехода ионов через мембраны. Ясно, что только рассуждениями о порах в мембране здесь не обойтись. И тогда была высказана идея, что ионы могут пересекать мембрану совершенно другим способом, с помощью тайных до поры до времени союзников — особых органических молекул-переносчиков, спрятанных в самой мембране. С помощью такой молекулы ионы могут пересекать мембрану в любом месте, а не только через поры. Причем эти молекулы-такси хорошо различают своих пассажиров, они не путают ионы натрия с ионами калия.
Интересно, что нервные волокна тратят на свою основную работу — проведение нервных импульсов — всего около 15 минут в сутки. Однако готовы к этому волокна в любую секунду: все элементы нервного волокна работают без перерыва — 24 часа в сутки. Нервные волокна в этом смысле подобны самолетам-перехватчикам, у которых непрерывно работают моторы для мгновенного вылета, однако сам вылет может состояться лишь раз в несколько месяцев.
Мы познакомились сейчас с первой половиной таинственного акта прохождения нервного импульса — вдоль одного волокна. А как же передается возбуждение от клетки к клетке, через места стыков — синапсы. Этот вопрос был исследован в блестящих опытах третьего нобелевского лауреата, Джона Экклса.
Возбуждение не может непосредственно перейти с нервных окончаний одной клетки на тело или дендриты другой клетки. Практически весь ток вытекает через синаптическую щель в наружную жидкость, и в соседнюю клетку через синапс попадает ничтожная его доля, неспособная вызвать возбуждение. Таким образом, в области синапсов электрическая непрерывность в распространении нервного импульса нарушается. Здесь, на стыке двух клеток, в силу вступает совершенно другой механизм.
Когда возбуждение подходит к окончанию клетки, к месту синапса, в межклеточную жидкость выделяются физиологически активные вещества — медиаторы, или посредники. Они становятся связующим звеном в передаче информации от клетки к клетке. Медиатор химически взаимодействует со второй нервной клеткой, изменяет ионную проницаемость ее мембраны — как бы пробивает брешь, в которую устремляются многие ионы, в том числе и ионы натрия.
Итак, благодаря работам Ходжкина, Хаксли и Экклса важнейшие состояния нервной клетки — возбуждение и торможение — можно описать в терминах ионных процессов, в терминах структурно-химических перестроек поверхностных мембран. На основании этих работ уже можно делать предположения о возможных механизмах кратковременной и долговременной памяти, о пластических свойствах нервной ткани. Однако это разговор о механизмах в пределах одной или нескольких клеток. Это лишь, азбука мозга. По-видимому, следующий этап, возможно, гораздо более трудный, — вскрытие законов, по которым строится координирующая деятельность тысяч нервных клеток, распознание языка, на котором говорят между собой нервные центры.
Мы сейчас в познании работы мозга находимся на уровне ребенка, который узнал буквы алфавита, но не умеет связывать их в слова. Однако недалеко время, когда ученые с помощью кода — элементарных биохимических актов, происходящих в нервной клетке, прочтут увлекательнейший диалог между нервными центрами мозга.
Детальное описание иллюстраций
Проведение нервных импульсов осуществляется при помощи отростков нервных клеток – нервных волокон. По особенностям строения и расположения осевых цилиндров в составе нерва (по отношению к шванновским клеткам) различают волокна мякот-ного (миелинизированные) и безмякотного типа. Как правило, нервы включают волокна обоих типов в разном количественном соотношении. Миелиновые волокна преобладают в чувствительных и двигательных нервах органов чувств, скелетных мышц и вегетативной нервной системы. Безмиелиновые преимущественно встречаются в составе нервов симпатического отдела.
Волокна мякотного и безмякотного типов состоят из одинаковых структурных компонентов: отростка нервной клетки (осевого цилиндра), шванновских клеток (леммоцитов) и базальной мембраны (тонкой пластинки, состоящей из рыхлой волокнистой соединительной ткани и покрывающей каждое нервное волокно снаружи). Миелиновое волокно представлено осевым цилиндром, окруженным миелиновой оболочкой. Шванновские клетки, окружающие отросток нейроцита, образуют дупликатуры цитоплазма-тической мембраны, называемые мезаксонами. Многократно обертываясь вокруг осевого цилиндра, мезаксон формирует миелиновую оболочку нервного волокна, представляющую собой плотный липидно-белковый футляр. На протяжении мякотного нервного волокна через примерно равные интервалы имеются участки истончения миелинового слоя, ширина которых составляет около 1 мкм. Эти участки получили название перехватов Ранвье. Они имеют большое функциональное значение при проведении нервного импульса. Расстояние от одного до другого перехвата называется межузловым сегментом. Мякотные волокна могут располагаться как в пределах центральной нервной системы, так и вне ее. В зависимости от этого среди них различают центральные и периферические. Миелиновые волокна характеризуются высокой скоростью проведения импульсов (60-120 м/с). Эта особенность обусловлена изолирующей функцией миелино-вого слоя и наличием перехватов Ранвье.
Безмякотные нервные волокна построены более примитивно. Они представлены цепочкой шванновских клеток, в которую вдавлено от одного и более осевых цилиндров. В среднем их количество составляет от 5 до 20. Миелинового слоя в этих клетках нет, леммоциты образуют между осевыми цилиндрами мезаксо-ны, не играющие существенной роли в проведении нервного возбуждения. Отростки нейроцитов расположены довольно рыхло и плохо изолированы. Поэтому они могут переходить из одного безмиелинового волокна в другое. Снаружи волокно покрыто соединительно-тканной базальной пластинкой. В связи с перечисленными особенностями строения безмякотные нервные волокна называют волокнами кабельного типа. Скорость проведения в них (3–5 м/с) гораздо ниже, чем в миелиновых.
Функцией нервного волокна является проведение нервных импульсов, способ которого в мякотных и безмякотных нервных волокнах несколько различается. В этом процессе большое значение имеют структуры нервного волокна.
Главный элемент с функциональной точки зрения – цито-плазматическая мембрана осевого цилиндра. Именно на мембране возникает нервный импульс, а затем распространяется по ней. Миелин одновременно служит изолятором и выполняет трофическую функцию по отношению к осевому цилиндру. Являясь веществом липидно-белковой природы, миелин благодаря своему высокому сопротивлению ограничивает передвижение ионов. В связи с этим потенциал действия, возникновение которого связано с перераспределением ионов, может возникать не на любом участке волокна, а в местах наименьшего сопротивления – истончениях миелина, перехватах Ранвье. Это имеет большое значение в проведении нервного возбуждения и обуславливает разницу механизмов проведения в мякотных и безмякотных волокнах. Трофическая функция миелиновой оболочки заключается в регуляции роста и обмена веществ осевого цилиндра. Кроме того, шванновские клетки при повреждении нерва осуществляют дегенерацию дистальной от тела клетки отсеченной части осевого цилиндра, а затем образуют ложе для нового волокна, обеспечивая направление продвижения колбы роста.
Свою роль в распространении импульса играют и внутриклеточные структуры осевого цилиндра, в частности нейрофибрил-лы, микротубулы и транспортные филаменты, соединяющиеся с различными веществами или органеллами и осуществляющие их транспорт по нервным волокнам.
Проведение импульса протекает при определенных условиях и подчиняется ряду законов. Во-первых, проведение нервного импульса по волокну может осуществляться полноценно лишь при условии его анатомической и физиологической целостности, или непрерывности. Анатомическая целостность может нарушаться при механических повреждениях, например при перерезке или сдавливании волокна, нарушение физиологической целостности может быть результатом сильного охлаждения, действия веществ – блокаторов натриевых каналов мембраны (местных анестетиков) или других подобных факторов. Все это вызывает частичное или полное нарушение проводимости нервного волокна.
С помощью многочисленных опытов (А. И. Бабухин, 1877 г., В. Кюне, 1886 г.) было доказано, что возникшее на мембране возбуждение распространяется в двух направлениях: в центробежном и в центростремительном. Потенциал действия возникает на нейроне в так называемой триггерной зоне (в месте перехода тела нервной клетки в нейрит), и распространяется по аксону и по телу к дендритам. В условиях целостного организма наблюдать двустороннее проведение нельзя из-за наличия клапанного аппарата синаптических соединений, передающих импульс только в одном направлении, и специфики места возникновения возбуждения (рецепторного аппарата).
В одном нерве могут находиться волокна разных видов – двигательные, чувствительные, вегетативные – различающиеся иннервируемыми структурами. В связи с этим очень важным свойством является изолированное проведение возбуждения в нервных волокнах. В первую очередь, роль изолятора выполняет миелиновая оболочка, обладающая высоким сопротивлением. Большое значение имеет также наличие жидкости в межклеточных пространствах. За счет более низкого ее сопротивления по сравнению с мембраной волокна ток между деполяризованным и покоящимся участками мембраны идет по межклеточным щелям и не затрагивает другие волокна.
Таким образом, проведение нервного импульса подчиняется трем основным законам: закону анатомической и физиологической целостности, двустороннего проведения и изолированного проведения возбуждения.
Возникновение его начинается со снятия потенциала покоя. Потенциалом покоя, или мембранным потенциалом, называют разницу потенциалов между наружной (положительно заряженной) и внутренней (отрицательно заряженной) поверхностями клеточной мембраны в состоянии относительного физиологического покоя. Эта величина составляет примерно 60–90 мВ.
В соответствии с мембранно-ионной теорией, предложенной в 1902 г. немецким физиологом Н. А. Берштейном, в поддержании постоянства потенциала покоя участвует ряд факторов.
Среди них называют в первую очередь особенности строения и функционирования клеточных мембран. Разность потенциалов существует за счет различного содержания ионов натрия, калия, кальция и хлора в клетке и вне ее и, следовательно, различной проницаемости мембраны для них. Важным свойством цитоплаз-матической мембраны является наличие в ней белков (60 %); среди них различают периферические, погруженные (полуинтегральные) и трансмембранные (интегральные), которые помимо рецепторной и ферментативной функции выполняют транспортную функцию и образуют поры, предназначенные для перемещения определенных ионов.
Поры, или каналы, имеют диаметр около 1 нм, образованы молекулами белков и работают в соответствии с воротным механизмом, регуляция которого обеспечивается разностью потенциалов или взаимодействием белка-рецептора с адекватным медиатором.
В соответствии с последним, различают поры электровозбудимые (преобладают в нервных и мышечных клетках) и хемовозбу-димые. Функционирование воротного механизма представляет собой последовательную смену состояний канала. В закрытом состоянии, которое существует в относительном физиологическом покое, пора не может пропустить ион. Из закрытого состояния канал переходит в открытое, соответствующее началу деполяризации. Это, собственно, время работы поры. После открытого следует состояние инактивации, когда канал все еще открыт, но ион не пропускает. После этого канал вновь закрывается.
Поры могут осуществлять транспорт только определенного иона, таким образом, обладая специфичностью (избирательностью), что объясняется разницей или сходством диаметра иона и канала, зарядом внутри него и вышеописанным воротным механизмом.
В покое клеточная мембрана обладает хорошей проницаемостью для ионов калия и хлора. Проницаемость для натрия гораздо ниже. Анионы и молекулы органических веществ не могут проникнуть через мембрану.
Следующим фактором, обусловливающим наличие и постоянство величины мембранного потенциала, является ионная асимметрия внутри клетки и снаружи. В клетке содержатся преимущественно ионы калия и анионы органических веществ, а вне ее больше ионов натрия, хлора и кальция. Ионную асимметрию можно считать главной причиной существования мембранного потенциала. Она поддерживается за счет избирательной проницаемости цитоплазматической мембраны и физико-химического равновесия Доннана.
Большое значение в поддержании мембранного потенциала имеют так называемые пассивные силы, объединяющие силы простой диффузии и электростатического взаимодействия ионов. Пассивными их называют потому, что для своей реализации они не требуют энергетических затрат. Действие этих сил можно проследить, например, на ионе калия. Клеточная мембрана хорошо проницаема для калия, в клетке содержание калия больше, чем вне ее. По градиенту концентрации калий выходит из клетки. Он заряжен положительно, следовательно, выходя на поверхность, он попадает под действие сил электростатического отталкивания, так как в состоянии относительного физиологического покоя наружная мембрана также заряжена положительно, и снова возвращается в клетку. Таким образом, калий находится преимущественно внутри нее.
Для хлора, находящегося большей частью снаружи, клеточная мембрана тоже легко проницаема. Аналогично калию, на хлор действуют силы простой диффузии, под воздействием которых он устремляется в клетку. Отрицательный заряд внутренней поверхности мембраны отталкивает анионы, и хлор остается снаружи.
Клеточная мембрана не пропускает ионы натрия, находящегося вне клетки, и анионы органических веществ (внутри), они не могут пройти через нее при помощи диффузии и соответственно, остаются за пределами клетки и в ней.
Фактически мембранный потенциал основывается на электрохимическом равновесии по иону калия (количество калия, вышедшего из клетки благодаря диффузии и вернувшегося путем электростатического отталкивания, одинаково). Это равновесие развивается при условии, если создается равенство сил диффузии и отталкивания в клетке.
Важным звеном в формировании и поддержании мембранного потенциала является работа натрий-калиевого насоса, активного транспорта ионов через цитоплазматическую мембрану против градиента концентрации с затратой энергии. Функционирование насоса происходит при участии натрий-калиевой АТФ-азы, которая за один цикл работы насоса выводит за пределы клетки три иона натрия и возвращает два калия.
При возникновении потенциала действия происходит снятие мембранного потенциала и перезарядка мембраны, что является результатом постепенного изменения проницаемости для определенных ионов. Он способен генерироваться только в определенных участках мембраны нервной клетки, называемых электровозбудимыми. Они располагаются в аксональном холмике (триггерная зона), нейрите, иногда в теле (но не на постсинапти-ческих мембранах), в перехватах Ранвье. Наиболее значимая из этих структур – аксональный холмик. Потенциал действия возникает лишь при достижении критического уровня деполяризации (влияние распространяющегося возбуждающего постсинап-тического потенциала из синапсов или рецепторного потенциала с чувствительных нейронов). Мембрана аксонального холмика содержит большое количество натриевых и калиевых каналов и, кроме того, имеет сравнительно низкий с другими структурами порог возбуждения. Это обусловливает более легкое достижение критического уровня деполяризации. Таким образом, этот участок является наиболее легко возбудимым в клетке.
С точки зрения мембранно-ионной теории, потенциал действия развивается благодаря повышению мембранной проницаемости для двух видов ионов – натрия и калия. Сначала открываются натриевые каналы, проницаемость увеличивается в 400–500 раз, и натрий устремляется в клетку. Для калия проницаемость становится возможна позже, к началу реполяризации, и увеличивается всего в 10–15 раз. В итоге в процессе формирования потенциала действия натрия выходит гораздо больше, чем выходит калия.
Потенциал действия – достаточно сложное комплексное явление, включающее несколько фаз. В первую очередь возникает локальный ответ, затем следует пик, или спайк, состоящий из фазы деполяризации и реполяризации, позже наблюдаются следовые потенциалы – отрицательный и положительный.
Локальный ответ (начальная фаза) возникает, когда сила действующего раздражителя еще не достигла пороговой (подпорого-вый раздражитель). Этот этап характеризуется увеличением ионной проницаемости мембраны для всех ионов (неспецифически). Мембранный потенциал начинает смещаться в положительную сторону. В процессе увеличения силы воздействия при достижении 50–70 % от порога уже имеет место специфическое увеличение проницаемости для натрия, который силами простой диффузии входит в клетку и частично деполяризует мембрану. Мембранный потенциал стремится к критическому уровню деполяризации, который в разных клетках может составлять примерно от 50 до 40 мВ
При достижении раздражителем пороговой силы начинается первый этап фазы спайка: деполяризация (восходящее колено). Мембранный потенциал достигает критического уровня, что вызывает открытие всех натриевых каналов и лавинообразный ток ионов внутрь клетки. Это становится причиной увеличения мембранного потенциала до нуля, а затем и перезарядки мембраны. Деполяризация прекращается после развития нового электрохимического равновесия по натрию и инактивации натриевых каналов.
В процессе реполяризации (нисходящее колено) мембрана начинает пропускать ионы калия. Он покидает клетку, удаляя таким образом положительный заряд. Перераспределение ионов активирует работу натрий-калиевого насоса.
За реполяризацией следует отрицательный следовой потенциал. Он характеризуется повторным незначительным увеличением проницаемости для натрия, который в небольшом количестве входит в клетку и вызывает приближение мембранного потенциала к критическому уровню – частичную деполяризацию. Эта фаза сменяется положительным следовым потенциалом, основным явлением которого называют следовую гиперполяризацию (формирование положительного заряда на наружной поверхности мембраны, возникающее в результате продолжения тока калия из клетки). В итоге величина мембранного потенциала вновь отдаляется от критического уровня.
Таким образом, возбуждение включает локальный ответ, который не распространяется и быстро затухает, и волновой ответ, распространяющийся далее по волокнам.
Осуществляясь в соответствии с одинаковыми закономерностями, процессы проведения в миелиновых и безмиелиновых волокнах отличается рядом особенностей. В безмиелиновых волокнах импульс распространяется от возбужденного участка к соседним, расположенным рядом с ним. Распространение потенциала действия происходит в обе стороны без изменения амплитуды с помощью круговых токов.
Различная скорость проведения, продолжительность фаз потенциала действия, строение волокон позволяет подразделять их на 3 вида: А, В, С.
Волокна типа А – миелиновые, они, в свою очередь, подразделяются еще на 4 подгруппы: α-, β-, γ– и δ-волокна. Альфа-волокна, или, как их еще называют, А-а, среди этой группы имеют самый большой диаметр (12–22 мкм) и высокую скорость проведения импульса (70-120 м/с). В организме они достаточно распространены. К этой группе относятся двигательные и чувствительные волокна скелетной мускулатуры. Что касается длительности различных фаз потенциала действия, то продолжительность пика составляет 0,4–0,5 мс, следовой деполяризации – 15–20 мс, следовой гиперполяризации – 40–60 мс. Следующие подгруппы типа А – А-β, А-γ, А-δ – отличаются меньшим диаметром и скоростью проведения возбуждения, но их потенциал действия более продолжителен, чем А-α. Эти волокна участвуют в основном в проведении импульсов от различных рецепторов внутренних органов в нервные центры. А-β волокна идут от тактильных рецепторов, А-γ – от тактильных и барорецепторов, а также к мышечным веретенам как двигательные волокна. В А-δ подгруппу входят афферентные волокна, несущие импульс от термо-, барорецепторов и ноцицепторов. Проведение в трех последних подгруппах составляет 1-12 м/с. Самое быстрое проведение – в А-β волокнах. Пик потенциала действия может продолжаться до 0,9–1,0 мс, (наиболее длительное – в А-β волокнах). Следовая деполяризация и гиперполяризация продолжаются примерно в течение того же времени, что и в А-α.
Волокна типа В также покрыты миелиновой оболочкой, имеют достаточно низкую скорость проведения и находятся в составе вегетативных отделов нервной системы. В основном, это преганг-лионарные вегетативные волокна. Проведение осуществляется со скоростью от 3 до 18 м/с. Наряду с этим, в В-волокнах потенциал действия по длительности превосходит более чем в 3 раза это значение в А-волокнах. Важнейшим отличием является тот факт, что в этих волокнах не наблюдается фаза следовой гиперполяризации. Следовая гиперполяризация может продолжаться до 100 мс.
Волокна типа С по строению безмиелиновые, очень тонкие, всего 0,5–2,0 мкм в диаметре. Они также обнаруживаются в вегетативной нервной системе в составе постганглионарных волокон, осуществляя проведение от рецепторов тепла, холода, давления и боли. Эти волокна отличаются наиболее медленной скоростью проведения (не больше 3 м/с). Их потенциал действия отличается самой большой (по сравнению с другими типами) длительностью развития потенциала действия: у теплокровных животных и человека – до 2 мс.
Волокна способны существовать и выполнять свою функцию только тогда, когда они связаны с телом нейрона. Повреждение волокна ведет к нарушению или утрате способности проводить возбуждение. Перерезка ведет к гибели отсеченной части волокна. Однако волокна способны регенерировать за счет гипертрофии (эндорепродукции). Разрушение отделенного волокна и образование нового является сложным и длительным процессом и протекает в несколько последовательных стадий. После перерезки миелиновый слой перерождается в жировые капли. Леммо-циты осуществляют дегенерацию осевого цилиндра с помощью гидролитических ферментов лизосом. В результате на месте волокна остается цепь леммоцитов. Дальше начинается регенерация волокна. От места его отделение в ложе, образованной шваннов-скими клетками, начинает прорастать колба роста. Регенерация осуществляется с примерной скоростью 0,5–4,5 мм за сутки в зависимости от строения волокна и местоположения его в организме.
Группа нервных тканей объединяет ткани эктодермального происхождения, которые в совокупности образуют нервную систему и создают условия для реализации ее многочисленных функций. Обладают двумя основными свойствами: возбудимостью и проводимостью.
Структурно-функциональной единицей нервной ткани является нейрон (от др.-греч. νεῦρον — волокно, нерв) - клетка с одним длинным отростком - аксоном, и одним/несколькими короткими - дендритами.
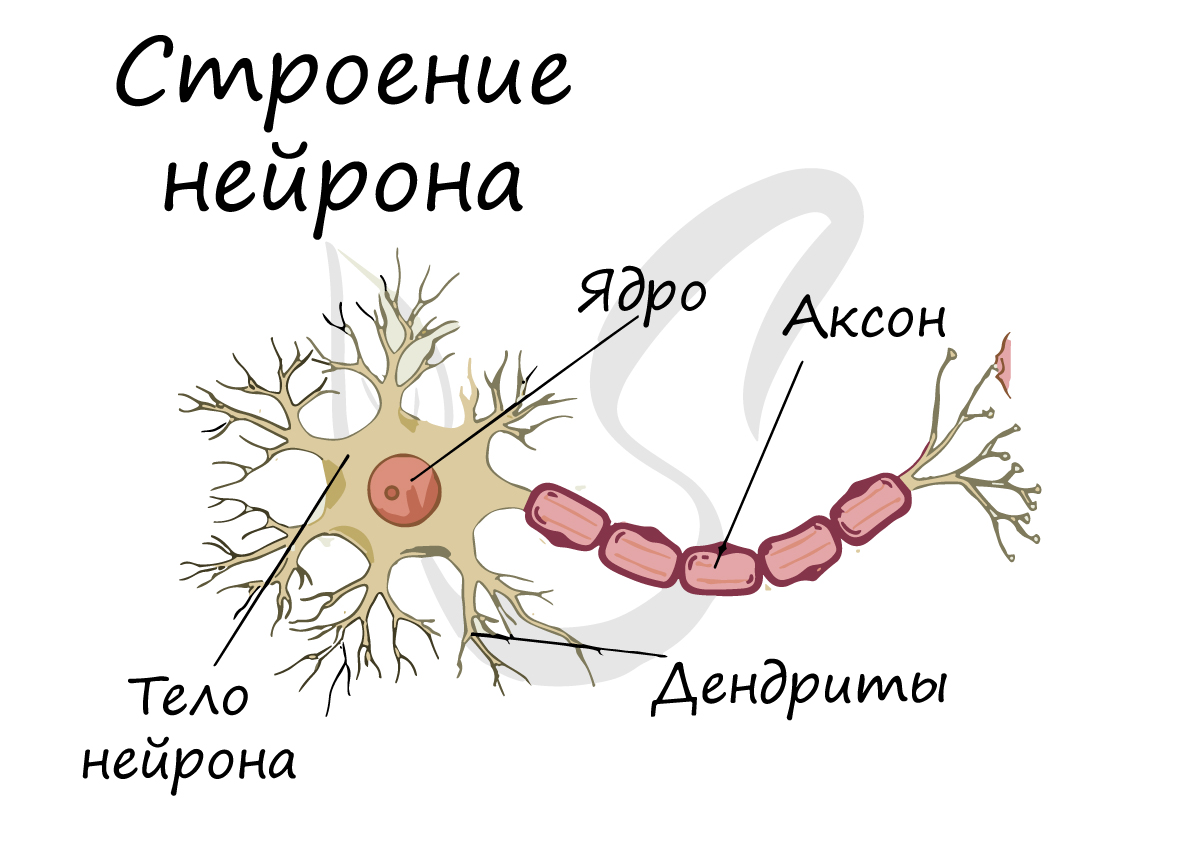
Спешу сообщить, что представление, будто короткий отросток нейрона - дендрит, а длинный - аксон, в корне неверно. С точки зрения физиологии правильнее дать следующие определения: дендрит - отросток нейрона, по которому нервный импульс перемещается к телу нейрона, аксон - отросток нейрона, по которому импульс перемещается от тела нейрона.
Отростки нейронов проводят сгенерированные нервные импульсы и передают их другим нейронам, эффекторам (мышцы, железы), благодаря чему мышцы сокращаются или расслабляются, а секреция желез усиливается или уменьшается.
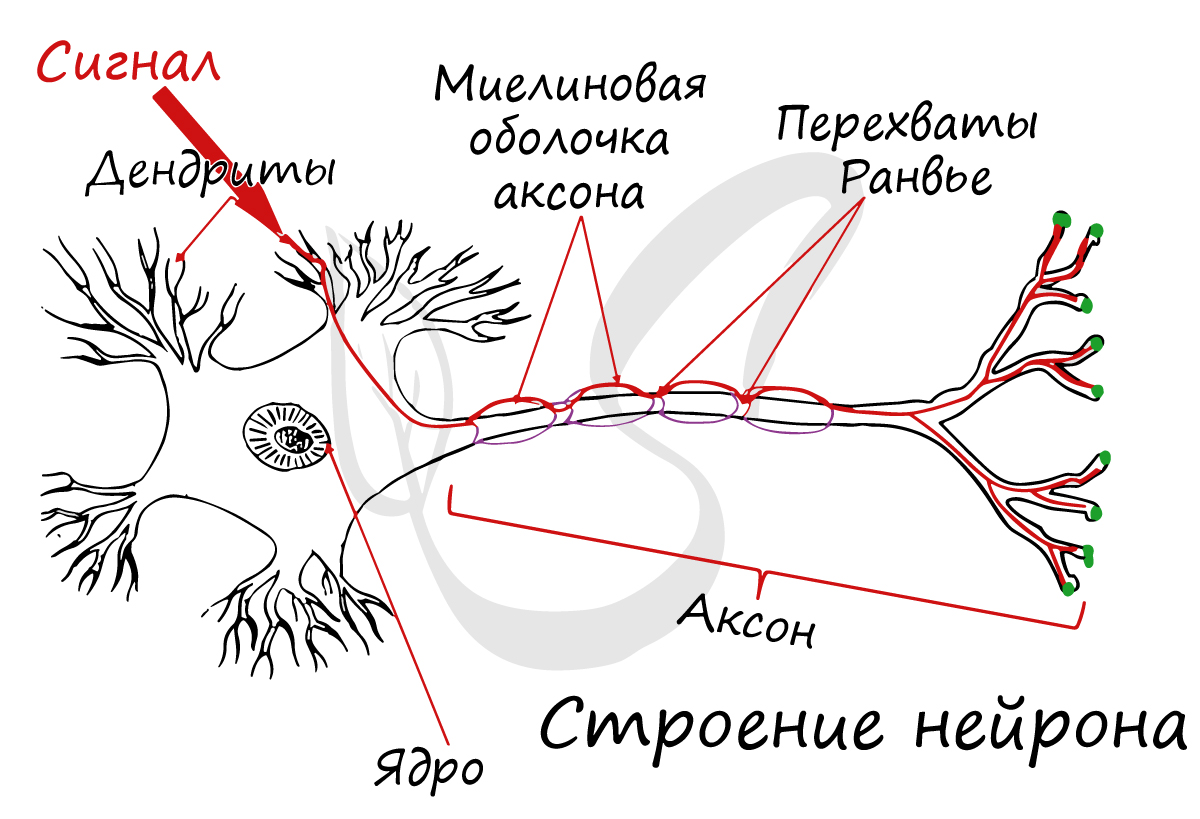
Отростки нейронов покрыты жироподобным веществом - миелиновой оболочкой, которая обеспечивает изолированное проведение нервного импульса по нерву. Если бы не было миелиновой оболочки (вообразите!) нервные импульсы распространялись бы хаотично, и, когда мы хотели сделать движение рукой, двигалась бы нога.
Существует болезнь, при которой собственные антитела уничтожают миелиновую оболочку (случаются и такие сбои в работе организма.) Эта болезнь - рассеянный склероз, по мере прогрессирования приводит к разрушению не только миелиновой оболочки, но и нервов - а значит, происходит атрофия мышц и человек постепенно становится обездвиженным.

Вы уже убедились, насколько значимы нейроны, их высокая специализация приводит к возникновению особого окружения - нейроглии. Нейроглия - вспомогательная часть нервной системы, которая выполняет ряд важных функций:
- Опорная - поддерживает нейроны в определенном положении
- Изолирующая - ограничивает нейроны от соприкосновения с внутренней средой организма
- Регенераторная - в случае повреждения нервных структур нейроглия способствует регенерации
- Трофическая - с помощью нейроглии осуществляется питание нейронов: напрямую с кровью нейроны не контактируют
В состав нейроглии входят разные клетки, их в десятки раз больше чем самих нейронов. В периферическом отделе нервной системы миелиновая оболочка, изученная нами, образуется именно из нейроглии - шванновских клеток. Между ними хорошо заметны перехваты Ранвье - участки, лишенные миелиновой оболочки, между двумя смежными шванновскими клетками.
Нейроны функционально подразделяются на чувствительные, двигательные и вставочные.
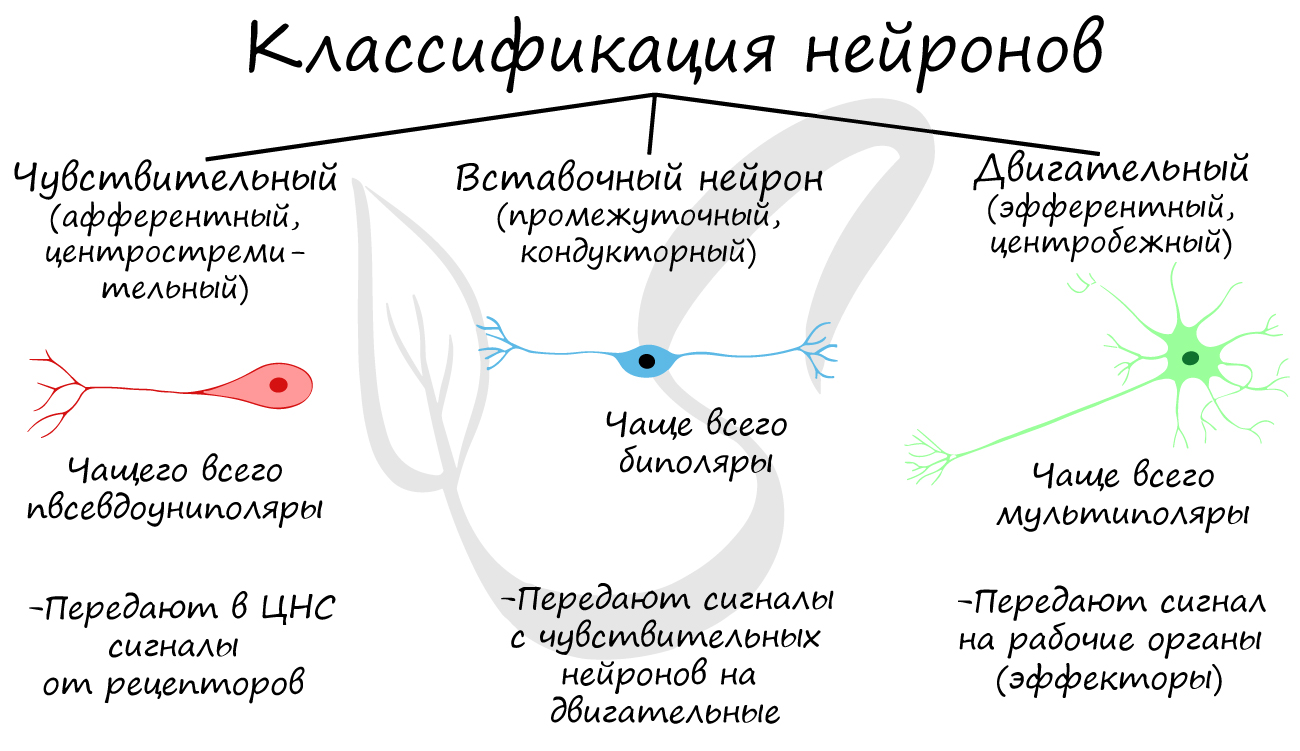
Чувствительные нейроны также называются афферентные, центростремительные, сенсорные, воспринимающие - они передают возбуждение (нервный импульс) от рецепторов в ЦНС. Рецептором называют концевое окончание чувствительных нервных волокон, воспринимающих раздражитель.
Вставочные нейроны также называются промежуточные, ассоциативные - они обеспечивают связь между чувствительными и двигательными нейронами, передают возбуждение в различные отделы ЦНС.
Двигательные нейроны по-другому называются эфферентные, центробежные, мотонейроны - они передают нервный импульс (возбуждение) из ЦНС на эффектор (рабочий орган). Наиболее простой пример взаимодействия нейронов - коленный рефлекс (однако вставочного нейрона на данной схеме нет). Более подробно рефлекторные дуги и их виды мы изучим в разделе, посвященном нервной системе.
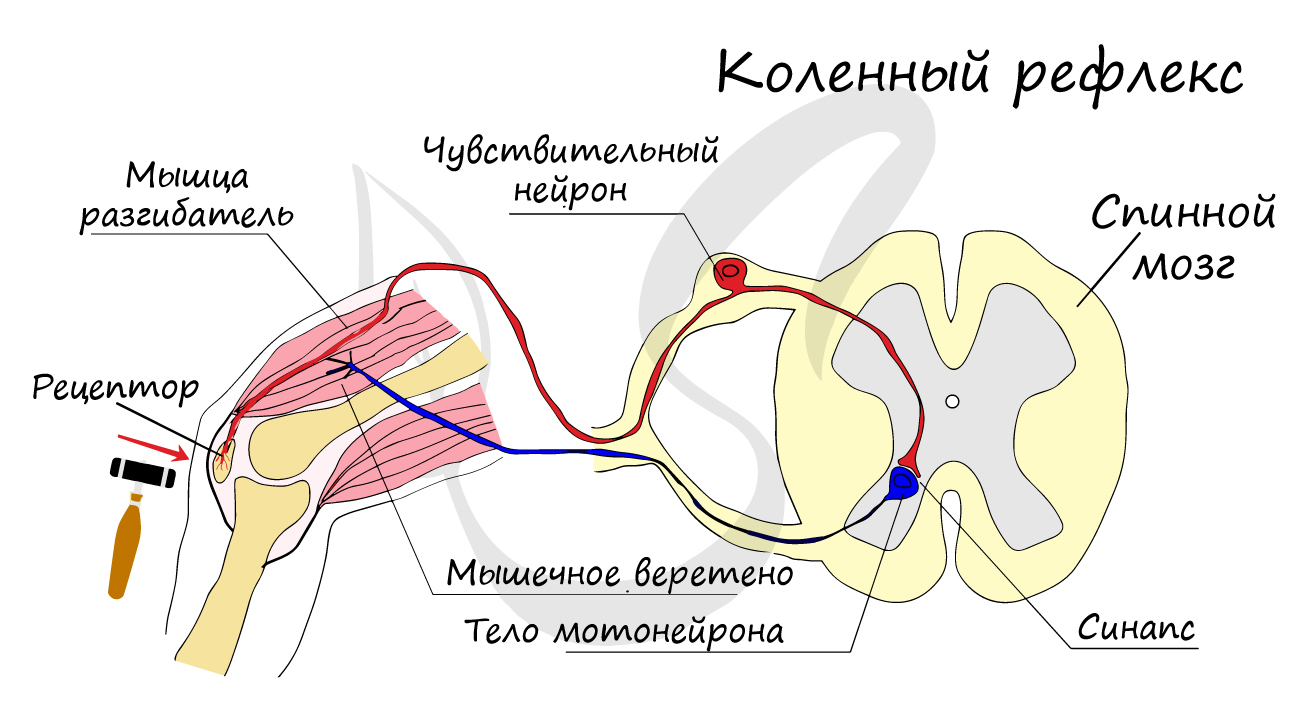
На схеме выше вы наверняка заметили новый термин - синапс. Синапсом называют место контакта между двумя нейронами или между нейроном и эффектором (органом-мишенью). В синапсе нервный импульс "преобразуется" в химический: происходит выброс особых веществ - нейромедиаторов (наиболее известный - ацетилхолин) в синаптическую щель.
Разберем строение синапса на схеме. Его составляют пресинаптическая мембрана аксона, рядом с которой расположены везикулы (лат. vesicula — пузырек) с нейромедиатором внутри (ацетилхолином). Если нервный импульс достигает терминали (окончания) аксона, то везикулы начинают сливаться с пресинаптической мембраной: ацетилхолин поступает наружу, в синаптическую щель.
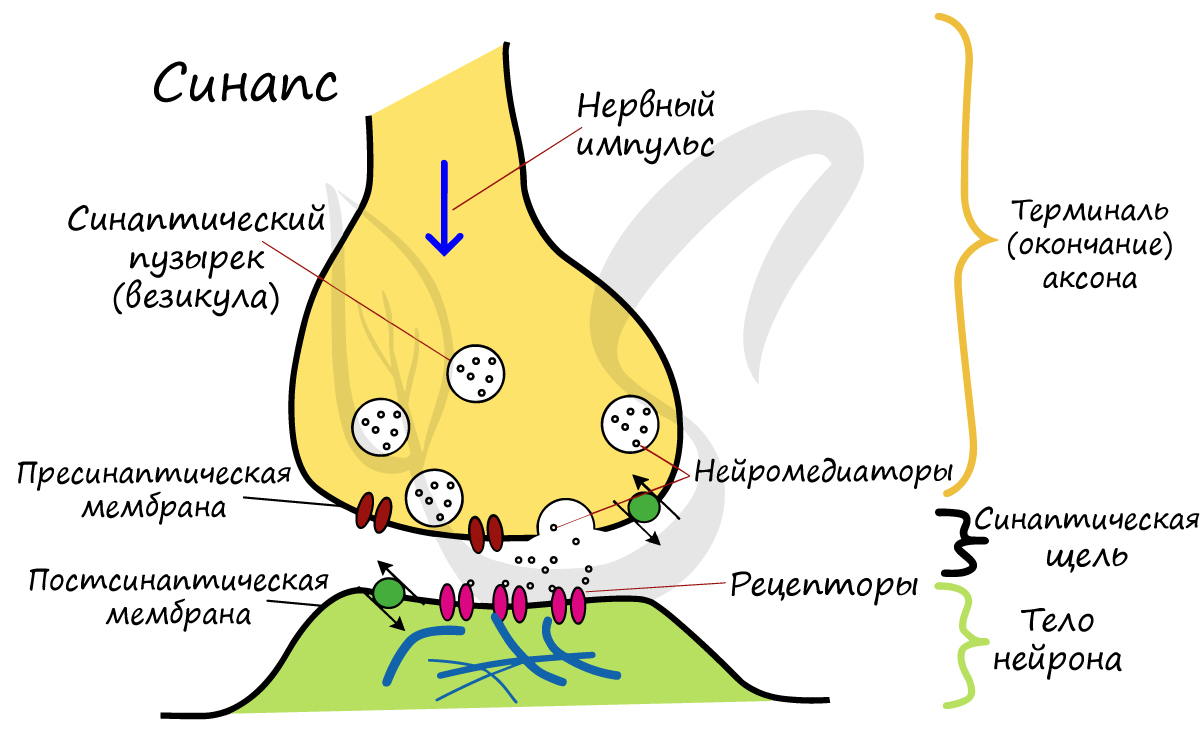
Попав в синаптическую щель, ацетилхолин связывается с рецепторами на постсинаптической мембране, таким образом, возбуждение передается другому нейрону, и он генерирует нервный импульс. Так устроена нервная система: электрический путь передачи сменяется химическим (в синапсе).
Гораздо интереснее изучать любой предмет на примерах, поэтому я постараюсь как можно чаще радовать вас ими ;) Не могу утаить историю о яде кураре, который используют индейцы для охоты с древних времен.
Этот яд блокирует ацетилхолиновые рецепторы на постсинаптической мембране, и, как следствие, химическая передача возбуждения с одного нейрона на другой становится невозможна. Это приводит к тому, что нервные импульсы перестают поступать к мышцам организма, в том числе к дыхательным мышцам (межреберным, диафрагме), вследствие чего дыхание останавливается и наступает смерть животного.

Собираясь вместе, аксоны образуют нервные пучки. Нервные пучки объединяются в нервы, покрытые соединительнотканной оболочкой. В случае, если тела нервных клеток концентрируются в одном месте за пределами центральной нервной системы, их скопления называют нервные узлы - или ганглии (от др.-греч. γάγγλιον — узел).
В случае сложных соединений между нервными волокнами говорят о нервных сплетениях. Одно из наиболее известных - плечевое сплетение.
Неврологические болезни могут развиваться в любой точке нервной системы: от этого будет зависеть клиническая картина. В случае повреждения чувствительного пути пациент перестает чувствовать боль, холод, тепло и другие раздражители в зоне иннервации пораженного нерва, при этом движения сохранены в полном объеме.
Если повреждено двигательное звено, движение в пораженной конечности будет невозможно: возникает паралич, но чувствительность может сохраняться.

Постепенно любые движения мышцами становятся для пациента все труднее, становится тяжело долго говорить, повышается утомляемость. Наблюдается характерный симптом - опущение верхнего века. Болезнь может привести к слабости диафрагмы и дыхательных мышц, вследствие чего дыхание становится невозможным.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также:
