
Как распространяется возбуждение по нервной клетке
В состояние покоя любая живая (нервная, мышечная, железистая, нервное волокно) клетка изнутри заряжена отрицательно, а снаружи положительно. Чтобы в этом убедиться подводят два электрода, к внешней мембране клетки, между ними измерительный прибор, который регистрирует ноль на экране – нет разницы потенциалов между двумя точками на мембране. Затем один электрод вводим внутрь, и касаемся внутренней поверхности мембраны, в результате получаем резкий скачок на экране прибора, показатели которого зависят от клетки, с которой проводится опыт, от –30 до -90 мВ. Вывод: есть разность потенциалов между наружной и внутренней поверхностями мембраны.
Так заложено природой, что внутри клетки (с внутренней стороны мембраны) избыток ионов калия, концентрация ионов калия всегда больше в клетке, чем в окружающей среде. Каким же образом создается разница в потенциалах? Внутри калия много, снаружи калия мало, мембрана в состоянии покоя проницаема для ионов калия. Калий будет выходить из клетки, по законам диффузии. Энергия заложена в разнице концентраций, это происходит без затрат энергии. Калий (внутри он заряжен положительно) выходит из клетки, и выносит с собой положительный заряд, и теперь этот положительный заряд не выпускает калий. Создается равновесность потенциалов по калию, т. е. уравновешиваются две силы:
Первая - это диффузия, которая выталкивает калий из клетки,
Вторая – это электростатическая сила, не выпускает калий из клетки.
В каждой клетки свой равновесный потенциал.
Что можно очень легко изменить, чтобы мембранный потенциал стал меняться? - концентрацию калия в окружающей среде. Если мы увеличиваем концентрацию калия в окружающей среде, из клетки будет меньше выходить калия. Если уравняем концентрацию калия в клетке и в окружающей среде, клетка будет мертвой и ни к чему не способной.
При возбуждении:
Сначала меняется проницаемость мембраны для калия, и для натрия. Для калия проницаемость резко понижается, а для натрия резко возрастает. Если мы будем раздражать мембрану, то повышается проницаемость для натрия, натрий начинает входить в клетку. Теперь мембрана дэполяризуется, внутри у нас будет плюс, а снаружи минус, причем не до нуля.
После ответа на один стимул, мембрана должна вернуться в состояние покоя, чтобы ответить на другой стимул. Для чего нам нужен фермент, который разрушает медиатор? Медиатор возбуждает мембрану постсинаптическую, он должен быть разрушен для восстановления в клетке состояния покоя. Состояние покоя восстанавливается через процесс реполяризации. Этот процесс возникает потому, что инактивируются натриевые каналы. Сначала они закрыты, потом они открыты, а на пике деполяризации они инактивируются, т.е. они не могут дальше открываться. В этот момент открывается канал для калия, калий выходит из клетки и восстанавливается исходный мембранный потенциал. Количество каналов подсчитано, структура их определена за счет блокаторов: -калиевые каналы блокируются тетроэтиламонием, в результате не будет процесса рэполяризации;
-натриевые каналы блокируются – тетродотоксином, в результате не возникает потенциал действия при раздражении мембраны.
При раздражении возникает потенциал действия, который обычно называем или возбуждение, или сигнал, или биопотенциал. При слабом раздражении может открыться недостаточное количество каналов для натрия, т.е. есть критический уровень деполяризации (КУД) или порог. Например на мышцу наносим стимул, а мышца не сокращается, там происходит локальная деполяризация, которая не приводит к сокращению мышцы, локальный процесс дэполяризации не достигает КУД.
Если натрий в состоянии возбуждения входит в клетку, в итоге в клетке повысится концентрация натрия. Если калий уходит из клетки, в итоге в клетке понизится концентрация калия. У нас нарушится та исходная разность концентраций, которая обеспечивает и потенциал покоя и потенциал действия. Калиево-натриевый насос – это белковая молекула, их задача перекачивать калий в клетку, а натрий из клетки, т. е. он работает против концентрационного грэдиента. В этом случае необходима энергия, АТФ. Если мы энергию заблокируем, то насосы работать не будут. Постепенно будет накапливаться в наружной среде калий, во внутренней – натрий, и мышечное волокно постепенно перестает работать.
По отношению к нервной клетке.
Как происходит передача возбуждения при осуществлении движения. Мотонейрон, его аксон выходит из передних рогов, подходит к скелетной мышце. Каждое отдельное нервное волокно иннервирует отдельное мышечное волокно, к мышце подходит целый нерв, который ее иннервирует. Аксонная терминаль, везикулы, в которых содержится ацетилхолин. Именно работая с мотонейронами, Дейл сформулировал свой принцип:
Доказательство ниже. Рисуем: мотонейрон, миэлиновая оболочка, аксонная терминаль. Доказано, что в аксонной терминале выделяется ацетилхолин. Действительно это медиатор? Нужно было найти фермент, который синтезирует ацетилхолин – нашли, и нашли фермент, который разрушает ацетилхолин - это холинэкстераза. Ацетилхолин выделился, провзаимодействовал с Н-холинорецепторами, и дальше происходит возбуждение мышечного волокна. Окончание мотонейрона, мышечное волокно. Постсинаптическая мембрана в невромышечном синаптическом контакте имеет сложное строение для увеличения площади. В постсинаптической мембране содержатся Н-холинорецепторы, ацетилхолин взаимодействует с Н-холинорецепторами в результате чего мышечное волокно обязательно сокращается.
В постсинаптической мембране возникают процессы, результат мы видим – мышечное волокно сократилось. Для того, чтобы Мышечное волокно сократилось в нем тоже должен возникнуть потенциал действия. Вначале возникают потенциалы концевой пластинки ПКП – это маленькие деполяризации. Выделяется медиатор, сначала возникает один ПКП, другой ПКП … Они суммируются и обязательно достигают критического уровня деполяризации, и обязательно возникает потенциал действия.
Цепочка: … потенциал действия в нервном волокне, выделяется медиатор, взаимодействует с Н-холинорецепторами, возникает потенциал в концевой пластинке, они суммируются между собой потенциалы концевой пластинки, все вместе достигают критического уровня деполяризации, в ответ на это возникает потенциал действия в мышечной мембране, мышечное волокно сокращается.
Если потенциал действия не распространяется по аксону мотонейрона. В этом случае выделяются квантомедиаторы, т. е выделяется чуть-чуть ацетилхолина. Никогда не бывает, чтобы мотонейрон был абсолютно заторможен. Даже при самом большом расслабления мышцы чуть-чуть сокращены, есть такой тонус мускулатуры. Когда у нас потенциал действия не приходит, в постсинаптической мембране возникает миниатюрные потенциалы – это маленькая деполяризация, которая не вызывает потенциал действия. Если ацетилхолин выделился, деполяризация все равно должна возникнуть, но эти миниатюрные потенциалы, даже если суммируются, они не могут вызвать такую деполяризацию, чтобы у нас лавинообразно натрий пошел в клетку.
Это то, что происходит на постсинаптической мембране мышечного волокна, а что в нервной клетке происходит?
Нервная клетка, сюда приходит по аксону потенциал действия, после чего выделился медиатор. Если пришел сигнал только по одному, двум, трем аксонам к одной нервной клетке, с ней ничего не произойдет. Потому, что на теле одного аксона заканчиваются до тысячи окончаний других нервных клеток. Мембрана дэполяризуется, но деполяризация возникает и угасает. Такая деполяризация называется ВПСП – возбуждающий постсинаптический потенциал.
В нервной клетке, в нервном волокне, мышечном волокне есть такое явление как рефрактерность, т. е. если нервное волокно возбудилось, то при повторном сигнале оно уже не возбудимо, т. е. натриевые каналы больше уже раскрываться не могут. Самый большой рефрактэрный период был обнаружен у сердечной мышцы. Сердце сокращается затем расслабляется, естественно находится в состоянии невозбудимости, которое необходимо, чтобы закончилась одна систула, после этого миокарт должен прийти в такое состояние, чтобы снова возбуждаться. Для сердца нужны ритмичные сокращение – расслабление, поэтому у сердца самый большой рефрактэрный период. Меньше рефрактерный период у скелетных мышц. Самый маленький рефрактэрный период у нервных волокон, которое возбуждаются с частотой 400 импульсов в сек.
Есть период рефрактерности т. е невозбудимости. Например, придет важный сигнал, а клетка в состоянии рефрактэрности. Поэтому не любой сигнал вызывает возбуждение нервной клетки. Если к большому числу возникают стимулы, в результате процесса суммации деполяризация достигает критического уровня и в клетке возникает потенциал действия.
Суммация может быть пространственной, а может быть временной.
Пространственная возникает тогда когда ВПСП возникает на большей части мембраны нервного волокна.
Временная возникает при повышении частоты раздражающих стимулов, когда следующий стимул приходит, а предыдущие еще не успели угаснуть.
Рефрактэрный период начинается с точки ____. Потенциал действия – это процесс возбуждения, процесс дэполяризации, и ВПСП – это тоже процесс деполяризации.
Разница между потенциалом действия и ВПСП:
- потенциал действия возникает по закону все или ничего, т е если раздражающий стимул достиг порога то потэнциал обязательно возникает и уже не меняется .
- ВПСП – градуальный процесс, он зависит от количества медиаторов.
- Задача потенциала действия, если он возник дальше распространяться.
- ВПСП – это локальный процесс.
- ВПСП, ПКП суммируются
- Потенциалы действия суммироваться не могут.
У мотонейрона, пока миэлиновая оболочка не началась, где-то около сомы, как правило, есть веточка – коллотераль, из нее тоже выделяется ацетилхолин. Отсюда ДЕЙЛ сделал вывод, что в каждом нейроне вырабатывается только один медиатор. В ЦНС существуют медиаторные системы, в каждой из которых вырабатывается свой медиатор, если вырабатывается ацетилхолин – холинэргическая и т. д.
ГАМэрбическая система – тормозная система в мозге. Что происходит при торможении? Если выделяется тормозный медиатор, например гамаминомаслянная кислота приводит к тому что в клетку начинает входить хлор. В результате увеличится разность потенциалов между внутренней и внешней поверхностью, возникает гиперполяризация мембраны. Возникает ТПСП – тормозные постсинаптические потенциалы. Они тоже обладают свойством суммации, они также локальны, как и ВПСП. Когда возникает ТПСП, порог возбудимости резко повышается, а возбудимость понижается.
Критика принципа ДЕЙЛА
В гипоталамусе обнаружили нейросекреторные клетки, в которых содержатся нейропептиды: статины и либерины. Они выполняют свою эндокринную функцию, статины тормозят выработку гормонов в гипофизе, либерины – усиливают. Эти же нейропептиды были найдены во многих других нервных клетках ЦНС. Потом оказалось, что нейропептиды синтезируются еще и в кишечнике, и в желудке. Последнее, что подвергло сомнению принцип ДЕЙЛА,
было обнаружено: парасимпатическая нервная систем иннервирует слюнные железы, здесь находится ацетилхолин, который усиливает выработку слюнных желез. Здесь же находится вазоинтеспинальный пептид (ВИП), который сначала был обнаружен в кишечнике у свиньи – его задача расширять сосуды в кишечнике. Потом его обнаружили в окончаниях парасимпатической нервной системы вместе с ацетилхолином. Оказалось, что на самом деле в одной аксонной терминале могут сосуществовать в основном классические медиаторы с нейропептидами. Зачем это сосуществование? Эти нейропептиды являются модуляторами – они видоизменяют эффекты классических медиаторов. Каким образом:
1) Повышают чувствительность пре- и пост- синаптической мембраны к медиатору;
2) Усиливают синтез медиатора в аксонной терминале
3) Ускоряют выброс медиатора из аксонной терминали
4) Удлиняют действие медиатора, замедляют его разрушение
5) Оказывают внесинаптическое действие, выходят за пределы синаптического контакта, например, расширяют кровеносные сосуды как ВИП
Эти модуляторы, обладают признаками медиаторов, но не в полной степени, или рецептора нет, или плохо выделяются из аксонной терминали, или мы чего- то не знаем.
И вот эти вещества, которые сосуществуют с классическими медиаторами, их в настоящее время порядка 50-ти, называются кандидаты в медиаторы.
Что с чем сосуществует:
Ацетилхолин + ВИП + люлиберин (лю-гормон, ускоряет процессы в половых железах)
Норадреналин + соматостатин (нейропептид, который тормозит выработку соматотропного гормона роста)
Серотонин + тиролиберин (Тирелоидная железа, там тироксин вырабатывается) + энкефалин
Гаммааминомаслянная кислота ГАМК + дофамин + серотонин + энкефалин
Дофамин и серотонин – классические медиаторы.
Рисуем последнюю схему:
Аксонная терминаль , классический медиатор , нейропептид , постсинаптическая мембрана , где рецептор к нейропептиду, и рецептор к классическому медиатору. Кроме того, есть рецептор и в пресинаптической мембране, потому, что некоторые медиаторы выходят в синаптическую щель, а потом захватываются обратно, чтобы их сохранить. Нейропептид усиливает скорость прохождения медиатора через пресинаптическую мембрану, усиливает синтез классического медиатора, и повышает чувствительность рецептора в постсинаптической мембране к классическому медиатору.
Возбуждение как ответ на раздражение. Явления, связанные с возбуждением, издавна изучались на изолированном нервно-мышечном препарате лягушки, для получения которого чаще всего из задней лапки вырезают икроножную мышцу вместе с подходящим к ней седалищным нервом. При раздражении нерва в нем возникает возбуждение. Оно волной пробегает по нервным волокнам, переходит на мышцу и вызывает ее сокращение, которое легко зарегистрировать на специальном приборе—кимографе. Волна возбуждения, или импульс, распространяется по нервам с различной скоростью: в двигательных нервах—до 120 м/сек, а в симпатических — всего лишь несколько метров в секунду. Обнаружить возбуждение можно не только по сокращению мышцы, но и по тем изменениям, которые происходят в самом нерве. Первый, и притом обязательный, признак возбуждения, где бы оно ни возникло,— электрическая реакция.
Ритмический характер возбуждения. Одиночную волну возбуждения легко получить в искусственных условиях опыта. В естественных условиях, как правило, каждое, даже кратковременное раздражение рецепторов вызывает не одну волну, а ряд волн, следующих друг за другом с определенной частотой. Иными словами,
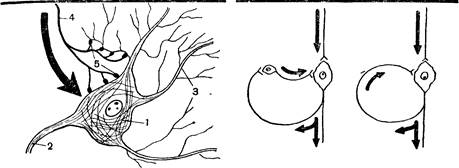
Рис. 28. Схема синапсов:
/—тело нервной клетки; 2—ее аксон;
3 — ее дендриты; 4 — аксон другой нервной клетки; 5 — синапсы.
Рис. 29. Кольцевая связь между нейронами.
возбуждение носит ритмический характер. Ритмическое возбуждение можно получить и в опыте на нервно-мышечном препарате лягушки. В качестве раздражителя обычно применяют электрический ток. Возбуждение возникает при каждом включении и выключении тока, а также при изменении его направления. Для ритмического раздражения пользуются прерывистым постоянным током или индукционным током. Новая волна возбуждения может возникнуть лишь по прекращении предыдущей волны. В двигательных нервных волокнах человека волна возбуждения длится около 0,001 доли секунды. Поэтому за одну секунду по нерву могло бы пройти до 1000 волн. Однако в естественных условиях волны возбуждения, или импульсы, проходят по нервам с небольшой частотой — обычно 10—30 импульсов в секунду.
Проведение возбуждения в центральной нервной системе. Аксон, т. е. длинный отросток одного нейрона, разветвляясь, подходит к телу или дендритам другого нейрона, образуя на его поверхности небольшие бляшки, или утолщения. Контакты между нейронами получили название синапсов (рис. 28). Возбуждение передается через синапсы с аксона одного нейрона на дендриты или тело другого нейрона. В передаче возбуждения участвуют химические вещества, образующиеся в окончаниях аксона. К телу и дендритам каждого нейрона подходят аксоны многих других нейронов. В свою очередь аксон образует ветви, которые подходят к разным нейронам, часто расположенным далеко друг от друга.
Многие группы нервных клеток, находящиеся в различных частях центральной нервной системы, связаны между собой двусторонне: возбуждение, возникшее в одной из них, передается в другую. Особое значение имеет своеобразная кольцевая связь: по ответвлению аксона импульс непосредственно или через промежуточные нейроны возвращается к той же самой нервной клетке (рис. 29).
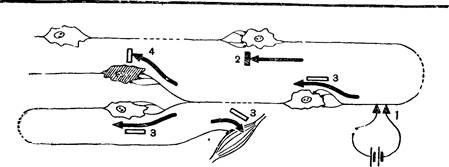
Рис. 30. Схема прохождения импульсов с одного нейрона на другой:
/—раздражаемый участок нерва; 3 — возбуждение не переходит с тела клетки на дендрит;
3 — переход возбуждения на следующий нейрон или на мышцу; 4 — торможение.
(аксон) — от тела клетки в спинной мозг. В спинном мозгу этот отросток разветвляется: одна ветвь идет по белому веществу в нижележащие части спинного мозга, а другая направляется вверх. Обе чти ветви дают боковые веточки, которые вступают в серое вещество и здесь оканчиваются. Эфферентные нейроны имеют один длинный отросток (аксон) и -несколько коротких (дендритов). Тело нейрона находится в передних выступах, или рогах, серого вещества. Отсюда длинный отросток через передний корешок, а затем в составе спинномозгового нерва доходит до мышцы.
В двухнейронной дуге веточки аксона афферентного нейрона, подойдя к передним рогам серого вещества, соприкасаются с эфферентным нейроном. В трехнейронной дуге имеется еще один нейрон:
он называется промежуточным или вставочлым. Однако в подавляющем большинстве случаев возбуждение проходит через большое количество нейронов к различным отделам мозга.
Такая кольцевая связь может поддерживать рабочее состояние нервной клетки: в ней возникают все новые и новые импульсы.
Торможение нервных- клеток. Импульсы, поступающие в мозг, могли бы через многочисленные промежуточные нейроны распространиться по всем его отделам и вызвать общее возбуждение организма. В нормальных условиях импульсы проходят лишь по некоторым из множества возможных путей. Это объясняется возникновением в нервных клетках состояния торможения, при котором они временно теряют способность возбуждаться, а тем самым передавать импульсы другим клеткам. Торможение может возникать то в одних, то в других нейронах. В зависимости от того, какие нейроны в данный момент находятся в состоянии торможения, импульсы пройдут по тому или иному, но всегда определенному пути (рис. 30). Вот почему на одно и то же раздражение ответные реакции могут быть весьма различными.
Рефлекторные дуги. Простейшие рефлекторные дуги состоят из двух или тре-х нейронов (цв. табл. IX). Примером двухнейронной дуги может служить дуга коленного рефлекса. Если человека посадить на стул, предложив ему закинуть одну ногу на другую, а затем ударить ребром ладони или лучше легким молоточком по сухожилию ниже коленной чашечки, то нога подскакивает. Сухожилие, подвергшееся удару, прогибается и тянет за собой мышцу, разгибающую ногу в колене. Мышца растягивается, что вызывает раздражение находящихся в ней рецепторов. Возникающий при этом поток импульсов по афферентным нейронам доходит до спинного мозга, а оттуда по эфферентным возвращается к той же самой мышце, вызывая ее ответное укорочение (цв. табл. IX).
Тела афферентных нейронов находятся в заднем корешке спинномозгового нерва. Они имеют два длинных отростка: один проводит импульсы с рецепторов мышцы к телу клетки, а другой
Последнее изменение этой страницы: 2016-12-27; Нарушение авторского права страницы
Нейроны обеспечивают передачу информации благодаря присущему им свойству проводимости — способности возбуждения распространяться от места возникновения (рис. 2.12) по всей длине своих отростков, а затем и передавать его на другие нейроны, т.е. последовательно возбуждать соседние участки и распространять волну деполяризации.
Все нейроны разделяются на три основных типа, соответствующих их основному месту в ЦНС и функции по передаче информации:
- • афферентные, обеспечивающие передачу нервных импульсов от периферии к центру;
- • эфферентные, обеспечивающие передачу от центров к периферии;
- • промежуточные (вставочные), осуществляющие связь между афферентными, эфферентными, между одноименными и др.; в нервной системы они функционируют не изолированно, а включаясь в линейные нервные цепи (пути) и сети со сложной паутиной взаимосвязей в них.

Рис. 2.12. Схема распространения возбуждения по нейрону (по К. Вилли и В. Детъе):
А — состояние физиологического покоя; Б, В, Г — последовательные стадии распространения возбуждения
- 1) индукционным влиянием электрических полей возбужденных нервных клеток на соседние;
- 2) передачей через особые соединения — синапсы 1 .
Первый механизм обнаруживает свое влияние на небольших участках нервной системы и не является основным [1] [2] .
Синапсы играют важнейшую роль в соединении отдельных нейронов в цепи и сети в передаче нервных процессов по ним. Это место контакта нейронов, специализированная структура на теле или отростке нейрона, обеспечивающая передачу информации (возбуждения) с одного нейрона на другой (рис. 2.13).

Рис. 2.13. Синапс
К особенностям физиологической динамики в нервных путях и сетях, определяемых активной и обильной работой множества нейронов и синапсов и их взаимодействием, относятся (рис. 2.14):
- • одностороннее проведение возбуждения;
- • синаптическая задержка — время поведения возбуждения через синапс в тысячи раз медленнее, чем в нервном волокне, т.е. характеризуется замедлением, что в основном и сказывается на латентном периоде реакции — хорошо известном психологам и другим специалистам промежутке времени между действием раздражителя и ответной реакцией на него;
- • облегчение — каждый последующий импульс проводится быстрее предыдущего;
- • суммация — при действии подпороговых (для постсинаптической мембраны) раздражений возбуждение на следующем нейроне не возникает, однако если на нейрон поступает через разные синапсы несколько подпороговых, то возникновение потенциала действия возможно;
- • окклюзия — при одновременных раздражениях двух сходящихся нейронов суммарная реакция воспринимающего их нейрона (центра) может быть меньше арифметической суммы раздражения каждого из них;
- • трансформация ритма возбуждения (перекодирование) выражается в изменении рисунка импульсов (частоты импульсации, длительности межимпульсных интервалов и др.), что является результатом интегративных процессов в синапсах и других нервных образованиях, суммирующих в себе разные процессы; настройка группы нейронов и центров на один ритм <усвоение ритманейронной цепью, центром) имеет большое значение для взаимодействия между ними;
- • утомляемость — снижение проводимости вплоть до отключения в результате истощения запаса медиатора в клетках;
- • последействие — активность в нервных путях после прохождения возбуждения мгновенно не прекращается и сохраняется определенное время — от нескольких секунд до нескольких лет (например, непрерывная кольцевая циркуляция в части нервных образований), и продолжительность последействия зависит от силы раздражения, его продолжительности, функционального состояния нейронов, цепей и сетей, причем изменения сохраняются не только в виде продолжающихся нервных процессов, но и изменений в синаптических мембранах, тоническом повышении чувствительности и др.

Рис. 2.14. Схемы некоторых видов динамики возбуждения в нервных цепях и сетях
Важным фундаментальным свойством нервных клеток, синапсов и нервных образований является способность не только к возбуждению, но и к торможению. Явление центрального торможения было открыто в 1862 г. И.М. Сеченовым. Он накладывал кристаллики соли на участки мозга препарированной лягушки, и это временно вызывало торможение сгибательных реакций ее лапок даже при нанесении на них болевых раздражений. Раздражение определенных нервов сердца может привести к его остановке.
По современным данным торможение нельзя понимать только как утомление нервных клеток или центров либо их перевозбуждение. Теперь установлено, что это своеобразный нервный процесс — продукт встречи двух возбуждений, из которых одно подавляет другое. Выше отмечалось существование нейронов и синапсов, возбуждение которых приводит к выделению тормозящего медиатора. В одних из них он останавливает распространяющееся возбуждение в самом синапсе — это пресинаптическое торможение. Но есть и синапсы, выделяющие медиатор, оказывающий влияние на постсинаптическую мембрану, находящуюся на другом нерве. При этом происходит перекрытие распространения по нему возбуждения или вообще угнетается его возбудимость — это постсинаптическое торможение.
В больших полушариях тормозных синапсов много больше, чем в подкорковых центрах и периферических нервных путях, что подтверждает большие возможности центрального торможения.
Пессимальное торможение может проявляться при высокой длительности нервной активности возбуждающих синапсов из-за истощение в них медиатора, что прекращает синаптическое проведение.
Существует и следовое торможение, возникающее, когда в синапсах, нервных путях и центрах остается медленно исчезающий положительный следовый потенциал, мешающий включению их в новую работу.
В высшей нервной деятельности обнаруживаются и другие виды торможения.
Весь функционал ЦНС и нервной системы человека зависит от того, насколько хорошо нейроны взаимодействуют друг с другом. Только при совместной работе начинают образовываться сигналы, которые передаются железами, мышцами, клетками организма. Запуск и распространение сигналов происходит посредством ионов, генерирующих электрический заряд, проходимый через нейрон.
Общее число таких клеток в головном мозге человека – около 1011, в каждой из которых содержится примерно 10 тыс. синапсов. Если представить, что каждый синапс – это место для хранения информации, то теоретически мозг человека может хранить все данные и знания, которые накоплены человечеством за всю историю его существования.
Физиологические свойства и функции нейронов будут варьироваться в зависимости от того, в какой мозговой структуре они находятся. Объединения нейронов отвечают за регулирование какой-то конкретной функции. Это могут быть самые простые реакции и рефлексы человеческого организма (например, моргание или испуг), а также особо сложный функционал мозговой деятельности.
Особенности строения
Структура включает в себя три основных составляющих:
- Тело. Тело включает в себя нейроплазму, ядро, которое разграничено мембранным веществом. Хромосомы ядра содержат гены, отвечающие за кодировку синтеза белков. Здесь также осуществляется синтез пептидов, которые требуются для обеспечения нормальной работы отростков. Если тело будет повреждено, то в скором времени произойдет и разрушение отростков. При повреждении одного из отростков (при условии сохранения целостности тела) он будет постепенно регенерироваться.
- Дендриты. Образуют дендритное дерево, имеют безграничное число синапсов, сформированных аксонами и дендритами соседних клеток.
- Аксон. Отросток, который, кроме нейронов, не встречается больше ни в одних клетках. Сложно переоценить их значение (например, аксоны ганглиозных клеток ответственны за формирование зрительного нерва).
Классификация нейронов в соответствии с функциональными и морфологическими признаками выглядит следующим образом:
- по числу отростков.
- по типу взаимодействия с другими клетками.
Все нейроны получают грандиозное число электрических импульсов из-за наличия множества синапсов, которые расположены по всей поверхности нейронной структуры. Импульсы также получаются через молекулярные рецепторы ядра. Электрические импульсы передаются разными нейромедиаторами и модуляторами. Поэтому важным функционалом также можно считать способность интеграции полученных сигналов.
Чаще всего сигналы интегрируются и обрабатываются в синапсах, после чего в остальных частях нейронной структуры суммируются постсинаптические потенциалы.
Мозг человека содержит примерно сто миллиардов нейронов. Число будет варьироваться в зависимости от возраста, наличия хронических заболеваний, травм мозговых структур, физической и умственной активности человека.
Передача возбуждения в синапсах
Рассматривая механизм проведения возбуждения в нейронах, мы ознакомились с синапсами – образованиями, возникающими в месте контакта двух нейроцитов. Возбуждения в первой нервной клетке вызывает образование в коллатералях её аксона молекул химических веществ – медиаторов. К ним относятся аминокислоты, ацетилхолин, норадреналин. Выделяясь из пузырьков синоптических окончаний в синоптическою щель, он может влиять как на собственную постсинаптическую мембрану, так и воздействовать на оболочки соседних нейронов.
Молекулы нейромедиаторов служат раздражителем для другой нервной клетки, вызывая в её мембране изменения зарядов – потенциал действия. Таким образом, возбуждение быстро распространяется по нервным волокнам и достигает отделов центральной нервной системы или же поступает в мышцы и железы, вызывая их адекватное действие.
Развитие и рост нейронов
Современные ученые до сих пор дискутируют на тему деления нервных клеток, т.к. единого мнения по этому вопросу в сфере анатомии на данный момент нет. Многие специалисты в этой области уделяют больше внимания свойствам, а не строению нейронов, что является более важным и актуальным вопросом для современной науки.
Наиболее распространенная версия – развитие нейрона происходит из клетки, деление которой прекращается еще до момента выпуска отростков. Сначала развивается аксон, после чего дендриты.
Зависимо от основного функционала, места расположения и степени активности, нервные клетки развиваются по-разному. Их размеры существенно варьируются в зависимости от места расположения и выполняемых функций.
Структуры нервной ткани
Нейроциты окружены особым веществом, которому присущи опорные и защитные свойства. Для него также характерная способность к делению. Это соединение называется нейроглия.
Эта структура находится в тесной связи с нервными клетками. Так как главные функции нейрона – это генерация и проведение нервных импульсов, то глиальные клетки оказываются под воздействием процесса возбуждения и изменяют свои электрические характеристики. Кроме трофической и защитной функций, глия обеспечивает метаболические реакции в нейроцитах и способствует пластичность нервной ткани.
Основные свойства
Нервные клетки выполняют огромное количество функций. Основные свойства нейрона выглядят следующим образом: возбудимость, проводимость, раздражимость, лабильность, торможение, утомляемость, инертность, регенерация.
Раздражимость считается общей функцией всех нейронов, а также остальных клеток организма. Это их способность давать адекватный ответ на всевозможные раздражения с помощью изменений на биохимическом уровне. Подобные трансформации обычно сопровождаются изменениями ионного равновесия, ослаблением поляризации электрических зарядов в зоне воздействия раздражителя.
Несмотря на то, что раздражимость является общей способностью всех клеток человеческого организма, наиболее выражено она проявляется именно у нейронов, которые связаны с восприятием запаха, вкуса, света и иных подобных раздражителей. Именно процессы раздражимости, протекающие в нервных клетках, запускают другую способность нейронов – возбудимость.
Важнейшее физиологическое свойство нервных клеток, которое заключается в генерировании потенциала действия на раздражитель. Под ним подразумеваются различные изменения, происходящие внутри и снаружи организма человека, которые воспринимаются нервной системой, что и приводит к вызову ответной детекторной реакции. Принято различать два вида раздражителей:
- Физические (получение электрических импульсов, механическое воздействие на разные участки тела, изменение окружающей температуры и температуры тела, световое воздействие, наличие или отсутствие света).
- Химические (изменения на биохимическом уровне, которые считываются нервной системой).
При этом наблюдается разная чувствительность нейронов к раздражителю. Она может быть адекватной и не адекватной. Если в организме человека есть структуры и ткани, которые могут воспринимать конкретного раздражителя, то к нему нервные клетки имеют повышенную чувствительность. Подобные раздражители считаются адекватными (электроимпульсы, медиаторы).
Свойство возбудимости актуально только для нервной и мышечной ткани. Также принято считать, что возбудимостью обладает и ткань желез. Если железа активно работает, то могут отмечаться различные биоэлектрические проявления с ее стороны, потому что она включает в себя клетки разных тканей организма.
Соединительная и эпителиальная ткани не обладают свойством возбудимости. Во время их работы не генерируются потенциалы действия даже в том случае, если происходит непосредственное воздействие раздражителя.
Левое полушарие мозга всегда содержит большее количество нейронов, нежели правое. При этом разница совсем незначительная – от нескольких сотен миллионов до нескольких миллиардов.
Разговаривая о том, каковы свойства нейронов, после возбудимости практически всегда отмечают проводимость. Функция проводника у нервной ткани заключается в особенности проведения возникшего в результате воздействия раздражителя возбуждения. В отличие от возбуждения, функцией проводимости наделены все клетки человеческого тела – это общая способность ткани менять тип своей активной деятельности в условиях воздействия раздражителя.
Повышенная проводимость в нейронных структурах наблюдается при развитии доминантного очага возбуждения. В одном нейроне может происходить конвергенция (объединение сигналов множественных входов, которые исходят от одного источника). Подобное актуально для ретикулярной формации и ряда других систем человеческого организма.
При этом клетки, вне зависимости от структур, в которых они располагаются, могут по-разному реагировать на воздействие раздражителя:
- Изменяется выраженность и выполнение процессов по обмену веществ.
- Изменяется уровень проницаемости мембраны клеток.
- Изменяются биоэлектрические проявления нейронов, двигательная активность ионов.
- Ускоряются процессы развития и деления клеток, повышается выраженность структурных и функциональных реакций.
Выраженность этих изменений также может серьезно варьироваться в зависимости от типа раздражителя, ткани и структуры, в которых находятся нейроны.
Часто можно слышать выражение – нужно предотвращать гибель нервных клеток. Но их гибель запрограммировала природа – за один год человек теряет примерно 1% всех своих нейронов, и никак предупредить подобные процессы нельзя.
Под лабильностью нервных клеток подразумевается скорость течения простейших реакций, которые лежат в основе раздражителя. В обычных условиях, при нормальном развитии всех мозговых структур, у человека отмечается максимально возможная скорость течения. Нейроны, которые различаются электрофизиологическими свойствами и размерами, имеют разные значения лабильности за единицу времени.
В одной нервной клетке лабильность различных структур (аксонной и дендритной частей, тела) будет заметно отличаться. Показатели лабильности нервной клетки определяют с помощью степени ее мембранного потенциала.
Показатели мембранного потенциала должны находиться на определенном уровне, чтобы в нейроне могла получиться наиболее подходящая степень возбудимости и лабильности (зачастую вкупе с ритмической активностью). Только в этом случае нервная клетка сможет в полной мере передать полученную информацию в виде электрических импульсов. Подобные процессы и обуславливают работу нервной системы в целом, а также гарантируют нормальное протекание и формирование всех необходимых реакций.
В спинном мозге предельный уровень ритмической активности нервных клеток может достигать значения в 100 импульсов в секунду, что соответствует наиболее оптимальным значениям мембранного потенциала. В обычных условиях данные значения редко превышают уровень в 40-70 импульсов в секунду.
Существенное превышение показателей наблюдается при характерных выраженных реакциях, поступающих со стороны главных отделов ЦНС, мозговых структуры, коры. Частота разрядов при определенных условиях может достигать значений в 250-300 импульсов в секунду, но подобные процессы развиваются крайне редко. Также они являются кратковременными – их быстро сменяют замедленные ритмы активности.
Наиболее высокие показатели частоты разрядов обычно наблюдаются в нервных клетках спинного мозга. В возникающих в результате выраженного воздействия раздражителя очагах начальных реакций частота разрядов может составлять 700-1000 импульсов в секунду. Протекание подобных процессов в нейронных структурах является необходимостью, чтобы клетки спинного мозга могли резко и быстро воздействовать на мотонейроны. Спустя небольшой промежуток времени частота разрядов существенно снижается.
Нейроны существенно различаются по размеру (в зависимости от места расположения и других факторов). Размеры могут варьироваться от 5 до 100 мкм.
С точки зрения физиологии человека торможение, как ни странно, является одним из наиболее активных процессов, протекающих в нейронных структурах. Особенности строения и свойств нейронов подразумевают, что торможение вызывается возбуждением. Процессы торможения проявляются в снижении активности или предупреждении вторичной волны возбуждения.
Способность нервных клеток к торможению совместно с функцией возбуждения позволяет обеспечить нормальную работу отдельных органов, систем, тканей организма, а также всего человеческого тела в целом. Одна из наиболее важных характеристик процессов торможения в нейронах – обеспечение защитной (охранной) функции, что актуально для клеток, располагающихся в коре головного мозга. За счет процессов торможения также обеспечивается защита ЦНС от чрезмерного перевозбуждения. Если они нарушены, у человека проявляются негативные психоэмоциональные черты и отклонения.
Важной функцией торможения также является прямое взаимодействие с возбуждением, что позволяет анализировать и синтезировать в центральной нервной системе полученные электрические импульсы. Это помогает правильно согласовывать деятельность и функции всех систем, тканей и органов человеческого тела, а также адекватно контактировать с окружающей средой. Данную функцию также принято называть координационной.
Несмотря на то, что нейроны имеют удивительно малые размеры, современные технологии позволяют ученым провести измерение активности каждого найденного нейрона. Подобные процедуры зачастую проводятся для диагностики различных заболеваний (например, эпилепсии).
К общим признакам всех нейронов относится их способность к физиологической и репаративной регенерации. В нервных клетках она подразумевает протекание следующих процессов:
- Частичное увеличение количества хромосом в ядре.
- Восстановление синапсов (если они были повреждены).
- Развитие и возвращение в обычное состояние отростков (при их повреждении).
- Обновление метаболических и химических компонентов нервных клеток в процессе протекания внутриклеточного обмена веществ.
Если нервная ткань будет повреждена, то в зоне поражения сразу начнет развиваться нейроглия. Это невыраженная дифференцированная ткань, которая делится митозом.
В случае получения повреждений, которые нарушили целостность нервных волокон, происходит распадение периферических частей на отдельные части миелиновых оболочек и осевых цилиндров. Если отсутствуют воспалительные процессы, рубцы соединительной ткани, то есть высокая вероятность восстановления иннервации нервных тканей. Отростки нейронов регенерируются довольно быстро – 2-3 мм за 24 часа.
Вопреки распространенному мнению, нейроны вполне могут восстанавливаться – их генерирование происходит сразу в трех частях организма человека.
Образование нервной ткани
Прежде чем изучать функции нейрона, давайте разберемся, каким образом формируются клетки-нейроциты. На стадии нейрулы у зародыша закладывается нервная трубка. Она формируется из эктодермального листка, имеющего утолщение – нервной пластинки. Расширенный конец трубки в дальнейшем сформирует пять частей в виде мозговых пузырей. Из них образуются отделы головного мозга. Основная часть нервной трубки в процессе зародышевого развития сформировывает спинной мозг, от которого отходит 31 пара нервов.
Нейроны головного мозга объединяются, образуя ядра. Из них выходит 12 пар черепно-мозговых нервов. В организме человека нервная система дифференцируется на центральный отдел – головной и спинной мозг, состоящий из клеток-нейроцитов, и опорную ткань – нейроглию. Периферический отдел состоит из соматической и вегетативной части. Их нервные окончания иннервируют все органы и ткани организма.
Читайте также:
