
Механизм передачи нервного импульса в адренергических синапсах
ЛЕКАРСТВЕННЫЕ СРЕДСТВА, ВЛИЯЮЩИЕ НА АДРЕНЕРГИЧЕСКИЕ СИНАПСЫ
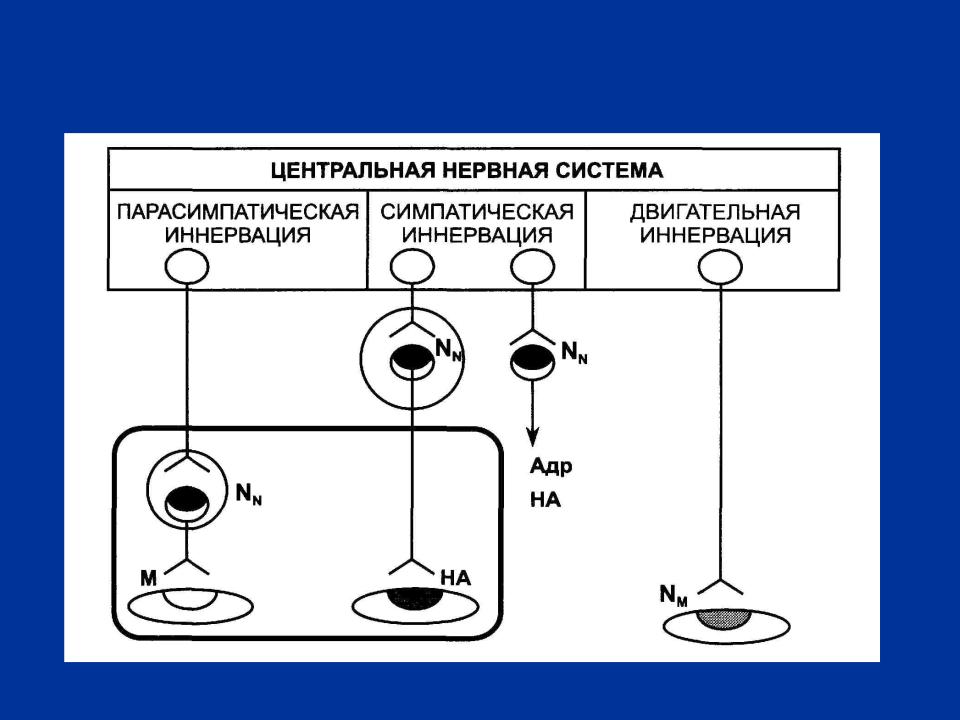
РИС. 1 СХЕМА ЭФФЕРЕНТНОЙ ИННЕРВАЦИИ
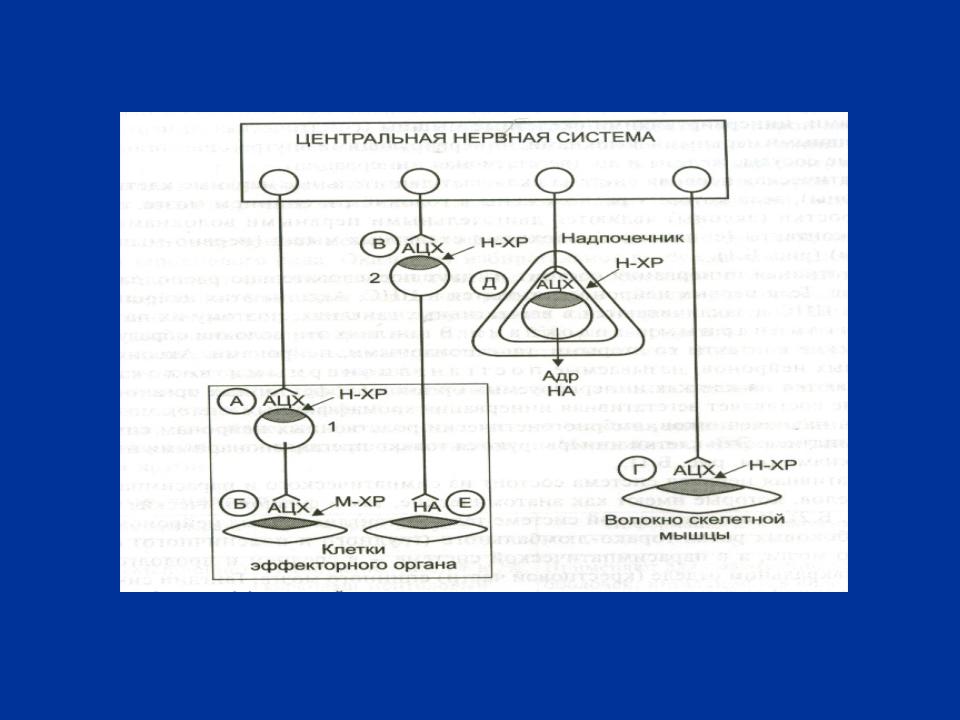
РИС. 2 СХЕМА ЛОКАЛИЗАЦИИ АДРЕНЕРГИЧЕСКИХ СИНАПСОВ
Обозначения: А, Б, В, Г, Д – холинергические синапсы; Е
– адренергический синапс, 1 - ганглий парасимпатической системы, 2 – ганглий симпатической системы

ЛОКАЛИЗАЦИЯ АДРЕНЕРГИЧЕСКИХ СИНАПСОВ
1. Исполнительные органы, получающие постганглионарную симпатическую иннервацию (кроме потовых желез)
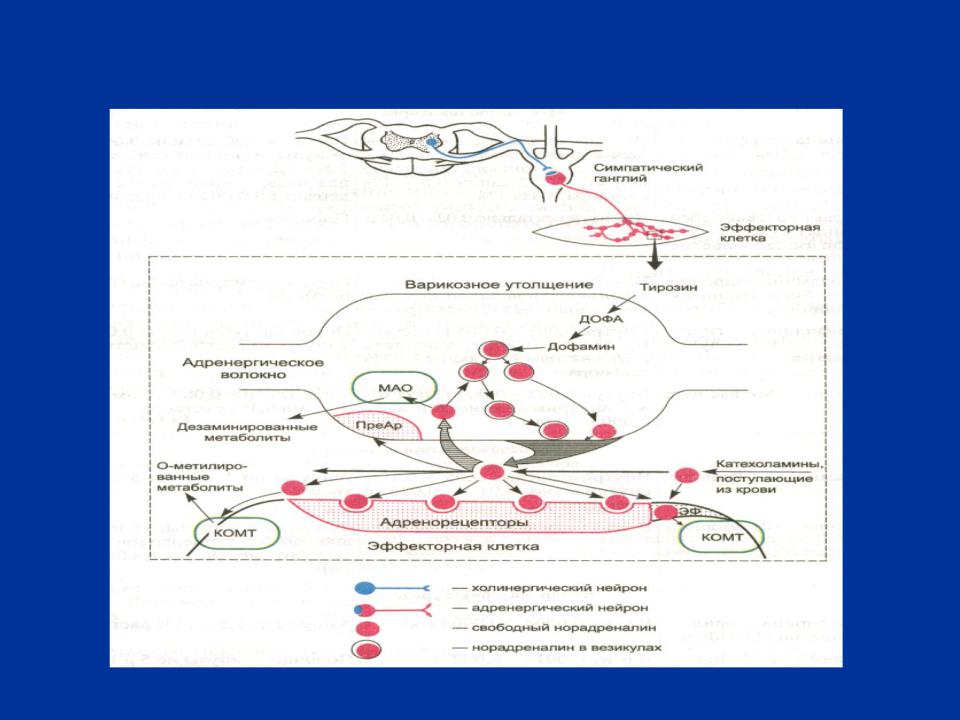
РИС. 3 СИМПАТИЧЕСКИЙ НЕРВ И АДРЕНЕРГИЧЕСКИЙ СИНАПС (схема)

МЕХАНИЗМ ПЕРЕДАЧИ НЕРВНОГО ИМПУЛЬСА В АДРЕНЕРГИЧЕСКОМ СИНАПСЕ
Медиатором в адренергических синапсах является норадреналин (НА).
Процесс синтеза НА начинается в печени, где из аминокислоты фенилаланина образуется аминокислота тирозин. Обе аминокислоты в большом количестве присутствуют в твороге, сыре, шоколаде, бобовых.
Тирозин с помощью активного транспорта поступает в адренергические окончания и в их цитоплазме сначала превращается в ДОФА (реакцию катализирует тирозингидроксилаза), а затем в дофамин (реакцию катализирует декарбоксилаза).
Дофамин далее поступает в синаптические пузырьки (из-за своего строения под электронным микроскопом они получили название гранулы) и превращается в НА (реакцию катализирует дофамин- - гидроксилаза). В гранулах НА депонирован в связи с АТФ и белком хромогранином и находится в виде двух фракций: стабильная (60- 70% НА прочно связанного с белком и не участвует в передаче нервного импульса) и лабильная (30-40% НА непрочно связанного с белком и который выделяется в синаптическую щель при деполяризации пресинаптической мембраны.

МЕХАНИЗМ ПЕРЕДАЧИ НЕРВНОГО ИМПУЛЬСА В АДРЕНЕРГИЧЕСКОМ СИНАПСЕ
Выделившийся в синаптическую щель НА взаимодействует с адренорецепторами как на постсинаптической мембране ( 1
– АР и 1 – АР), так и на пресинаптической мембране ( 2 – АР и 2 – АР).
Возбуждение АР на постсинаптической мембране сопровождается активацией вторичных посредников в эффекторных клетках и изменением функциональной активности органа-исполнителя.
Возбуждение АР на пресинаптической мембране влияет на процесс выделения НА в синаптическую щель (возбуждение 2 – АР приводит к снижению выделения НА, возбуждение 2
– АР приводит к усилению выделения НА).
После взаимодействия НА с АР происходит ряд процессов:
- большая часть НА (70-80%) путем активного транспорта сначала попадает в цитоплазму пресинаптического окончания и затем внутрь гранул;
- часть НА (10-20%) инактивируется ферментами КОМТ в синаптической щели, МАО - в аксоплазме;
- часть НА (10%) подвергается экстранейрональному захвату клетками исполнительных органов.
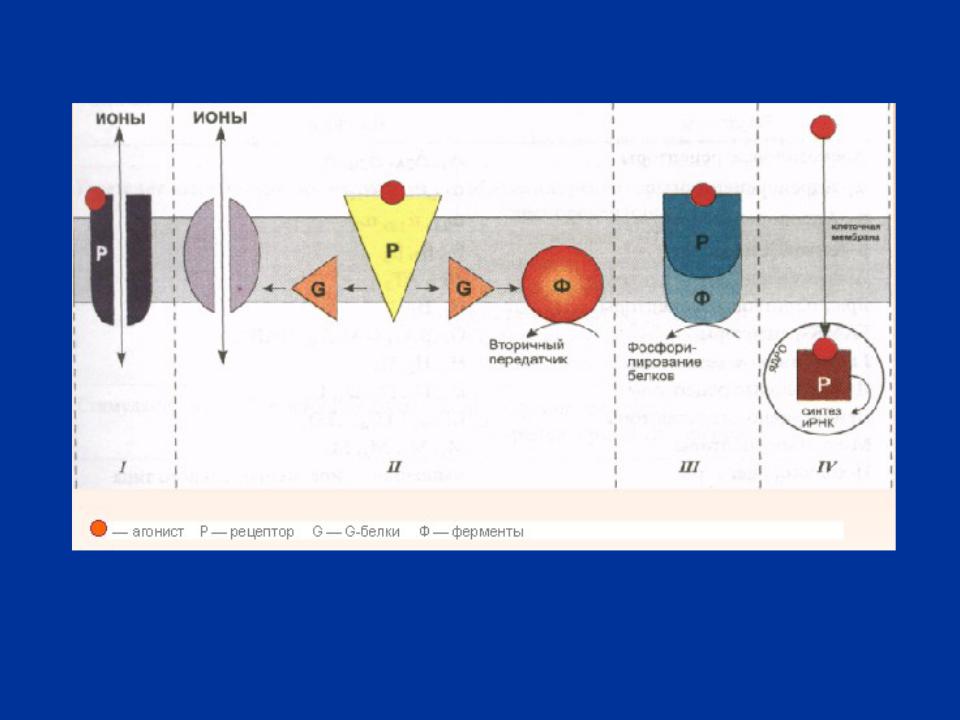
РИС. 4 ПРИНЦИП РАБОТЫ - И -
II – опосредованное влияние (через G-белки) на проницаемость ионных каналов или на активность ферментов, регулирующих образование вторичных передатчиков
Механизм химической передачи нервных импульсов через синапс сводится к тому, что содержимое синаптических пузырьков (медиатор) поступает в виде небольших порций (квантов) в синаптическую щель и взаимодействует затем с рецепторными белками постсинаптической мембраны. Это вызывает деполяризацию мембраны и возбуждение следующего нейрона. Ультраструктурные особенности синапса и механизм передачи импульсов определяют строгую однонаправленность передачи импульсов, что лежит в основе проведения импульсов по рефлекторным дугам.
В зависимости от того, какое вещество выполняет роль нейромедиатора, синапсы подразделяются на холинергические (медиатор — ацетилхолин), адренергические (адреналин и норадреналин), дофаминергические (дофамин), серотонинергические (серотонин), пептидергические (медиаторы — пептиды и аминокислоты, например, мет-энкефалин, гамма-аминомасляная кислота, глицин и др.).
Нейрохимические синапсы подразделяются в функциональном отношении на две противоположные по своему значению группы — возбуждающие и тормозные. Свойства этих синапсов зависят как от медиаторов, так и от ультраструктурных особенностей синапсов. Так, некоторые медиаторы (например, глутамат) характерны для возбуждающих синапсов, а в тормозных синапсах медиатором является гамма-аминомасляная кислота. Предполагают, что в возбуждающих синапсах к постсинаптической мембране прилежит электронно-плотное вещество, в связи с чем синапс приобретает асимметричное строение.
В пресинаптических частях таких соединений содержатся круглые синаптические пузырьки. Тормозные синапсы имеют симметричное строение. Синаптическая щель у них сужена и в пресинаптической части содержатся уплощенные синаптические пузырьки.
Кроме нейрохимических синапсов между нервными клетками (преимущественно между дендритами или телами нейронов) возникают электротонические синапсы. Последние у млекопитающих встречаются редко и по строению соответствуют щелевым контактам. Они проводят возбуждение благодаря формированию трансмембранных каналов — коннексонов.
Каждый нейрон на своей поверхности имеет огромное количество (до 10000) синапсов. Интеграция импульсов в пределах отдельного нейрона происходит так: от синапсов, расположенных на дендритах и теле, по его плазмолемме передается импульс к аксонному холмику (генераторному пункту нейрона), где путем суммирования всех возбуждающих и тормозных импульсов возникает результирующий потенциал действия.
Синаптические структуры обладают высокой чувствительностью к действию токсических факторов, в том числе психотропных отравляющих веществ. Их изменения играют важную роль в механизмах реактивности нервных тканей.
Рецепторные нервные окончания подразделяются на две группы: экстерорецепторы, воспринимающие раздражения из внешней среды, и интерорецепторы, служащие для восприятия раздражений из внутренней среды организма. В зависимости от специфичности раздражения, воспринимаемого рецептором, различают следующие чувствительные нервные окончания: механорецепторы, барорецепторы, хеморецепторы, терморецепторы, болевые рецепторы (ноцицепторы). Все рецепторы по особенностям их строения подразделяют на свободные и несвободные нервные окончания.
Первые из них состоят только из конечных разветвлений периферического отростка чувствительного нейрона; вторые — имеют в составе рецептора кроме нервного отростка глиальный компонент, а нередко и соединительнотканную капсулу. В последнем случае несвободные рецепторные окончания называют инкапсулированными. К числу таких рецепторов относятся пластинчатые, осязательные тельца, концевые колбы, мышечные веретена и др.
Эффекторные нервные окончания подразделяются на двигательные и секреторные. Двигательный, или моторный, эффектор передает нервные импульсы на рабочие органы и ткани. В скелетных мышцах они называются нервно-мышечными (аксомышечными) окончаниями. Терминальное разветвление аксона двигательного нейрона заканчивается булавовидным расширением. Оно соответствует пресинаптической части нервно-мышечного синапса, содержит скопление синаптических пузырьков и митохондрий, ограничено пресинаптической мембраной характерного строения.
Плазмолемма мышечного волокна в этой области образует многочисленные складки и углубления. В постсинаптической части волокна находится зернистая саркоплазма с большим количеством митохондрий и овальных ядер. Синаптическая щель имеет ширину 50-100 нм. У человека медиатором в нервно-мышечных синапсах является ацетилхолин. Моторные окончания в гладкой мышечной ткани, а также секреторные эффекторы имеют вид тонких пучков аксонов или их одиночных терминалей, прилежащих к клеткам иннервируемых тканей.
Возрастные изменения нервных тканей связаны с постепенным уменьшением запаса нервных клеток, особенно — чувствительных нейронов, а также снижением уровня метаболических процессов, что выражается в закономерном накоплении включений липофусцина ("пигмент изнашивания") в нейроплазме.
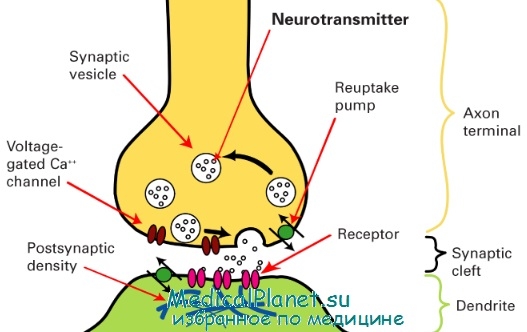
Схема строения синапса
(доц. Данильчук В.В.)
Адренергические синапсы располагаются в органах, получающих постганглионарную симпатическую иннервацию. Синапс представляет собой контакт окончания адренергического нерва с клеткой рабочего органа. Здесь посредством медиатора (нейротрансмиттера) осуществляется передача импульсов с симпатического нерва на орган.
В адренергическом синапсе, как и в холинергическом, различают пресинаптическую мембрану, которая представляет собой поверхность нервного окончания, синаптическую щель и постсинаптическую мембрану - поверхность клеток рабочего органа.
Передача импульса через адренергический синапс осуществляется с помощью медиатора норадреналина. Он синтезируется в аксоплазме симпатических нервных волокон из аминокислоты тирозина, которая превращается вначале в ДОФА, затем в дофамин, а он гидроксилируется в норадреналин. 10-20% медиатора располагается в области пресинаптической мембраны в свободном виде. Его количество регулируется ферментом МАО-азой. Основные запасы медиатора норадреналина (80%) сохраняются в нервных окончаниях в везикулах разных размеров в связанном состоянии с белками, АТФ, АДФ. Прочно связанный медиатор размещается в больших (80-120 нм) везикулах. Их всего 4-5%. В них осуществляется заключительный этап синтеза медиатора. Лабильно связанный медиатор, составляющий основную массу норадреналина, располагается в малых (40-50 нм) везикулах. Они размещены вблизи пресинаптической мембраны. Из них медиатор выделяется в синаптическую щель под влиянием нервного импульса. При этом везикулы подтягиваются к мембране, в ней образуются отверстия, везикулы вскрываются (экзоцитоз) и медиатор попадает в синаптическую щель. В механизме экзоцитоза важную роль играет кальций. Он активирует кальмодулин сократительных нитей, которые подтягивают везикулы к мембране и образуют в мембране отверстия. В процессе экзоцитоза играет роль также специальный белок синексин, который изменяет свойства мембраны. Медиатор, выброшенный в синаптическую щель в большом количестве, реагирует с адренорецепторами (адренореактивными структурами), расположенными в постсинаптической мембране синапсов.
Адренорецепторы представляют собой белковые макромолекулы с рядом активных центров, способных взаимодействовать с медиатором. Взаимодействие медиатора с рецепторами приводит к изменению свойств постсинаптической мембраны, особенно ее проницаемости для ионов, вызывает ток ионов Na из щели в клетку, который, при достижении определенной величины, формирует потенциал действия, что ведет к изменению состояния поляризации мембраны на деполяризацию и проведению импульса. Под влиянием последнего в клетках рабочего органа освобождается специальные нуклеотидсвязывающие белки (G-белки) разных типов. Они активируют ферменты аденилатциклазу, фосфолипазу С, Д (последние освобождают и увеличивают вход Ca ++ в клетку), активируют фосфолипазу А2 (она освобождает арахидоновую кислоту, из которой образуются простагландины, тромбоксаны, лейкотриены), активируют ионные каналы и Na + /Н + обменный механизм. Активация ферментов, увеличение Са ++ в клетке, образование местных гормонов приводит к изменению функции рабочего органа.
Как только импульс прошел через синапс, 80% медиатора возвращается из синаптической щели обратно в везикулы пресинаптической мембраны (механизм обратного захвата). Этот механизм может быть активным, направленным против градиента концентрации. В этом случае он носит название нейронального захвата и осуществляется с помощью специального переносчика, который работает по принципу К+/Na+ -насоса. Перенос медиатора может быть и пассивным (экстранейрональный захват ). Часть медиатора норадреналина (20%) инактивируется в клетках постсинаптической мембраны ферментом КОМТ.
Основная масса адренорецепторов размещена на постсинаптических мембранах адренергических синапсов (т.е. в исполнительных органах). Однако, есть адренорецепторы и в пресинаптических мембранах, т.е. в нервных окончаниях. С их помощью регулируется выброс медиатора в щель. В зависимости от типа рецепторов они увеличивают или уменьшают выброс медиатора в щель, и соответственно этому возбуждают или угнетают функцию органа. Имеются также адренорецепторы, расположенные вне синапсов. Они реагируют с теми катехоламинами, которые циркулируют в крови.
Адренорецепторы неоднородны. Всего их различают 4 типа: a1, a2, b1, b2.Каждый из них может иметь свой подтип, например,a1А, a1Б.
В рабочих органах адренорецепторы размещены неравномерно. Например, в сердце, сосудах, легких, ЖКТ имеются рецепторы нескольких типов, а в селезенке, глазах только одного типа.
Адренорецепторы неодинаково чувствительны к различным фармакологическим агентам. Так, a1-адренорецепторы (a1-АР) возбуждаются норадреналином и адреналином. Много a1АР заложено в периферических сосудах, в сосудах кожи и слизистых оболочек, в венах. Их находят в сердце, бронхах, сфинктерах ЖКТ и мочевого пузыря, в матке, селезенке, печени, почках, радиальной мышце глаза, в семенных пузырьках, семявыносящих протоках, в железах ЖКТ и в ЦНС.
При возбуждении a1-АР функция этих органов усиливается, сосуды суживаются, артериальное давление повышается, мускулатура сокращается, секреция желез усиливается, ЦНС возбуждается, усиливается гликогенолиз и липолиз, но продольная мускулатура ЖКТ расслабляется. a1-АР размещены в основном в постсинаптических мембранах.
Альфа2 адренорецепторы (a2 -АР) размещаются пресинаптически, постсинаптически и внесинаптически.
Пресинаптические a2-АР размещаются по ходу холинергических нервов в ЖКТ, т.е. в пресинаптических мембранах холинергических синапсов. Их возбуждение приводит к задержке (уменьшению) выброса ацетилхолина, что ведет к расслаблению мускулатуры ЖКТ и уменьшению секреции. Пресинаптические a2-АР находятся в жировой ткани (их возбуждение угнетает липолиз), в b-клетках поджелудочной железы (их возбуждение ведет к уменьшению секреции инсулина). Возбуждение пресинаптических a2-АР ЦНС задерживает выброс медиаторов ацетилхолина, серотонина и дофамина в синапсах ЦНС, что ведет к развитию седативного эффекта и аналгезии. Они избирательно возбуждаются клонидином (клофелином).
Постсинаптические a2-АР локализуются в понто-медуллярной области ЦНС, особенно в области сосудодвигательного центра, солитарного тракта и центра вагуса. Их возбуждение клонидином вызывает снижение артериального давления и брадикардию.
Внесинаптические a2-АР находятся в гладкой мускулатуре сосудов (они реагируют на катехоламины крови) и в тромбоцитах (их возбуждение вызывает агрегацию последних).
Бета1 адренорецепторы (b1-АР) локализованы постсинаптически в сердце (при их возбуждении увеличивается сила и частота сокращений миокарда, повышается возбудимость, проводимость, автоматизм, активируется гликогенолиз), в ЖКТ и матке (возбуждение сопроводается расслаблением органов), в жировой ткани (усиливается липолиз), в почках (возбуждение сопровождается увеличением секреции ренина). b1-АР избирательно возбуждаются добутамином.
Бета2 адренорецепторы (b2-АР) размещены в основном постсинаптически и внесинаптически. Их находят во многих органах: сердце, бронхах, сосудах, матке, почках, скелетных мышцах, печени, жировой ткани, ЖКТ, слизистых железах, щитовидной железе. Возбуждение b2-АР ведет к расширению бронхов, расслаблению матки, мускулатуры ЖКТ, к усилению работы сердца, увеличению секреции ренина, инсулина, гормонов, активации гликогенолиза и липолиза, усилению работоспособности скелетных мышц. b2-АР избирательно возбуждаются сальбутамолом и фенотеролом.
Пресинаптические b2-АР размещены в окончаниях симпатических нервов. При их возбуждении увеличивается выброс медиатора в синаптическую щель.
В общем, при возбуждении b-АР функция гладкомышечных органов угнетается, а работа сердца, скелетных мышц резко усиливается, стимулируются все обменные процессы, усиливается гликогенолиз в печени и развивается гипергликемия, усиливается расщепление гликогена в скелетных мышцах и увеличивается образование энергии, накапливается молочная кислота, усиливается липолиз, повышается основной обмен и резко возрастает потребление кислорода. b1и b2–адренорецепторы возбуждаются изадрином и адреналином.
Усиление работы сердца (кардиотоническое действие) и усиление обменных процессов при возбуждении b-АР наступает от включения универсального аденилатциклазного механизма обмена, когда от активации аденилатциклазы усиливается образование цАМФ, активируются фосфолипазы, что ведет к увеличению количества Са ++ в кардиомиоцитах, а также активируется фосфорилаза и процессы гликолиза, что ведет к увеличению образования энергии, необходимой для сокращения кардиомиоцитов.
Все средства, действующие в области адренергических синапсов, делятся на 2 основных группы.
А. Адреномиметики или адреностимуляторы Это вещества, которые возбуждают адренорецепторы. По механизму возбуждающего действия их подразделяют на две группы:
1. Адреномиметики прямого действия. Они непосредственно возбуждают рецепторы.
2. Адреномиметики непрямого действия или симпатомиметики. Они увеличивают концентрацию медиатора в синаптической щели и действуют через него.
Б. Антиадренергические средства. Они уменьшают влияние адренергического медиатора. По механизму действия также делятся на 2 группы :
1. Адреноблокаторы. Это препараты, которые блокируют рецепторы и тем самым препятствуют действию медиатора .
2. Симпатолитики. Это средства, которые тормозят накопление медиатора в синаптической щели и тем самым уменьшают его возбуждающее действие на рецепторы .
Передача возбуждения с постганглионарных нервных окончаний симпатической нервной системы на клетки эффекторных органов в основном осуществляется норадреналином. Исходным продуктом биосинтеза норадреналина является незаменимая аминокислота фенилаланин, которая в печени гидроксилируется и превращается в тирозин (тирозин может поступать и с пищей). Тирозин в цитоплазме нервного окончания окисляется в диоксифенилаланин (ДОФА) и декарбоксилируется. Образующийся дофамин в некоторых структурах мозга, например, в экстрапирамидной системе, является медиатором. С помощью особой транспортной системы дофамин переносится в везикулу, где дофамин-гидроксилаза превращает его в норадреналин.
В нервном окончании имеются три фракции норадреналина: лабильный фонд, который выбрасывается из везикулы в цитоплазму, а затем в синаптическую щель при поступлении нервного импульса; стабильный (резервный) фонд, сохраняющийся до истощения лабильного фонда везикулы, и цитоплазматическая свободная фракция, которая состоит из норадреналина, не депонированного в везикулах (при их насыщении). Последняя пополняется также за счет молекул медиатора, реабсорбированных из синаптической щели ("обратный захват").
В нервных окончаниях биосинтез заканчивается норадреналином. Хромаффинные клетки надпочечников метилируют норадреналин, превращая в адреналин.
Адреналин, норадреналин, дофамин и другие аналогичные амины, содержащие в бензольном кольце оксигруппу в положениях 3, 4, носят название катехоламинов ("катехол" - обозначает ортодиоксибензол).
Нормальное функционирование синапса во многом зависит от транспортных систем, осуществляющих перенос дофамина и норадреналина из цитоплазмы в везикулу и обратный (или нейрональный) захват норадреналина (около 70%) адренергическими окончаниями из синаптической щели.
В цитоплазме нервного окончания норадреналин разрушается (дезаминируется) моноаминоксидазой (МАО), кроме фракции, депонированной в везикулах, в синаптической щели - катехолометилтрансферазой (КОМТ). Последняя разрушает также катехоламины, циркулирующие в крови.
Локализация, типы и функции адренорецепторов. Адренорецепторы расположены частично в клетках эффекторных органов, иннервируемых постганглионарными волокнами симпатической нервной системы, а частично - вне синапсов. Различают ( a и b -адренорецепторы, каждый из которых имеет 2 типа - a 1, a 2 и b 1, b 2:
a 1-адренорецепторы локализованы в постсинаптических мембранах;
a 2-адренорецепторы - расположены пресинаптически в ЦНС и на адренергических окончаниях, а также внесинаптически в стенке сосудов.
a 1-адренорецепторы широко представлены:
1) в сосудах; их возбуждение суживает сосуды кожи, слизистых оболочек, брюшной полости и повышает артериальное давление;
2) в радиальной мышце радужной оболочки глаза; при их активации мышца сокращается и зрачок расширяется, но внутриглазное давление не повышается;
3) в желудочно-кишечном тракте - возбуждение этих рецепторов снижает тонус и моторику кишечника, но усиливает сокращение сфинктеров;
4) в гладкой мускулатуре дистальных отделов бронхов; стимуляция a 1-рецепторов этой области ведет к сокращению просвета дистальных отделов дыхательных путей.
a 2-Адренорецепторы по механизму обратной отрицательной связи регулируют выброс норадреналина; при возбуждении центральных пресинаптических a 2-адренорецепторов угнетается сосудо-двигательный центр и снижается артериальное давление; активация периферических пресинаптических a 2-адренорецепторов тормозит выделение норадреналина в синаптическую щель, что приводит к падению артериального давления. Внесинаптические a 2- адренорецепторы локализуются во внутреннем слое сосудов и возбуждаются адреналином, циркулирующим в крови; сосуды при этом суживаются и артериальное давление повышается.
Постсинаптические b 1-адренорецепторы локализуются в мышце сердца. Их возбуждение повышает все функции сердца: автоматизм, проводимость, возбудимость, сократимость. Увеличивается частота (тахикардия) и сила сердечных сокращений, возрастает потребление кислорода миокардом. При угнетении b 1-адренорецепторов развиваются противоположные эффекты: брадикардия, снижается сократимость. сердечный выброс и потребность сердца в кислороде. Постсинаптические b 2-адренорецепторы характерны для мышц бронхов, сосудов скелетной мускулатуры, миометрия. Возбуждение b 2-адренорецепторов бронхов приводит к их расширению. Механизм этого эффекта следующий: стимуляция b 2-адренорецепторов активирует аденилатциклазу, накапливается цАМФ, связывающий свободный кальций, снижение уровня кальция приводит к расслаблению бронхиальных мышц. Нечто подобное происходит и в тучной клетке (связывание кальция цАМФ и блок мембраны), вследствие чего тормозится высвобождение медиаторов аллергии (гистамина, серотонина, медленно реагирующей субстанции анафилаксии - ЛД4 и др.). Возбуждение b 2-адренорецепторов лежит в основе расширения сосудов (расслабление гладкомышечного слоя) скелетной мускулатуры, сердца, мозга, печени. Механизм положительной обратной связи реализуется пресинаптическими b 2-адренорецепторами: их возбуждение повышает выброс норадреналина.
Адренергические средства по аналогии с холинергическими разделяют на миметики и блокаторы.
Адренергические нейроны расположены в ЦНС (голубое пятно среднего мозга, мост, продолговатый мозг) и симпатических ганглиях.
Периферические адренергические синапсы образованы варикозными утолщениями разветвлений постганглионарных симпатических волокон.
Медиатором адренергических синапсов является норадреналин. Его биохимический предшественник дофамин выполняет медиаторную функцию в дофаминергических синапсах. Адреналин - гормон мозгового слоя надпочечников. Все три вещества относятся к группе катехоламинов, так как содержат гидроксильные группы в 3-м и 4-м положении ароматического кольца.
В гранулах норадреналин депонирован в связи с АТФ и белком хромогранином. В составе гранул обнаружены также ферменты и модулирующие нейропептиды (энкефалины, нейропептид Y).
Норадреналин синтезируется из аминокислоты тирозина. Превращение фенилаланина в тирозин является неспецифическим процессом и происходит в печени. Обе аминокислоты содержатся в больших количествах в твороге, сыре, шоколаде, бобовых.
Тирозин активным транспортом поступает в адренергические окончания. В их аксоплазме он приобретает второй гидроксил-радикал в 3-м положении ароматического кольца и превращается в диоксифенилаланин (ДОФА). Эту реакцию катализирует тирозингидроксилаза митохондрий. Затем ДОФА декарбоксилазой ароматических L-аминокислот декарбоксилируется в дофамин. Дофамин из аксоплазмы транспортируется в гранулы. На последнем этапе дофамин приобретает третий гидроксил в β-положении боковой цепи при участии дофамин- β-гидроксилазы.
В мозговом слое надпочечников норадреналин выходит из гранул и в цитоплазме метилируется в гормон адреналин под действием N-метилтрансферазы (донатором метильных групп служит S-аденозил-метионин) (рис. 9-1). Образование адреналина повышают глюкокортикоиды, эстрогены и тироксин. Глюкокортикоиды, поступая в мозговой слой по воротной системе надпочечников, активируют тирозингидроксилазу, дофамин- β-гидроксилазу и N-метилтрансферазу. У некоторых видов акул корковый и мозговой слои надпочечников представляют собой изолированные железы, поэтому у них адреналин не синтезируется, а единственным гормоном хромаффинных клеток является норадреналин.
• Нейронального захвата (захвата-1) - активного транспорта вначале через пресинаптическую мембрану (сопряжен с выходом ионов натрия), а затем через мембрану гранул под влиянием протонной АтФазы (при входе в гранулу одной молекулы норадреналина в аксоплазму выходят 2 протона).
• Экстранейронального захвата (захвата-2) нейроглией, фибробластами, кардиомиоцитами, клетками эндотелия и миоцитами сосудистой стенки.

Рис. 9-1.Биосинтез адреналина
Около 80% норадреналина подвергается нейрональному захвату и по 10% - экстранейрональному захвату и ферментативному расщеплению. Необходимость нейронального захвата диктуется дефицитом субстратов и большой потребностью в энергии для синтеза норадреналина из тирозина.
Для сохранения адреналина основное значение имеет экстранейрональный захват.
Ферменты инактивации катехоламинов - МАО и катехол-О-метил-трансфераза (КОМТ). МАО, локализованная на внешней мембране митохондрий и в гранулах, осуществляет окислительное дезаминирование катехоламинов с образованием биогенных альдегидов. Затем альдегиды окисляются НАД-зависимой альдегиддегидрогеназой в кислоты или восстанавливаются альдегидредуктазой в гликоли.
Цитоплазматический фермент КОМТ катализирует присоединение метильной группы к гидроксилу в 3-м положении ароматического кольца (только при наличии гидроксила в 4-м положении). Донатором метильных групп служит S-аденозилметионин. Метилированные продукты в 200-2000 раз (по результатам разных тестов) менее активны, чем норадреналин и адреналин.
АДРЕНОРЕЦЕПТОРЫ
В 1948 г. английский фармаколог Рассел Алквист выдвинул гипотезу о двух типах адренорецепторов. α-Адренорецепторы суживают сосуды, наиболее чувствительны к эпинефрину, меньше реагируют на норэпинефрин и очень слабо воспринимают действие изопреналина (изопропилнорадреналина). β-Адренорецепторы расширяют сосуды, обладают максимальной чувствительностью к изопреналину, в 10-50 раз слабее реагируют на эпинефрин и норэпинефрин.
Адренорецепторы обнаружены на постсинаптической, пресинаптической мембранах и в клетках, не получающих адренергической иннервации. Постсинаптические адренорецепторы имеют индексы 1 или 2, пресинаптические и внесинаптические адренорецепторы обозначаются индексом 2. Внесинаптические адренорецепторы активируются циркулирующими в крови норадреналином и адреналином. β-Адренорецепторы жировой ткани имеют индекс 3.
Все адренорецепторы характеризуются сходной последовательностью аминокислот (у α1- и α2-адренорецепторов идентичны 30% аминокислот, у β1- и β2-адренорецепторов - 60%).
В сосудах и внутренних органах расположены α- и β-адренорецепторы различных типов. Например, в сосудах легких обнаружено 30% β1-адренорецепторов и 70% β,,-адренорецепторов.
Адренорецепторы являются гликопротеинами, ассоциированы с G-белками и имеют такое же строение, как и другие рецепторы этого типа. Их белковая цепь состоит из семи гидрофобных доменов в виде трансмембранной спирали, Домены соединены гидрофильными петлями, расположенными попеременно по обе стороны мембраны. N-конец белковой молекулы адренорецептора расположен внеклеточно, C-конец - внутри клетки. Активный центр адренорецептора представляет собой карман, образованный высококонсервативными аминокислотами, расположенными в средней и во внеклеточной третях гидрофобных трансмембранных спиралей. Аминогруппа катехоламинов соединяется ионной связью с карбоксилом аспарагиновой кислоты в третьем трансмембранном домене. Гидроксилы катехоламинов образуют водородную связь с остатками серина в пятом и седьмом доменах, что необходимо для активации адренорецепторов.
Сведения о механизмах функционирования, чувствительности к агонистам и антагонистам, физиологической роли адренорецепторов представлены в табл. 9-2-9-4.
Таблица 9-2.Адренорецепторы и их эффекторные системы
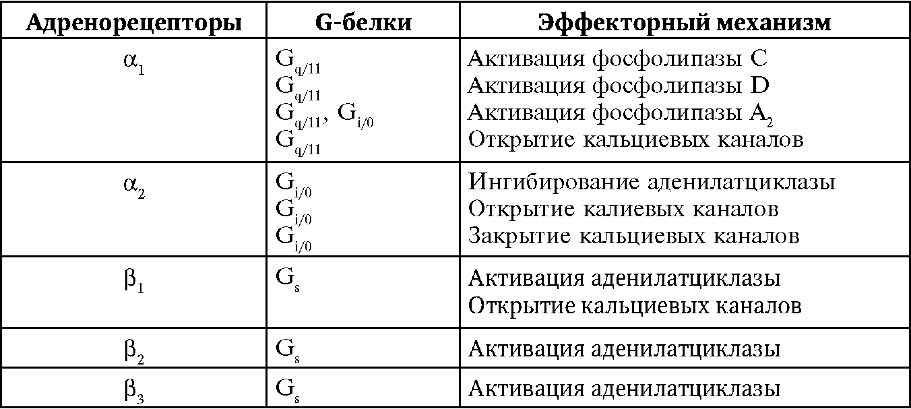
α-Адренорецепторы
Постсинаптические α1-адренорецепторы (типы A, B, D) активируют мембранные фосфолипазы и увеличивают проницаемость кальциевых каналов. В гладких мышцах ионы кальция активируют
Таблица 9-3.Адренорецепторы
Примечания. Э - эпинефрин; НЭ - норэпинефрин; И - изопреналин.
Таблица 9-4.Влияние вегетативной нервной системы на функции эффекторных органов
| Эффекторный орган | Адренергическое влияние | Холинергическое влияние (м-холинорецепторы*), функции** | |
| тип рецептора | функции** | ||
| Глаза | |||
| Радиальная мышца радужки | α1 | Расширение зрачков (мидриаз) ++ | - |
| Круговая мышца радужки | - | - | Сужение зрачков (миоз) +++ |
| Цилиарная мышца | β2 | Расслабление (для ясного видения вдали)+ | Сокращение (для ясного видения вблизи) +++ |
| Слезные железы | α | Повышение секреторной функции + | Повышение секреторной функции +++ |
| Миокард*** | |||
| Синусный узел | β1, β2 | Учащение сердечных сокращений ++ | Урежение сердечных сокращений +++ Вагусная остановка сердца |
| Предсердия | β1, β2 | Повышение сократимости и скорости проведения возбуждения ++ | Уменьшение сократимости, укорочение потенциалов действия ++ |
| Атриовентрикулярный узел | β1, β2 | Повышение автоматизма и проводимости ++ | Уменьшение проводимости, атриовентрикулярная блокада +++ |
| Система Гиса-Пуркинье | β1, β2 | Повышение автоматизма и проводимости ++ | Действие слабое |
| Желудочки | β1, β2 | Повышение сократимости, автоматизма и скорости проведения возбуждения +++ | Незначительное уменьшение сократимости |
Продолжение табл. 9-4
| Эффекторный орган | Адренергическое влияние | Холинергическое влияние (м-холинорецепторы*), функции** | |
| тип рецептора | функции** | ||
| Артериолы**** | |||
| Коронарные | α1, α2 β2 | Сужение + Расширение ++ | Сужение + |
| Кожи и слизистых оболочек | α1, α2 | Сужение +++ | - |
| Скелетных мышц | β2 | Расширение +++ | Расширение + |
| Головного мозга | α1 | Слабое сужение | - |
| Легких | α1 β2 | Сужение + Расширение + | - |
| Органов брюшной полости | α1 | Сужение +++ | - |
| Почек | α1 | Сужение +++ | - |
| Вены | α1, α2 β2 | Сужение ++ Расширение ++ | - |
| Легкие | |||
| Гладкие мышцы трахеи и бронхов | β2 | Расслабление + | Сокращение ++ |
| Бронхиальные железы | β2 | Уменьшение секреторной функции + | Повышение секреторной функции +++ |
| Слюнные железы | α1 β | Секреция К + и воды + Секреция амилазы + | Секреция К + и воды ++ + |
Продолжение табл. 9-4
| Эффекторный орган | Адренергическое влияние | Холинергическое влияние (м-холинорецепторы*), функции** | |
| тип рецептора | функции** | ||
| Желудок | |||
| Перистальтика и тонус | α1, α2, β2 | Уменьшение + | Повышение +++ |
| Сфинктеры | α1 | Сокращение + | Расслабление + |
| Секреция желудочного сока | - | Уменьшение (?) | Стимуляция +++ |
| Кишечник | |||
| Перистальтика и тонус | α1, α2, β1, β2 | Уменьшение + | Повышение +++ |
| Сфинктеры | α1 | Сокращение + | Расслабление + |
| Секреция кишечного сока | α2 | Уменьшение + | Стимуляция ++ |
| Печень | β, | Гликогенолиз, глюконеогенез +++ | - |
| Желчный пузырь и желчные протоки | β2 | Расслабление + | Сокращение + |
| Поджелудочная железа | |||
| Ацинусы | α | Уменьшение секреторной функции + | Повышение секреторной функции ++ |
| β-Клетки островков | β2 | Уменьшение секреции инсулина +++ Стимуляция секреции инсулина + | - |
| Почки | |||
| Секреция ренина | β1 | Повышение ++ | - |
| Мочевой пузырь | |||
| Мышца-детрузор | β2 | Расслабление + | Сокращение +++ |
| Треугольник и сфинктер | α1 | Сокращение ++ | Расслабление ++ |
Окончание табл. 9-4
| Эффекторный орган | Адренергическое влияние | Холинергическое влияние (м-холинорецепторы*), функции** | |
| тип рецептора | функции** | ||
| Мочеточники | |||
| Перистальтика и тонус | α1 | Повышение | Повышение (?) |
| Матка | α1,β2 | Во время беременности: сокращение (otj) или расслабление (β2) Вне беременности: расслабление (β9) | Сокращение ++ |
| Мужские половые органы | α1 | Эякуляция ++ | Эрекция +++ |
| Капсула селезенки | α1 | Сокращение +++ | - |
| Кожа | |||
| Пиломоторные мышцы | α1 | Сокращение ++ | - |
| Потовые железы | - | - | Повышение секреторной функции |
| Скелетные мышцы | β2 | Повышение сократимости, гликогенолиз, вход К + | - |
| Жировая ткань | β3 | Липолиз +++ | - |
* В гладких мышцах и железах находятся различные типы м-холинорецепторов (преобладают м-холинорецепторы 3-го типа), в сердце локализованы м2-холинорецепторы.
** Указана сила влияния на соответствующие функции.
*** В сердце преобладают β1-адренорецепторы.
**** Указаны преобладающие типы адренорецепторов, в артериолах органов брюшной полости и почек находятся сосудорасширяющие рецепторы дофамина.
кальмодулинзависимую киназу легких цепей миозина, что необходимо для образования актомиозина и сокращения. Только в желудке и кишечнике α1-адренорецепторы, открывая кальцийзависимые калиевые каналы, вызывают гиперполяризацию сарколеммы и расслабление гладких мышц. Эффекты активации α1-адренорецепторов следующие:
• сужение сосудов кожи, слизистых оболочек, органов пищеварения, почек и головного мозга;
• сокращение капсулы селезенки с выбросом депонированной крови в циркуляторное русло;
• сокращение сфинктеров пищеварительного тракта и мочевого пузыря;
• подавление моторики и снижение тонуса желудка и кишечника. α2-Адренорецепторы (типы A, B, C) локализованы на постсинаптической, пресинаптической мембранах и вне синапсов.
Пресинаптические α2-адренорецепторы ингибируют аденилатциклазу и тормозят синтез цАМФ, увеличивают проницаемость мембран для K+ с развитием гиперполяризации, блокируют кальциевые каналы. По принципу отрицательной обратной связи они тормозят выделение норадреналина из адренергических окончаний при избыточной активации адренорецепторов.
Постсинаптические α2-адренорецепторы суживают сосуды кожи и слизистых оболочек, угнетают моторику желудка и кишечника, подавляют секрецию кишечного сока.
Внесинаптические α2-адренорецепторы суживают сосуды кожи и слизистых оболочек, угнетают моторику желудка и кишечника, секрецию инсулина, повышают агрегацию тромбоцитов.
β-Адренорецепторы
β-Адренорецепторы, активируя аденилатциклазу, вызывают превращение АТФ в цАМФ - активатор цАМФ-зависимых протеинкиназ. Протеинкиназа А транспортируется в ядро клеток и фосфорилирует фактор транскрипции - ДНК-связывающий белок. Этот белок регулирует активность цАМФ-чувствительного элемента в промоторном участке гена, в результате активируется синтез β-адренорецепторов.
Для постсинаптических β1-адренорецепторов характерны следующие эффекты:
• стимуляция сердечной деятельности: тахикардия, ускорение проведения возбуждения по проводящей системе, усиление сокращений миокарда, увеличение потребности в кислороде (β1-адренорецепторы при участии Gs-белков стимулируют фосфорилирование кальциевых каналов, что сопровождается их открытием, входом в саркоплазму Ca 2 + и мобилизацией этого иона из саркоплазматического ретикулума; также фосфорилируется белок фосфоламбан, способствующий депонированию Ca 2 + в саркоплазматическом ретикулуме);
• стимуляция секреции ренина;
• угнетение моторики кишечника.
Постсинаптические и внесинаптические β2-адренорецепторы расслабляют гладкие мышцы и вызывают гипергликемию. В гладких мышцах они при участии цАМФ снижают активность киназы легких цепей миозина, в печени и скелетных мышцах активируют фермент гликогенолиза фосфорилазу, ингибируют гликогенсинтазу. Кроме того, в гладких мышцах блокируются кальциевые каналы и активируются калиевые каналы. Типичные эффекты β2-адренорецепторов следующие:
• расширение сосудов сердца, легких и скелетных мышц;
• расширение бронхов и угнетение секреторной функции бронхиальных желез;
• угнетение моторики желудка и кишечника;
• расслабление желчного пузыря, мочевого пузыря, матки;
• усиление цАМФ-зависимого гликогенолиза и глюконеогенеза;
• активация гликогенолиза в скелетных мышцах;
• стимуляция секреции инсулина.
Пресинаптические β2-адренорецепторы осуществляют обратную связь, стимулируя выделение норадреналина при недостаточной активации адренорецепторов.
β3-Адренорецепторы усиливают цАМФ-зависимый липолиз в жировых депо с повышением в крови содержания свободных жирных кислот. Полиморфизм гена β3-адренорецептора создает предрасположенность к развитию ожирения и сахарного диабета 2-го типа.
β-Адренорецепторы подвергаются гомологической и гетерологической десенситизации. Гомологическая десенситизация развивается только к действию β-адреномиметиков. Под их влиянием медленно фосфорилируется специфическая цАМФ-независимая протеинкиназа - киназа β-адренорецепторов. Этот фермент фосфорилирует комплекс "β-адренорецепто-β^-адреномиметик". В дальнейшем присоединяется белок β-аррестин, нарушающий связь регуляторного домена β-адренорецепторов с G-белком. Инактивированный комплекс подвергается интернализации в цитоплазму и деградации в лизосомах.
При гетерологической десенситизации в ответ на действие β-адреномиметиков ослабляется реакция на многие вещества, повышающие синтез цАМФ. Она обусловлена быстрым (в течение миллисекунд) фосфорилированием β-адренорецептора протеинкиназой А с последующей утратой способности передавать сигнал на эффекторные системы.
Дата добавления: 2016-02-20 ; просмотров: 3309 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Читайте также:
