
Механизмы преобразования энергии фотона в нервный импульс
Нервная система человека выступает своеобразным координатором в нашем организме. Она передаёт команды от мозга мускулатуре, органам, тканям и обрабатывает сигналы, идущие от них. В качестве своеобразного носителя данных используется нервный импульс. Что он собой представляет? С какой скоростью работает? На эти, а также на ряд других вопросов можно будет найти ответ в этой статье.
Чем является нервный импульс?
Исследование строения и работы
Впервые прохождение нервного импульса было продемонстрировано немецкими учеными Э. Герингом и Г. Гельмгольцем на примере лягушки. Тогда же и было установлено, что биоэлектрический сигнал распространяется с указанной ранее скоростью. Вообще, такое является возможным благодаря особенному построению нервных волокон. В некотором роде они напоминают электрический кабель. Так, если проводить параллели с ним, то проводниками являются аксоны, а изоляторами – их миелиновые оболочки (они являют собой мембрану шванновской клетки, которая намотана в несколько слоев). Причем скорость нервного импульса зависит в первую очередь от диаметра волокон. Вторым по важности считается качество электрической изоляции. Кстати, в качестве материала организмом используется липопротеид миелин, который обладает свойствами диэлектрика. При прочих равных условиях, чем больше будет его слой, тем быстрее будут проходить нервные импульсы. Даже на данный момент нельзя сказать, что эта система полноценно исследована. Многое, что относится к нервам и импульсам, ещё остаётся загадкой и предметом исследования.
Особенности строения и функционирования
Где они создаются?
Типы клеток
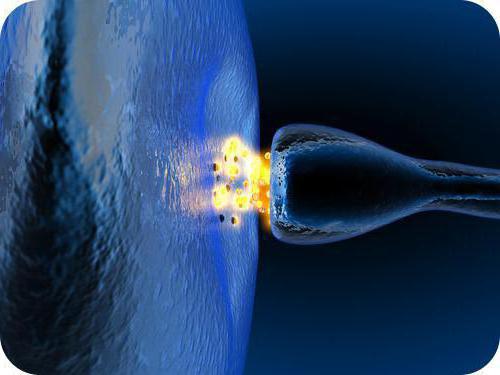
- Рецепторные (чувствительные). Ими кодируются и превращаются в нервные импульсы все температурные, химические, звуковые, механические и световые раздражители.
- Вставочные (также называются кондукторными или замыкательными). Они служат для того, чтобы перерабатывать и переключать импульсы. Наибольшее их число находится в головном и спинном мозге человека.
- Эффекторные (двигательные). Они получают команды от центральной нервной системы на то, чтобы были совершены определённые действия (при ярком солнце закрыть рукой глаза и так далее).
Каждый нейрон имеет тело клетки и отросток. Путь нервного импульса по телу начинается именно с последнего. Отростки бывают двух типов:
- Дендриты. На них возложена функция восприятия раздражения расположенных на них рецепторов.
- Аксоны. Благодаря им нервные импульсы передаются от клеток к рабочему органу.
Интересный аспект деятельности
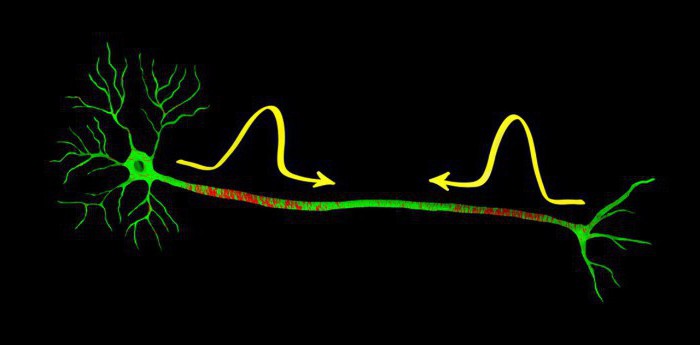
О потенциале действия
Как всё работает в мозгу?
Работа нейромедиаторов
Когда они передают нервные импульсы, то существует несколько вариантов, что произойдёт с ними:
- Они будут диффундированы.
- Подвергнутся химическому расщеплению.
- Вернутся назад в свои пузырьки (это называется обратным захватом).
В конце 20-го века сделали поразительное открытие. Ученые узнали, что лекарства, что влияют на нейромедиаторы (а также их выброс и обратный захват), могут изменять психическое состояние человека коренным образом. Так, к примеру, ряд антидепрессантов вроде "Прозака" блокируют обратный захват серотонина. Есть определённые причины считать, что в болезни Паркинсона виноват дефицит в головном мозге нейромедиатора дофамина.
Если кратко, то они могут работать с тысячами нейромедиаторов, которые посылаются их соседями. Детали относительно обработки и интеграции данного типа импульсов нам почти не известны. Хотя над этим работает много исследовательских групп. На данный момент получилось узнать, что все полученные импульсы интегрируются, а нейрон выносит решение – необходимо ли поддерживать потенциал действия и передавать их дальше. На этом фундаментальном процессе базируется функционирование головного мозга человека. Ну что ж, тогда это неудивительно, что мы не знаем ответа на эту загадку.
Некоторые теоретические особенности
Где же создаются нервные импульсы?
Откуда они начинают свой путь? Ответ на этот вопрос может дать любой студент, который прилежно изучал физиологию возбуждения. Есть четыре варианта:
- Рецепторное окончание дендрита. Если оно есть (что не факт), то возможным является наличие адекватного раздражителя, что создаст сначала генераторный потенциал, а потом уже и нервный импульс. Подобным образом работают болевые рецепторы.
- Мембрана возбуждающего синапса. Как правило, такое возможно только при наличии сильного раздражения или их суммирования.
- Триггерная зона дентрида. В этом случае локальные возбуждающие постсинаптические потенциалы формируются как ответ на раздражитель. Если первый перехват Ранвье миелинизирован, то они на нём суммируются. Благодаря наличию там участка мембраны, которая обладает повышенной чувствительностью, здесь возникает нервный импульс.
- Аксонный холмик. Так называют место, где начинается аксон. Холмик – это наиболее частый создать импульсов на нейроне. Во всех остальных местах, которые рассматривались ранее, их возникновение гораздо менее вероятное. Это происходит из-за того, что здесь мембрана имеет повышенную чувствительность, а также пониженный критический уровень деполяризации. Поэтому, когда начинается суммирование многочисленных возбуждающих постсинаптических потенциалов, то раньше всего на них реагирует холмик.
Пример распространяющегося возбуждения
Вспомните сводки из новостей прошлого лета (также это скоро можно будет услышать опять). Пожар распространяется! При этом деревья и кустарники, которые горят, остаются на своих местах. А вот фронт огня идёт всё дальше от места, где был очаг возгорания. Аналогичным образом работает нервная система.
Часто бывает необходимо успокоить начавшееся возбуждение нервной системы. Но это не так легко сделать, как и в случае с огнем. Для этого совершают искусственное вмешательство в работу нейрона (в лечебных целях) или используют различные физиологические средства. Это можно сравнить с заливанием пожара водой.
Формулирование законов полярного, "силы-времени", градиента, физиологического электротона. Свойства и классификация рецепторов. Механизм преобразования энергии стимула в нервный импульс. Пути фармакологической регуляции проводимости и лабильности.
Нажав на кнопку "Скачать архив", вы скачаете нужный вам файл совершенно бесплатно.
Перед скачиванием данного файла вспомните о тех хороших рефератах, контрольных, курсовых, дипломных работах, статьях и других документах, которые лежат невостребованными в вашем компьютере. Это ваш труд, он должен участвовать в развитии общества и приносить пользу людям. Найдите эти работы и отправьте в базу знаний.
Мы и все студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будем вам очень благодарны.
Чтобы скачать архив с документом, в поле, расположенное ниже, впишите пятизначное число и нажмите кнопку "Скачать архив"
| Рубрика | Медицина |
| Вид | лекция |
| Язык | русский |
| Дата добавления | 30.07.2013 |
| Размер файла | 26,6 K |
- посмотреть текст работы
- полная информация о работе
Законы раздражения возбудимых тканей и следствия, которые из них вытекают. Физиология человека, регуляция и сенсорное обеспечение движений. Минимальная сила раздражителя. Законы силы, времени и адаптации организма человека. Наличие внешнего раздражителя.
контрольная работа [16,7 K], добавлен 23.07.2009
Правила по технике безопасности при работе в физиологической лаборатории. Этапы приготовления нервно-мышечного препарата. Строение и физиологические функции биологических мембран возбудимых тканей. Первый и второй опыты Гальвани. Порог раздражения мышцы.
методичка [1,4 M], добавлен 07.02.2013
Механизм передачи возбуждения в химическом синапсе, особенности его строения. Виды и свойства медиаторов. Электрические и тормозные синапсы, особенности передачи сигнала. Пути фармакологической регуляции возникновения синаптического возбуждения.
презентация [2,1 M], добавлен 09.12.2014
Чувствительные нервные окончания. Супрапороговое раздражение рецепторов. Лекарственные средства, понижающие и стимулирующие чувствительность окончаний нервов. Механизм действия местных анестетиков. Блокада проведения нервных импульсов по аксонам.
презентация [233,2 K], добавлен 13.04.2015
Классификация специфических холинергических рецепторов с фармакологической точки зрения: никотиновые и мускариновые. Немедиаторное действие ацетилхолина в целостном организме. Действие ацетилхолина на постсинаптическую мембрану постганглионарных нейронов.
реферат [469,9 K], добавлен 11.02.2014
Понятие о физиологических функциях и их регуляции. Механизм и законы проведения возбуждения. Функциональное значение его структурных элементов нейрона. Особенности строения и функций вегетативной нервной системы. Строение и роль в организме надпочечников.
контрольная работа [22,2 K], добавлен 14.01.2010
Высшая нервная деятельность. Работа аппаратов рецепции и высших этажей мозга. Проблема адекватности отражения. Дифференциация раздражений, их дробный анализ. Энергия внешнего раздражения. Афферентная импульсация от мышечно-суставных рецепторов.
реферат [18,7 K], добавлен 16.06.2013
Оpганизм, его стpоение и жизнедеятельность. Совpеменные пpедставления о стpоении и функции мембpан. Биоэлектpические явления в живых тканях. Возбуждение: потенциал действия, механизм пpоисхождения, фазы. Раздpажитель: значение, виды и классификация.
лекция [25,4 K], добавлен 26.04.2012
Понятие и функции органов чувств как анатомических образований, воспринимающих энергию внешнего воздействия, трансформирующих ее в нервный импульс и передающих этот импульс в мозг. Строение и значение глаза. Проводящий путь зрительного анализатора.
презентация [667,7 K], добавлен 27.08.2013
Раздражимость как основное свойство живых клеток. Физиология возбудимых клеток. Строение и основные свойства клеточных мембран и ионных каналов. Физиология нервной ткани и синапсов. Классификация антиадренергических средств, механизм их действия.
курсовая работа [194,6 K], добавлен 02.03.2014
В главе 16 излагаются химические аспекты структурно-функциональной организации нервной системы, при этом особое внимание обращается нахимические основы возникновения и передачи нервных импульсов,молекулярной рецепции, кратко рассматриваются современные представленияо механизмах памяти, обучения, эмоций и опущений (вкуса, запаха). Содержание данной главы логически взаимосвязано с содержанием последующих глав раздела. [c.456] МЕХАНИЗМЫ ВОЗНИКНОВЕНИЯ И ПЕРЕДАЧИ НЕРВНОГО ИМПУЛЬСА
Какие молекулярные механизмы лежат в основе возникновения ипередачи нервного импульса [c.474] Позвольте мне проиллюстрировать этот тезис. Истинная функция нейрона — передача сигналов. Однако мы увидим (гл. 5), что в нервной системе существуют только два типа сигналов электрические и химические. Важно отметить, что сам сигнал содержит очень мало информации. Его специфичность зависит от мест возникновения и приема, т. е. от клеток органов, между которыми он передается. Так, например, причина того, что мы слышим, а не видим звук, кроется не в электрическом или химическом коде нервного импульса, а в том, что зрительная кора затылочной доли головного мозга соединена с нейронами сетчатки, а не уха. При электрическом или механическом, а не оптическом воздействии на сетчатку мы также будем видеть . Любой, у кого искры из глаз сыпались после сильного удара, может подтвердить это. Следовательно, качественно информация, передаваемая нейроном, зависит исключительно от специфичности его соединения, и толькоколичественная характеристика содержится, по-видимому, в самом сигнале сильный стимулятор посылает больше нервных импульсов от рецептора к воспринимающему органу, чем слабый. Опять же нервные импульсы, скажем, оптической или акустической области нашей нервной системы практически неотличимы от нервных импульсов в совершенно других системах, например у более примитивных форм жизни. Сами по себе эти импульсы очень малоинформативны даже для узкого специалиста. Таким образом, нейрохимик, изучающий биохимию нейронов, может выяснить только механизм возникновенияи передачи сигналов, специфическое содержание (смысл) сигналов недоступно его методам. Он может изучать общие молекулярные реакции, лежащие в основе обработки сигналов, но не результаты этой обработки, т. е. информацию . [c.8] Какова бы ни была роль регуляции числа мотонейронов у эмбриона, ее механизм находится в интересной зависимости от мышечной активности. Узародышей позвоночных начинаются беспорядочные, конвульсивные движения почти сразу же после того, как у них начинают формироваться нервно-мышечные соединения. Эти движения-результат спонтанного возникновения нервных импульсов в центральной нервной системе, и они наблюдаются даже у тех эмбрионов, у которых сенсорные нейроны разрушены. Если зародыша обработать ядом, блокирующим передачу в нервно-мышечных соединениях (таким, как кураре), то даижения прекратятся. Можно было бы предположить, что подобное воздействие либо никак не повлияет на гибель мотонейронов, либо усилит ее. Насамом деле эта обработка дает противоположный эффект до тех пор пока поддерживается блокада, практически все мотонейроны сохраняются. Каков бы ни был механизм этого явления, ясно, что /мышечная активность важна длянормального развития двигательной систе мы, точно так же как получение внешних стимулов необходимо для нормального развития сенсорных систем (см. ниже) в обоих случаях электрические сигналы способствуют поддержанию нервных связей. [c.144] В настоящее время достаточно много известно о молекулярных механизмахвозникновения и проведения нервного импульса и механизмах синаптической передачи импульса. Что касается интегральных функций мозга, то мы не знаем, как работает наш мозг, например, когда мы протягиваем руку, чтобы нажать на определенную клавишу компьютера, а тем более когда мы избегаем опасности поставить неправильный диагноз болезни. Современное состояниенейробиохими-ческих исследований можно охарактеризовать лишь как поиски подходов к таким проблемам. [c.531]
Генерация нервного импульса представляет собой динамический процесс, сопровождающийся структурными изменениями в возбудимой мембране. Эти изменения проявляются как изменения ионной проницаемости, а также реакционноспособ-ности функциональных групп к специфичным реагентам. [1]
Генерация нервного импульса вызывается перемещением ионов через мембрану нервной ( или другой возбудимой) клетки. Схематически система, ответственная за генерацию импульса, может быть представлена в виде двух отсеков, ргзде-ленных мембраной. [2]
Исследование генерации нервного импульса и его распространения в аксоне - старая и традиционная проблема биофизики / Уже Гельмгольц измерял скорость распространения нервного возбуждения / В настоящее время решены важные физические задачи, относящиеся к функции аксона. Напротив, современное состояние науки позволяет лишь формально моделировать работу центральной нервной системы и мы еще далеки от понимания физической природы высших ее функций - памяти и мышления. [3]
Проблема генерации нервного импульса в ответ на раздражение аксона исследовалась экспериментально и теоретически во множестве работ. Эта проблема уже затрагивалась в § § 4.1 и 4.2. Трудности ее изучения в значительной мере связаны с отсутствием прямой информации о соответствующих молекулярных процессах. Приходится строить теоретические и полуэмпирические модели, основанные на косвенных данных, полученных в опытах, в которых мембраны подвергаются различным физическим и химическим воздействиям. [4]
Возможность возникновения концентрационных колебаний была показана в выдающихся исследованиях механизма генерации нервного импульса ( Hodgkjn, Huxley, 1952; Ходжкин, 1965) и внутриклеточной регуляции синтеза ферментов ( Mono. [5]
Следовательно, корреляция спайковой активности отдельных нейронов не зависит от самого механизмагенерации нервных импульсов. Поэтому должен существовать специальный механизм, осуществляющий вязь спайковых разрядов соседних нейронов. Скорее всего, функцию этого механизма эффективно выполняет микроструктура медленных потенциалов, возникающих на контактах между нейронами. Таким образом, маловероятно что устойчивое состояние и дискретные нейронные процессы являются взаимосвязанными процессами и отражают друг друга. Правила преобразования, которые позволяют перекодировать один процесс в другой, должны быть определены эмпирически для каждого из тех условий, которые интересуют исследователей. На данном этапе исследования любые общие утверждения относительно типов таких преобразований являются, вероятно, преждевременными. [6]
Во-первых, это любые триггерные, пороговые процессы переключения системы из одного режима в другой, например, генерация нервного импульса или сокращение мышцы. [7]
Чем значительнее емкость конденсатора, тем больше зарядов абсорбируется, до тех пор пока не достигается деполяризация, необходимая для генерации нервного импульса. [8]
Мы не в состоянии пока ответить на вопросы, каким образом фотовозбуждение единственной молекулы пигмента может вызвать прохождение тока через синапс на отдаленном конце рецепторной клетки и как благодаря ничтожно малой энергии всего лишь одного фотона могут осуществляться значительные перемещения заряда, необходимые для генерации нервного импульса. Считают, что молекулы пигментов каким-то образом контролируют ионные каналы через внутренние мембраны рецепторных клеток. Фотовозбуждение молекулы пигмента вызывает изменение ее конфигурации или заряда. В результате этого открывается канал и возникает поток ионов, который вызывает поляризацию клетки и таким образом стимулирует синапс. Затем ионные насосы должны получить энергию для восстановления нормального распределения ионов. Такой механизм может объяснить необходимое усиление и быстроту реакции на относительно большом расстоянии. [9]
Na и К 1; т, п и h - безразмерные нормированные величины, характеризующие плотность открытых каналов; т носит название натриевой активации; h - натриевая инактивация; п - калиевая активация; jt - неидентифицированный ток утечки; J0 - внешний ток, с помощью которого можно управлять режимом мембраны и вызывать генерацию нервных импульсов. [10]
Это неравновесное распределение ионов поддерживается специальными механизмами, встроенными в мембрану. Однако для генерации нервного импульса происхождение разности концентраций несущественно, важно только наличие этой разности. [11]
Модель описывает генерацию стандартного сигнала в ответ на малое, но конечно 1 внеш. Модель ( 5) используют при описании генерации нервного импульса, возникновения биол. [12]
Проблемы активного транспорта, описанные здесь, имеют непосредственное отношение к возбудимым мембранам. Рассмотрение свойств этих мембран в связи с генерацией нервного импульса приведено в гл. [1]
Эти несколько замечаний показывают, что данный случайный процесс более сложен, чем чистый дробовой шум. Возможно, что могло бы быть уместным предположение о некоторой корреляции между д моментамигенерации нервных импульсов. [2]
В течение многих лет было невозможно провести различие между потенциалами и теми пейсмекерамп, которые связаны с явлением нарастания и убывания нейроэлектрических состояний, возникающих в микроструктуре соединений. Но, как уже было отмечено, Соколов ( 1970) показал, что в определенных лабораторных условиях механизм генерации аксонных нервных импульсов мошет действовать относительно независимо от изменений медленных потенциалов мембраны тела клетки. [3]
Одна из них была хорошо обоснована, другая - нет. Первая касалась вопросов генерации нервных импульсов и их передачи через синапсы. Другая имела отношение к электрическим полям, создаваемым потенциалами мозга. [4]
К гДа внешний ток невелик, устанавливается новый стационарный потенциал. Однако если деполяризация превзойдет некоторый порог ( - - 20 мв), то дальнейший процесс деполяризации идет с самоускорением. Этот процесс и есть генерация нервного импульса. Он происходит в ответ на надпорого-вую ступенчатую деполяризацию мембраны. Если ступенчатая деполяризация достаточно велика, то наступает автогенерация. [5]
Поскольку современные биохимические и физико-химические методы развиваются очень быстро, можно ожидать, что вскоре накопится обширная информация о циклах превращений зрительных пигментов и их промежуточных продуктах, а также об опсин-хромофорных взаимодействиях, особенно для родопсина палочек. Пройдет, однако, еще немало времени, прежде чем станут известны все детали структуры некоторых короткоживу-щих промежуточных продуктов, что позволит оценить значение небольших изменений конформации, взаимодействий белок - - хромофор и особенностей поглощения света. Следует также выяснить механизм генерации нервного импульса в ответ на поглощение фотона зрительным пигментом. Даже после того как мы ответим на некоторые вопросы о функционировании родопсина у тех немногих видов, которые наиболее подробно изучены ( человек, крыса, крупный рогатый скот), предстоит огромная работа по изучению биохимии цветового зрения у млекопитающих, а также зрительных пигментов и циклов их превращений у других животных. [6]
Дедал вспоминает, что основу механизма памяти, как и всех прочих функций человеческого мозга, составляют так называемые синапсы - области контактов нейронов друг с другом и с клетками исполнительных органов. Сигнал, передаваемый от клетки к клетке, имеет химическую природу. По команде нейрона соответствующее нервное окончание выделяет особое химическое вещество-передатчик, которое вызывает генерацию нервного импульса в другой клетке. Новые нервные импульсы, распространяясь по нервным волокнам, вызывают в свою очередь возбуждение все новых нервных клеток. Механизм действия транквилизаторов обычно состоит в том, что молекулы этих веществ блокируют действие веществ-передатчиков; некоторые такие лекарства оказывают нежелательное побочное действие на память. [7]
Мембранная теория встречается, однако, с трудностями при попытках объяснить тепловые явления в нерве. Если перенос ионов адиабатичен, то система, совершая электрическую работу, должна охлаждаться. Эти тепловые явления до сих пор не объяснены. Несомненно, что мембрана аксона представляет собой диссипативную систему, и можно думать, что к рассмотрению генерации нервного импульса применима неравновесная термодинамика. Соответствующая теория еще не построена, она должна описать и теплопродукцию нерва. Подлинное объяснение этих явлений требует, однако, раскрытия их молекулярных механизмов. [8]
Он обнаружен в продуктах животного происхождения. Пигмент каротин ( оранжевого цвета), присутствующий в моркови, и схожие пигменты, называемые каротинами, часто встречающиеся в растениях, могут преобразовываться в витамин А в процессе пищеварения. Структура каротинов и витамина А особенно хорошо адаптирована для поглощения света, как в растениях в форме каротинов, так и у животных, у которых витамин А превращается в светопоглощающую молекулу ретиналь. Три группы животных, у которых имеются глаза ( моллюски, членистоногие и позвоночные), используют ретиналь в качестве светопоглощающей части фоторецепторных молекул. Свет вызывает довольно большие изменения в структуре ретиналя, достаточные для генерации нервного импульса. [9]
Эти данные, а также другая работа Томпсона ( Thompson and Spenser, 1966) показывают, что привыкание связано с взаимодействием нескольких нейронов. Томпсон использовал препарат, принесший известность Шеррингтону: животное, у которого спинной мозг отделен от головного. Ни входные, ни выходные нейроны не участвуют в изменении рефлекса и, следовательно, не ответственны за привыкание: решающее значение имеют интернейроны, - связывающие нейроны входа и выхода. Это значит, что привыкание не связано с изменениями медленных потенциалов соединительных контактов. Он обнаружил, что медленные потенциалы мембраны нейрона и генераторные потенциалы, записанные от его аксона, которые дают начало нервным импульсам, могут изменяться независимо друг от друга. Кроме того, полученные им результаты свидетельствуют о том, что механизм генерации нервных импульсов может обнаружить привыкание. [10]
Потенциал действия и нервный импульс
Характеристики мембранных потенциалов
| Показатели | Рецепторный потенциал | Постсинаптический потенциал | Потенциал действия |
| Амплитуда | 0,1-10 мВ | 0,1-10 мВ | 70-110 мВ |
| Длительность | 5-100 мс | 5 мс — 20 мин | 1-10 мс |
| Время/пространство | Убывающий | Убывающий | Незатухающий |
| Движение | Локальный | Локальный | Распространяющийся |
| Функциональная зависимость | Величина зависит от силы раздражителя | Величина зависит от количества нейромедиатора | Амплитуда стандартна для данного нейрона и не зависит от силы раздражителя или от количества нейромедиатора |
Понятие потенциала действия и нервного импульса. Описание графика потенциала действия и механизмов нервного импульса.
Строго говоря, движущееся по нервам возбуждение представляет собой нервные импульсы, а не потенциалы действия.
Нервный импульс - это движущаяся волна изменений в состоянии мембраны, включающая в себя структурные изменения (открытие и закрытие ионных каналов), химические (изменяющиеся потоки ионов) и электрические (деполяризацию, позитивную поляризацию и реполяризацию).
Но в физиологической литературе в качестве синонима для нервного импульса принято использовать также и термин "потенциал действия". Хотя потенциал действия - это только электрический компонент нервного импульса.
Потенциал действия - это электрический компонент нервного импульса, характеризующий изменения электрического заряда (потенциала) на локальном участке мембраны во время прохождения через него нервного импульса (от -70 до +30 мВ и обратно).
Сравните два приведённых выше рисунка (покликайте по ним) и, как говорится, почувствуйте разницу!
Где рождаются нервные импульсы?
Как ни странно, не все студенты, изучившие физиологию возбуждения могут ответить на этот вопрос. ((
Хотя ответ не сложен. Нервные импульсы рождаются на нейронах всего в нескольких местах:
1) аксонный холмик,
2) рецепторное окончание дендрита,
3) первый перехват Ранвье на дендрите (триггерная зона дендрита),
4) постсинаптическая мембрана возбуждающего синапса.
Места возникновения нервных импульсов:
1. Аксонный холмик - главный породитель нервных импульсов.
Аксонный холмик - это самое начало аксона, там где он начинается на теле нейрона. Именно аксонный холмик является главным породителем нервных импульсов на нейроне. Во всех остальных местах вероятность рождения нервного импульса намного меньше. Дело в том, что у мембраны аксонного холмика повышена чувствительность к возбуждению и понижен критический уровень деполяризации (КУД) по сравнению с остальными участками мембраны. Поэтому, когда на мембране нейрона начинают суммироваться многочисленные возбуждающие постсинаптические потенциалы (ВПСП), которые возникают в самых разных местах на постсинаптических мембранах всех его синаптических контактов, то раньше всего КУД достигается именно на аксонном холмике. Там-то эта сверхпороговая для холмика деполяризация и открывает потенциал-чувствительные натриевые каналы, в которые входит поток ионов натрия, порождающий потенциал действия и нервный импульс.
Итак, аксонный холмик является интегративной зоной на мембране, он интергрирует все возникающие на нейроне локальные потенциалы (возбуждающие и тормозные) - и первый срабатывает на достижение КУД, порождая нервный импульс.
Важно также учесть следующий факт. От аксонного холмика нервный импульс разбегается по всей мембране своего нейрона: как по аксону к пресинаптическоим окончаниям, так и по дендритам к постсинаптическим "начинаниям". Все локальные потенциалы при этом снимаются с мембраны нейрона и со всех его синапсов, т.к. они "перебиваются" потенциалом действия от пробегающего по всей мембране нервного импульса.
2. Рецепторное окончание чувствительного (афферентного) нейрона.
Если нейрон имеет рецепторное окончание, то на него может воздействовать адекватный раздражитель и порождать на этом окончании сначала генераторный потенциал, а затем и нервный импульс. Когда генераторный потенциал достигает КУД, то на этом окончании открываются потенциал-зависимые натриевые ионные каналы и рождается потенциал действия и нервный импульс. Нервный импульс бежит по дендриту к телу нейрона, а затем по его аксону к пресинаптическим окончаниям для передачи возбуждения на следующий нейрон. Так работают, к примеру, болевые рецепторы (ноцицепторы), являющиеся дендритными окончаниями болевых нейронов. Нервные импульсы в болевых нейронах вознимают именно на рецепторных окончаниях дендритов.
3. Первый перехват Ранвье на дендрите (триггерная зона дендрита).
Локальные возбуждающие постсинаптические потенциалы (ВПСП) на окончаниях дендрита, которые формируются в ответ на возбуждения, приходящие к дендриту через синапсы, суммируются на первом перехвате Ранвье этого дендрита, если он, конечно, миелинизирован. Там находится участок мембраны с повышенной чувствительностью к возбуждению (пониженным порогом), поэтому именно в этом участке легче всего преодолевается критический уровень деполяризации (КУД), после чего открываются потенциал-управляемые ионные каналы для натрия - и возникает потенциал действия (нервный импульс).
4. Постсинаптическая мембрана возбуждающего синапса.
В редких случаях ВПСП на возбуждающем синапсе может быть настолько силён, что прямо там же достигает КУД и порождает нервный импульс. Но чаще это бывает возможно только в результате суммации нескольких ВПСП: или с нескольких соседних синапсов, сработавших одновременно (пространственная суммация), или за счёт того, что на данный синапс пришло несколько импульсов подряд (временная суммация).
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Читайте также:
