
Мера лабильности нервов мышц и нервно-мышечных синапсов
В синапсе различают пресинаптичзское окончание аксона, пресинаптическую мембрану, синаптическую щель и постсинаптическую мебрану. Часть медиатора синтезируется в теле нервной клетки, и затем с помощью аксонного транспорта со скоростью около 40 см/сутки попадает в пресинаптическое окончание. Другая часть медиатора синтезируется непосредственно в пресинаптическом окончании. Порции медиатора окружаются мембранами и в виде везикул хранятся в пресинаптическом окончании. При проведении возбуждения ионы Са ++ входят в пресинаптическое окончание и присоединяются к везикулам, обеспечивая их перемещение к пресинаптической мембране и слияние с ней. Медиатор выходит в синаптическую щель, где с помощью диффузии пассивно передвигается к постсинаптической мембране.
Медиатор действует на постсинаптические рецепторы, образованные белково-липидными молекулами, что приводит к открытию пор для ионов Nа + и деполяризации постсинаптической мембраны. При этом на мембране возникает возбуждающий постсинаптический потенциал (ВПСП), который за счёт местных токов вызывает потенциал действия в соседних участках поверхностной мембраны клетки и далее проводится по ней (рис. 1).
Подействовавший на постсиналтическую мебрану медиатор разрушается ферментами. Продукты его распада всасываются в кровь или закачиваются в пресинаптическое окончание, где происходит синтез новых порций медиатора. Ацетилхолин расщепляется холинзстеразой, а норадреналин расщепляется последовательно действующими ферментами КОМТ и МАО.
В тормозных синапсах в качество медиатора выступает гаммааминомасляная кислота (ГАМК) или глицин, которые увеличивают проницаемость постсинаптической мембраны для ионов К + и С1 - . Ионы К + выходят, а ионы С1 - входят в клетку, в результате чего возникает гиперполяризация мебраны и на ней формируется тормозной постсинаптический потенциал (ТПСП) (рис. 1). Порог раздражения увеличивается, возбудимость падает и возбуждение не проводится, что и характеризует состояние торможения в нейрональных синапсах.
Рис. 1. Постсинаптические потенциалы.
Синапсы имеют определенные функциональные свойства.
Свойство 1. В синапсах осуществляется одностороннее проведение возбуждения от пре- к постсинаптической мембране.
Свойство 2. За счет диффузии медиатора в синаптической щели происходит синаптическая задержка проведения возбуждения.
Свойство 3. Наличие постсинаптических рецепторов обуславливает высокую чувствительность синапсов к химическим веществам.
Свойство 4. Постсинаптические потенциалы в синапсе не подчиняются закону "Все или ничего" и способны к суммации. Амплитуда постсинаптических потенциалов зависит от количества действующего на постсинаптическую мембрану медиатора.
Свойство 5. Синапсы имеют низкую лабильность по сравнению о лабильностью нервов и мышц. Лабильность или функциональная подвижность характеризуется быстротой появления и исчезновения возбуждения и количественно отражает минимальную длительность процесса возбуждения. Мерой лабильности является максимальная частота раздражения, которую возбудимая ткань воспроизводит без трансформации ритма. Количественно мера лабильности является обратной величиной от длительности возбуждения или точнее от длительности рефрактерного периода (таблица 1).
Свойство 6. Синапсы обладают повышенной утомляемостью, по сравнению с утомляемостью нервов и мышц при их длительном ритмическом раздражении. Утомляемость связана с истощением запасов медиатора в синапсах.
Величины лабильности нерва, мышцы и синапса.
| Возбудимое образование | Время абсолютного рефрактерного периода, мс | Мера лабильности, имп/с |
| Нерв миелиновый | ||
| Нерв безмиелиновый | ||
| Мышца поперечно-полосатая | ||
| Синапс нервно-мышечный |
Цель работы.Сравнить меру лабильности возбудимых образований нервно-мышечного препарата.
Оснащение:кимограф, универсальный штатив с вертикальным миографом, пластина с раздражающими электродами для нерва, электрический стимулятор, двухполюсный ключ, чернила, бумага, физиологический раствор для холоднокровных животных. Эксперимент выполняют на лягушке.
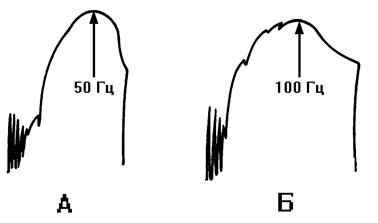
Рис. 2 Миограмма икроножной мышцы лягушки при увеличении частоты раздражения в ходе непрямого (А) и прямого (Б) раздражения.
Анализ результатов и выводы. В выводах указать структуру нервно-мышечного препарата, обладающую наименьшей лабильностью, объяснить причину наступления оптимума и пессимума при меньшей частоте раздражения в условиях непрямого раздражения мышцы.
Рекомендации к оформлению протокола.
1. Записать ход опыта, вклеить миограмму и сделать на ней обозначения. Указать оптимум и пессимум частоты при непрямом и прямом раздражении мышцы.
2. Записать вывод.
3. На основе теоретических знаний объясняют причины низкой или высокой лабильности различных возбудимых образований.
А - прямое раздражение мышцы; Б - непрямое раздражение мышцы
2. В выводах объяснить механизм возникновения оптимума и пессимума.
Задача 3. Локализация утомления в нервно-мышечном препарате
Под утомлением понимают временное понижение работоспособности клетки, органа или целого организма, наступающее в результате длительной работы и исчезающее после отдыха. В целостном организме, при совместной деятельности центральных и периферических аппаратов, явление утомления развивается прежде всего в структурах ЦНС. Из периферических образований наиболее быстро утомляются синапсы вследствие более низкой скорости происходящих в них химических процессов по сравнению с проведением электрических импульсов в нерве, уменьшения резерва медиаторами и ионов кальция и снижения чувствительности рецепторов постсинаптической мембраны к медиатору.
О с н а щ е н и е: см. Задачу 1.
С о д е р ж а н и е р а б о т ы. Приготовить нервно-мышечный препарат и укрепить его в миографе. Подобрать заведомо надпороговую величину тока, вызывающую сокращение мышцы как при прямом, так и при непрямом ее раздражении. Ключ установить в положение "Нерв" (непрямое раздражение мышцы) и при медленном вращении барабана кимографа включить ритмичное раздражение частотой 1 Гц. Записать сокращения мышцы вплоть до полного их исчезновения. Затем перевести ключ в положение "Мышца" (прямое раздражение). Обратить внимание на амплитуду первого после переключения сокращения мышцы. Продолжать регистрацию сокращений при ритмичном раздражении до полного утомления мышцы, отметить время его развития.
О ф о р м л е н и е п р о т о к о л а. 1. Вклеить кимограмму и сделать на ней обозначения
Развитие утомления в нервно-мышечном препарате
1 - сокращение мышцы при непрямом раздражении;
2 - сокращение мышцы при прямом раздражении;
а) - момент замыкания ключа в положении "Нерв";
б) - момент замыкания ключа в положении "Мышца".
2. Объяснить, почему утомление мышцы при непрямом ее раздражении развивается быстрее, чем при прямом.
Выводы
Домашнее задание:
§ В тетради для самостоятельной работы письменно ответьте на вопросы, предложенные преподавателем.
Подпись преподавателя
З А Н Я Т И Е 7,8
Физиология мышц. Зависимость амплитуды мышечного сокращения от силы и частоты раздражения. Сравнительная характеристика скелетных и гладких мышц. Динамометрия.
Мышечное сокращение. 1. Общие физические и физиологические свойства мышц. 2. Виды сокращения мышц. Виды работы. 3. Механизм сокращения скелетных мышц. 4. Функциональная лабильность. Оптимум и пессимум частоты раздражения. 5. Физиологические особенности скелетных мышц. 6. Физиологические особенности гладких мышц.
Задача 1. Зависимость амплитуды мышечных сокращений от силы раздражения
Скелетная мышца состоит из большого числа отдельных мышечных волокон. Амплитуда сокращения мышцы зависит от количества сократившихся в данный момент волокон. На одиночное пороговое раздражение каждое такое волокно возбуждается, а затем сокращается с максимальной амплитудой, т.е. отвечает по закону "все или ничего". Однако возбудимость различных типов волокон, составляющих мышцу, различна. Поэтому пороговая сила раздражения вызывает сокращение только наиболее возбудимых мышечных волокон, и величина ответа целой мышцы в этом случае минимальна. При увеличении силы раздражения амплитуда сокращения мышцы будет возрастать до тех пор, пока эта сила не окажется пороговой и для наименее возбудимых волокон. Дальнейшее увеличение силы раздражения не вызывает нарастания амплитуды сокращения.
О с н а щ е н и е: универсальный штатив с вертикальным миографом, кимограф, ключ для замыкания цепи, УЭС-1, бумага, чернила, эфир. Работу проводят на лягушке.
С о д е р ж а н и е р а б о т ы. Приготовить нервно-мышечный препарат, укрепить его в миографе. Включить стимулятор, ключ поставить в положение "Мышца" и, постепенно вращая ручку плавной регулировки, определить порог раздражения. Зарегистрировать на миографе минимальное сокращение мышцы. Затем последовательно увеличивать силу раздражения до тех пор, пока в ответ на каждый импульс наблюдается увеличение амплитуды мышечного сокращения. Отметить момент, когда, несмотря на дальнейшее увеличение силы тока, амплитуда сокращений перестанет увеличиваться.
О ф о р м л е н и е п р о т о к о л а. Вклеить полученную кимограмму и сделать на ней обозначения
Н.Е. Введенский (1886) на нервно-мышечном препарате установил, нерв, мионевральные синапсы и мышца по-разному реагируют на сверхпороговые раздражения различной частоты. Так, оказалось, что нерв способен воспроизводить наиболее широкий диапазон частотных раздражений, средний воспроизводится мышцей и наименьший - синапсами. Отсюда был сделан вывод, что разные структуры имеют неодинаковую функциональную подвижность или лабильность (от лат.labilis- скользящий, неустойчивый).
Лабильность -это скорость протекания процессов возбуждения в возбудимых тканях. Н.Е Введенский считал мерой лабильности максимальную частоту циклов возбуждения, воспроизводимых в единицу времени (секунду). Таким образом, зная величину лабильности, можно определить полную длительность одного цикла возбуждения.
Лабильность находится в прямой зависимости от фазы абсолютной рефрактерности: чем она короче, тем больше лабильность и наоборот. Поэтому лабильность любой возбудимой ткани можно высчитать, зная величину абсолютной рефрактерности. Так, в нервах она длится 0,001 сек, а отсюда огромная лабильность нерва - 1000 имп/сек. В скелетных мышцах рефрактерная фаза составляет 0,004-0,005 сек, а лабильность - 250-300 имп/сек. Очень низка лабильность сердца, всего 3 имп/сек, так как здесь очень продолжительная рефрактерная фаза (0,3 сек). Следовательно, фаза абсолютной рефрактерности ограничивает лабильность ткани.
Однако, доказано, что при частоте раздражения с интервалом абсолютной рефрактерности возникает лишь 1 ПД, ткань реагирует лишь на первый стимул, а на последующие - не отвечает. Это связано с тем, что сверхчастые раздражители удлиняют рефрактерность в ткани и развивается катодическая депрессия (пессимум) и восстановление мембранного потенциала после первого ПД не происходит. Поэтому, для получения максимальной частоты ритмические раздражители можно наносить с интервалом, превышающим рефрактерную фазу приблизительно в 2 раза. Поэтому, максимальная частота возбуждений в нервах составляет не 1000, а 500 имп/сек; в мышцах 100-125 имп/сек. Отсюда ясно, что максимальная частота возбуждения представляет крайнюю форму активности и может воспроизводится лишь в особых условиях и весьма короткое время. Следовательно, максимальнымназывается ритм, который генерирует ткань в экстремальных условиях и короткое время. Даже возбуждение с частотой в 2 фазы абсолютной рефрактерности оказывается слабым, так как он формируется в фазу относительной рефрактерности, когда возбудимость полностью еще не восстановлена.
Л.В. латманизова, ученица Ухтомского, обнаружила, что в естественных условиях возбудимые системы функционируют с намного меньшей частотой, чем максимальная. Но эта частота воспроизводится длительно и стойко, без утомления. Тот ритм возбуждения, в котором ткань функционирует длительное время и без утомления, называетсяоптимальным ритмом.Возникновение оптимального ритма связано с тем, что последующий раздражитель попадает в фазу супернормальной возбудимости, экзальтации, что благоприятствует возникновению возбуждения, и при этом может возникать максимальное сокращение мышцы. Частота, которая вызывает максимальный сократительный эффект, была названа Введенскимоптимальным ритмомраздражения, а сокращение -оптимальным. Этот ритм обычно наблюдается и в естественных условиях, возникая в период повышенной возбудимости ткани.
Максимальный и оптимальный ритмы связаны между собой математической зависимостью. Оптимальный ритм приблизительно в 5-10 раз реже, чем максимальный. Так для нерва максимальная частота составляет 1000 имп/сек, а оптимальная - 50-100 имп/сек. Для скелетных мышц они соответственно равны 250 и 50 имп/сек.
Таким образом, различают две частотные характеристики тканей. Максимальный ритм является мерой лабильности и проявляется в экстренных условиях короткое время. Вторая частотная характеристика - оптимальный ритм используется тканями в естественных условиях. Максимальным ритмом пользуются для выяснения и сравнения лабильности, а оптимальным - для характеристики изменений в функции данной возбудимой системы.
В процессе филогенеза лабильность тканей увеличилась. Лабильность ЦНС позвоночных несравненно выше, чем у беспозвоночных. Наиболее она высока в нервах, обеспечивающих срочную связь в организме. В эволюции физиологии есть яркие примеры изменения лабильности определенных органов в зависимости от их роли в жизни животного. Так, у млекопитающих ресничная (цилиарная) мышца является гладкомышечной структурой, поэтому изменение формы хрусталика и аккомодация глаза осуществляется очень медленно. У хищных птиц эта мышца - поперечно-полосатая и с высокой быстротой меняет форму хрусталика, обеспечивая ясное видение при падении птиц на добычу.
Лабильность меняется в процессе онтогенеза: при рождении она мала, в 20-30 лет достигает максимума, а после 60 лет быстро снижается, что соответствующим образом меняет работоспособность.
Лабильность неодинакова не только у разных тканей, но и у разных структурных единиц одной и той же ткани. Более того, даже у клетки лабильность непостоянна и определяется ее функциональным состоянием. Она может изменяться в процессе длительного воздействия раздражителей. Это, в частности, подтверждается способностью ткани повышать свою функциональную подвижность в процессе жизнедеятельности. При этом у ткани возникают новые свойства, и она приобретает способность воспроизводить более высокий ритм раздражения. Это явление, наблюдаемое в тканях, исследовал ученик и последователь введенского, академик А.А.ухтомский, и назвалпроцессом усвоения ритма.Повышением лабильности объясняется втягивание в работу, усвоение все большего ритма, а в результате укорочения рефрактерной фазы она способна возрасти в 2 раза. Это связано с ускорением процессов выкачивания ионов натрия из цитоплазмы и более быстрым восстановлением мембранного потенциала.
Теория по нормальной физиологии на тему: Синапс. Физиология мышечных волокон. В данной статье рассматривается механизм синаптической передачи, НМС…
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Синапс — это специфическое место контакта двух возбудимых систем (клеток) для передачи возбуждения.
По способу передачи сигналов:
- механические,
- химические,
- электрические.
По виду медиатора: холинэргические и др.
Нервно-мышечный синапс (НМС) — химический, передача с помощью медиатора ацетилхолина.
Синонимы к слову НМС:
- Нервно-мышечное соединение;
- Моторная концевая пластинка.

Аксоны нервных клеток на своих окончаниях теряют миелиновую оболочку , ветвятся, и концевые веточки аксона утолщаются. Это пресинаптическая терминаль или бляшка или пуговка , которая погружается в углубление на поверхности мышечного волокна.
Покрывающая концевую веточку аксона поверхностная мембрана называется пресинаптической мембраной , т.е. это мембрана, покрывающая поверхность синаптической бляшки (терминали аксона).
Мембрана, покрывающая мышечное волокно в области синапса, называется постсинаптической мембраной , или концевой пластинкой. Она имеет извитую структуру, образуя многочисленные складки, уходящие вглубь мышечного волокна, за счет чего увеличивается площадь контакта.

На постсинаптической мембране находятся белковые структуры — рецепторы , способные связывать медиатор . В одном синапсе количество рецепторов достигает 10-20 млн.
Между пре- и постсинаптическими мембранами находится синаптическая щель , размеры ее в среднем 50 нм, она открывается в межклеточное пространство и заполнена межклеточной жидкостью .
В синаптической щели находится мукополисахаридное плотное вещество в виде полосок, мостиков и содержится фермент ацетилхолинэстераза .
В пресинаптической терминали находится большое количество пузырьков или везикул , заполненных медиатором — химическим веществом посредником, осуществляющим передачу возбуждения.
В нервно-мышечном синапсе медиатор — ацетилхолин (АХ).

АХ синтезируется из холина и уксусной кислоты (ацетил-коэнзима А) с помощью фермента холинэстеразы. Эти вещества перемещаются из тела нейрона по аксону к пресинаптической мембране. Здесь в пузырьках происходит окончательное образование АХ.
3 фракции медиатора:
- Первая фракция — доступная — располагается рядом с пресинаптической мембраной.
- Вторая фракция — депонированная — располагается над первой фракцией.
- Третья фракция — диффузно рассеянная — наиболее удаленная от пресинаптической мембраны.
Механизм синаптической передачи



Ионы Ca вызывают образование специального белкового комплекса , который включает в себя везикулу и структуры, расположенные непосредственно около пресинаптической мембраны.
Они связаны между собой так называемыми белками экзоцитоза.
Часть белков расположена на везикулах (синапсин, синаптотагмин, синаптобревин), а часть — на пресинаптической мембране (синтаксин, синапсо-ассоциированный белок). Данный комплекс получил название секретосома .



Излитию содержимого пузырька в щель способствует белок синаптопорин , формирующий канал, по которому идет выброс медиатора.
Квант медиатора — количество молекул, содержащихся в одной везикуле.
На 1 ПД выбрасывается 100 квантов АХ.





На постсинаптической мембране возникает потенциал концевой пластинки (ПКП). Он является аналогом локального ответа (ЛО).
Потенциал действия на постсинаптической мембране не возникает ! Он формируется на соседней мембране мышечного волокна.
- связывание с рецептором,
- разрушение ферментов (ацетилхолинэстеразой),
- обратное поглощение в пресинаптическую мембрану,
- вымывание из щели и фагоцитоз.
События в синапсе :
- ПД приходит к терминали аксона;
- Он деполяризует пресинаптическую мембрану;
- Ca2+ входит в терминаль, что приводит к выделению АХ;
- В синаптическую щель выделяется медиатор АХ;
- Он диффундирует в щель и связывается с рецепторами постсинаптической мембраны;
- Меняется проницаемость постсинаптической мембраны для ионов Na+;
- Ионы Na+ проникают в постсинаптическую мембрану и уменьшают ее заряд — возникает потенциал концевой пластинки (ПКП) .
На самой постсинаптической мембране ПД возникнуть не может, так как здесь отсутствуют потенциалзависимые каналы, они являются хемозависимыми!
- ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и его распространению по мышечному волокну (около 5 м/с).
Достигнув пороговой величины, то есть КУД, ПКП возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна за счет местных круговых токов.

Особенности проведения возбуждения в нервно-мышечном синапсе
Одностороннее проведение возбуждения — только в направлении от пресинаптического окончания к постсинаптической мембране.
Суммация возбуждения соседних постсинаптических мембран.
Синаптическая задержка — замедление в проведении импульса от нейрона к мышце составляет 0,5-1 мс. Это время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, взаимодействие с рецептором, формирование ПКП, их суммацию.
Низкая лабильность — она составляет 100-150 имп/с для сигнала, что в 5-6 раз ниже лабильности нервного волокна.
Чувствительность к действию лекарственных веществ, ядов, БАВ, выполняющих роль медиатора.
Утомляемость химических синапсов — выражается в ухудшении проводимости вплоть до блокады в синапсе при длительном функционировании синапса. Главная причина утомляемости — исчерпание запасов медиатора в пресинаптическом окончании.
Законы проведения возбуждения по нервам:
- Закон функциональной целостности нерва.
- Закон изолированного проведения возбуждения.
- Закон двустороннего проведения возбуждения.
В зависимости от скорости проведения возбуждения нервные волокна подразделяются на 3 группы: A, B, C. В группе A выделяют 4 подгруппы: альфа, бетта, гамма и сигма.
Физиология мышечных волокон
- скелетная (40-50% массы тела),
- сердечная (менее 1%),
- гладкая (8-9%).
Физиологические свойства скелетных мышц:
- Возбудимость — способность отвечать на действие раздражителя возбуждением.
- Проводимость — способность проводить возбуждение из места его возникновения к другим участкам мышцы.
- Лабильность — способность мышцы сокращаться в соответствии с частотой действия раздражителя (200-300 Гц для скелетной мышцы).
- Сократимость — для мышцы является специфическим свойством — это способность мышцы изменять длину или напряжение в ответ на действие раздражителя.
Физические свойства скелетных мышц:
- Растяжимость — способность мышцы изменять длину под действием растягивающей силы.
- Эластичность — способность мышцы восстанавливать первоначальную длину или форму после прекращения действия растягивающей силы.
- Силы мышц — способность мышцы поднять максимальный груз.
- Способность мышцы совершать работу.
Режимы сокращения:
- Изотонический,
- Изометрический,
- Ауксотонический.
Изотонический режим — сокращение мышцы происходит с изменением ее длины без изменения напряжения (тонуса) (напр.: сокращение мышц языка).
Изометрический режим — длина постоянная, увеличивается степень мышечного напряжения (тонуса) (напр.: при поднятии непосильного груза).
Ауксотонический режим — одновременно изменяется длина и напряжение мышцы (характерен для обычных двигательных актов).
Механизм сокращения поперечно-полосатых мышц
Любая скелетная мышца состоит из мышечных волокон, которые, в свою очередь, состоят из множества тонких нитей — миофибрилл , расположенных продольно. Каждая миофибрилла состоит из протофибрилл — нитей сократительных белков: миозина (миозиновая протофибрилла), актина (актиновая протофибрилла).
Кроме сократительных белков в миофибрилле имеются два регуляторных белка: тропомиозин и тропонин .
Миозиновые волокна соединены в толстый пучок, от которого в торону актиновых нитей отходят поперечные мостики. У каждого мостика выделяют шейку и головку.
Нить актина располагается в виде 2 скрученных ниток бус. На ней имеются актиновые центры.

Тропомиозин в виде спиралей оплетает поверхность актина, закрывая в покое ее центры. Одна молекула тропомиозина контактирует с 7 молекулами актина.
Тропонин образует утолщение на конце каждой нити тропомиозина.
Под влиянием возникшего в мышечном волокне ПД из саркоплазматического ретикулума (СПР — депо Ca2+) высвобождаются ионы Ca. Кальций связывается с тропонином, который смещает тропомиозиновый стержень, что приводит к открытию актиновых центров.

В результате, к актиновым центрам присоединяются головки поперечных миозиновых мостиков.
Процесс расслабления происходит в обратной последовательности с использованием энергии АТФ за счет функционирования кальциевого насоса.
При отсутствии повторного импульса ионы Ca не поступают из СПР. В результате отсутствия Ca-тропонинового комплекса, тропомиозин возвращается на свое прежнее место, блокируя актиновые центры актина. Актиновые протофибриллы легко скользят в обратном направлении благодаря эластичности мышцы, и мышца удлиняется (расслабляется).
Гладкие мышцы
Гладкие мышцы — это мышцы, формирующие слой стенок полых внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток без поперечной исчерченности за счет хаотичного расположения миофибрилл.
Особенности гладких мышц:
- Иннервируются волокнами вегетативной нервной системы (ВНС);
- Обладают низкой возбудимостью:
- Обладают низкой величиной МП (мембранного потенциала) — -50 — -60 мВ из-за более высокой проницаемости для ионов Na+
- ПД (потенциал действия) отличается меньшей амплитудой и большей длительностью. Он формируется в основном за счет ионов Ca2+
- Медленная проводимость:
Клетки в гладких мышцах функционально связаны между собой посредством щелевидных контактов — нексусов, которые имеют низкое электрическое сопротивление. За счет этих контактов ПД распространяется с одного мышечного волокна на другое, охватывая большие мышечные пласты, и в реакцию вовлекается вся мышца.
Гладкие мышцы способны осуществлять относительно медленные ритмические и длительные тонические сокращения.
Медленные ритмические сокращения обеспечивают перемещение содержимого органа из одной области в другую.
Длительные тонические сокращения, особенно сфинктеров полых органов, препятствуют выходу из них содержимого.
Это способность сохранять приданную им при растяжении или деформации форму. Благодаря пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии.
Особенность гладких мышц, отличающая их от скелетных. Благодаря автоматии гладкие мышцы могут сокращаться в условиях отсутствия иннервации . Важную роль в этом играет растяжение.
Растяжение является адекватным раздражителем для гладкой мускулатуры. Сильное и резкое растяжение гладких мышц вызывает их сокращение.
Сравнительная характеристика скелетных и гладких мышц:
Уровень Л. характеризует скорость возникновения и компенсации возбуждения в любых клетках и уровень их функц, состояния. Можно измерять Л. мембран, клеток, органов, причем в системе Л. определяется через Л. ее наименее лабильного звена. Так, если Л. моторного аксона лягушки ок. 500 сокращений в 1 сек., Л. мышечного волокна ок. 200, а Л. синапса ок. 100, то Л. нервно-мышечной системы будет ок. 100 сокращений в 1 сек. Вследствие различной Л. различных участков мембраны нейрона его Л., измеряемая с различных синаптических входов, будет различной. Л. различных рефлекторных дуг неодинакова и в процессе деятельности может изменяться по ходу реакции как в сторону повышения, так и в сторону снижения. При усилении активного ионного транспорта и при гиперполяризации мембран Л. возрастает, при деполяризации мембран Л. снижается с возникновением сначала возбуждения, а затем пессимального (запредельного) торможения. Возбуждение и ритмическая активность возникают при среднем, оптимальном уровне Л. и поляризации нейронов. Л. измеряется при регистрации вызванной биоэлектрической активности клеток, тканей и органов в условиях макроэлектродного (суммарного) и микроэлектродного (вне- и внутриклеточного) отведения в лаб. и клин, практике для оценки функц. состояния различных органов (см. Биоэлектрические потенциалы, Электрокардиография, Электромиография, Электроэнцефалография). С помощью различных функц, проб, в частности ритмической адекватной стимуляции органов чувств, определяется Л. соответствующих корковых полей и даже отдельных нейронов (реакция следования ритму).
Нарушение лабильности
Снижение Лабильности, проявляющееся в виде уменьшения или исчезновения ответов на второй и последующие стимулы при ритмическом раздражении (максимальной адекватной частоты), свидетельствует о нарушении способности структуры восстанавливать свои энергетические и пластические запасы, необходимые для осуществления полноценной реакции. Однако и увеличение способности структуры реагировать на высокочастотную стимуляцию далеко не всегда свидетельствует о ее оптимальном состоянии; чаще такое явление наблюдается при экзальтации и может говорить о так наз. раздражительной слабости, или истериозисе. В таких случаях быстро наступает утомление или истощение резервов реагирующей структуры.
При значительных нагрузках или в неблагоприятных условиях часть возбудимых систем обычно обратимо повреждается и Л. этих структур, естественно, снижается. В условиях обратимого нарушения проведения в нервно-мышечных синапсах (частичное блокирование секреции медиатора) мышца способна отвечать на стимуляцию нерва с большей частотой, чем в норме. Одновременно может возрастать амплитуда ответов мышцы. Это объясняется тем, что синхронная волна возбуждения, достигая нервно-мышечных синапсов, застает их в различном функц, состоянии — часть синапсов передает возбуждение на мышцу, другая часть (обратимо поврежденные синапсы) еще не восстановила свой исходный функц, уровень. Импульс возбуждения может пройти, следовательно. лишь через синапсы первой группы. Следующий импульс застает их уже в состоянии невозбудимости (рефрактерности). Обратимо же поврежденные синапсы к моменту прихода второго импульса могут восстановить свою возбудимость благодаря влиянию первого импульса, который хотя и не вызывает передачу возбуждения, но повышает Л. этих синапсов. Именно в этом лежит причина того, что при обратимом повреждении части синапсов повышается способность нервно-мышечного аппарата давать реакцию за счет несинхронного включения в реакцию различных групп мышечных волокон. В условиях же нормы в реакцию уже на первый стимул вступают почти все мышечные волокна, поскольку они и их синаптический аппарат находятся практически в одинаковом функциональном состоянии. Отсюда следует, что так наз. пессимальная реакция мышцы (в норме) в виде крутого падения амплитуды ответов на первые стимулы при высокочастотном сверхпороговом раздражении нерва не является показателем патологии. Напротив, такая реакция говорит о том, что все группы мышечных волокон обладают высокой и одинаковой реактивностью.
Т. о., обычные критерии Лабильности не всегда адекватно отражают функц, состояние ткани, особенно состояние сложных образований, напр, нервных центров, обладающих специальными механизмами регулирования функц, состояния составляющих их элементов (см. Нервная система). Используемый для изучения Л. тест усвоения ритма должен применяться с известной осторожностью, т. к. нельзя отождествлять механизмы усвоения ритма в элементарной физиол. структуре и в сложной системе, обладающей специальным механизмом регулирования активности и находящейся под интегративным контролем со стороны других систем. Применяемые критерии Лабильности пригодны лишь для характеристики рабочих возможностей и скорости восстановления реактивности элементарных физиологических структур.
Библиография: Введенский Н. Е. Соотношения между ритмическими процессами и функциональной активностью возбужденного нервно-мышечного аппарата, Полн. собр. соч., т. 3, с. 84, Л., 1952; он же, Возбуждение, торможение и наркоз, там же, т. 4, с. 9, Л., 1953; Голиков Н. В. Физиологическая лабильность и ее изменения при основных нервных процессах, Л., 1950, библиогр.; Крыжановский Г. Н., Поздняков О. М. и Полгар А. А. Патология синаптического аппарата мышцы, М., 1974, библиогр.; Ухтомский А. А. Собрание сочинений, т. 2, с. 33 и др., Л., 1951, т. 6, с. 168, Л., 1962.
Н. В. Голиков; Г. Н. Крыжановский (пат. физ.).
Читайте также:
