
Наибольшая скорость проведения нервного импульса в аксоне
В 1830 г. один из крупнейших физиологов XIX века Иоганн Мюллер заявил, что скорость распространения ПД измерить невозможно. По его мнению, поскольку ПД – это электрический импульс, он должен проводиться со скоростью, примерно равной скорости света (3–10 10 см/с); учитывая небольшие размеры биологических объектов, даже с помощью лучших инструментов того времени измерить такую скорость было невозможно.
Спустя 15 лет один из студентов Мюллера Герман фон Гельмгольц с помощью простого и изящного эксперимента, который легко воспроизвести на студенческом лабораторном практикуме (рис. 6–8), измерил скорость распространения импульсов в нерве лягушки. В своих опытах Гельмгольц раздражал нерв в двух участках, отстоящих друг от друга на 3 см, и измерял время от момента подачи стимула до максимума мышечного сокращения. Предположим, что при раздражении дистального (расположенного ближе к мышцам) участка это время уменьшается на 1 мс. Тогда скорость распространения импульсов V равна
| V | = | d | / | t | = | 3 см | / | 1 мс | = | · | 10 3 | см/с |
Эта величина оказалась на семь порядков меньше, чем скорость распространения электрического тока в медном проводнике или в растворе электролита. Отсюда Гельмгольц сделал совершенно правильный вывод, что проведение нервного импульса –это более сложный процесс, чем простое продольное распространение тока в нервном волокне.
Скорость распространения импульсов в различных аксонах варьирует от 120 м/с (в некоторых крупных волокнах) до нескольких сантиметров в секунду (в очень тонких аксонах). Эти различия между скоростью проведения в разных волокнах иллюстрируют табл. 6–1 и рис. 6–9.
Скорость распространения импульса в значительной степени зависит от того, как быстро участок мембраны, расположенный на определенном расстоянии от места подачи стимула, деполяризуется местными токами до порогового уровня. Чем выше постоянная длины волокна, тем дальше могут распространяться эти токи, тем быстрее происходит деполяризация мембраны впереди от места возбуждения и, следовательно, тем выше скорость распространения импульса. Влияние постоянной длины на эту скорость можно продемонстрировать, если поместить аксон в масло или в воздух. При этом на поверхности аксона остается лишь тонкая пленка солевого раствора, и постоянная длины уменьшается из–за увеличения наружного продольного сопротивления [в уравнении (6–2) –r0]. В этих условиях скорость проведения возбуждения будет ниже чем при погружении аксона в солевой раствор.
Таблица 6–1. Классификация нервных волокон лягушки по их диаметру и скорости проведения возбуждения (Erlanger, Gasser, 1937)
| Группа волокон | Диаметр, мкм | Скорость, м/с |
| A α | 18,5 | |
| β | 14,0 | |
| γ | 11,0 | |
| B | — | 4,2 |
| C | 2,5 | 0,4 –0,5 |
В процессе эволюции живые организмы выработали два способа увеличения постоянной длины аксона и тем самым–скорости распространения импульса. Один из них (типичным примером могут быть гигантские аксоны кальмаров, членистоногих кольчатых червей, костистых рыб) – это увеличение диаметра аксона, т. е. уменьшение внутреннего продольного сопротивления [в уравнении (6–2) – ri] Подробнее этот вопрос рассмотрен в дополнении 6–2. Гигантские аксоны развились в процессе эволюции у некоторых видов животных для того, чтобы обеспечивать быструю синхронную активацию двигательных рефлексов, например движений мантии у кальмара и рефлекса отдергивания либо избегания у некоторых членистоногих (раков, тараканов) и кольчатых червей (например, земляных).
Сальтаторное проведение
Второй способ увеличить скорость проведения нервных импульсов, реализовавшийся только у позвоночных животных, состоит в изоляции участков аксона с помощью миелиновой оболочки. При этом постоянная длины соответствующих участков значительно увеличивается, и тем самым существенно облегчается проведение тока в продольном направлении. По мере развития животного миелин откладывается вокруг периферических и центральных аксонов глиальными клетками, расположенными вблизи этих аксонов. В результате вокруг волокон образуется плотная многослойная оболочка из клеточных мембран. К клеткам, синтезирующим миелин, относятся шванновские клетки (в области периферических нервов) и олигодендроциты (рис. 6–10) (в ЦНС). На поперечных срезах миелиновой оболочки видны периодически повторяющиеся промежутки в 12 нм, образующиеся в результате наслоения мембран глиальных клеток. С образованием каждого нового слоя поперечное сопротивление оболочки увеличивается. Поскольку слоев в этой оболочке много, ее емкость гораздо ниже, чем у одиночной мембраны. Многослойная миелиновая оболочка периодически прерывается (так называемые перехваты Ранвье>, и на этих небольших участках возбудимая мембрана аксона контактирует с внеклеточной средой. Между перехватами Ранвье миелиновая оболочка тесно прилегает к мембране аксона, практически вытесняя внеклеточную среду. Кроме того, участки мембраны аксона между перехватами Ранвье, по–видимому, не содержат натриевых каналов.
Благодаря изолирующим свойствам миелиновой оболочки постоянная длины аксона резко возрастает: наличие этой оболочки оказывает такой же эффект, как и увеличение rм [уравнение (6–2)]. Из–за высокого сопротивления миелиновой оболочки местные токи, текущие впереди от волны возбуждения, выходят из аксона почти исключительно в области перехватов Ранвье. Кроме того, поскольку емкость толстой миелиновой оболочки мала, на перезарядку этой емкости в участках между перехватами расходуется лишь очень небольшой ток. Благодаря этим особенностям ПД, возникающий в каком–либо перехвате, электротонически деполяризует лишь мембрану, расположенную в области следующего перехвата, и поэтому импульсы в таких аксонах не распространяются по всей их длине, как в немиелинизированных нервных волокнах (например, в аксоне кальмара). Они возникают лишь в небольших участках мембраны –перехватах Ранвье. Все это обусловливает сальтаторное (скачкообразное) проведение, при котором импульсы распространяются прерывисто от перехвата к перехвату (рис. 6–11). Скорость распространения. ПД при этом резко увеличивается, поскольку электротоническое проведение местных токов между перехватами осуществляется очень быстро. Таким образом, у позвоночных животных Природа решила проблему быстрого распространения нервных импульсов, не прибегая к созданию таких громоздких структур, как гигантские аксоны.
Нервная система человека выступает своеобразным координатором в нашем организме. Она передаёт команды от мозга мускулатуре, органам, тканям и обрабатывает сигналы, идущие от них. В качестве своеобразного носителя данных используется нервный импульс. Что он собой представляет? С какой скоростью работает? На эти, а также на ряд других вопросов можно будет найти ответ в этой статье.
Чем является нервный импульс?
Исследование строения и работы
Впервые прохождение нервного импульса было продемонстрировано немецкими учеными Э. Герингом и Г. Гельмгольцем на примере лягушки. Тогда же и было установлено, что биоэлектрический сигнал распространяется с указанной ранее скоростью. Вообще, такое является возможным благодаря особенному построению нервных волокон. В некотором роде они напоминают электрический кабель. Так, если проводить параллели с ним, то проводниками являются аксоны, а изоляторами – их миелиновые оболочки (они являют собой мембрану шванновской клетки, которая намотана в несколько слоев). Причем скорость нервного импульса зависит в первую очередь от диаметра волокон. Вторым по важности считается качество электрической изоляции. Кстати, в качестве материала организмом используется липопротеид миелин, который обладает свойствами диэлектрика. При прочих равных условиях, чем больше будет его слой, тем быстрее будут проходить нервные импульсы. Даже на данный момент нельзя сказать, что эта система полноценно исследована. Многое, что относится к нервам и импульсам, ещё остаётся загадкой и предметом исследования.
Особенности строения и функционирования
Где они создаются?
Типы клеток
- Рецепторные (чувствительные). Ими кодируются и превращаются в нервные импульсы все температурные, химические, звуковые, механические и световые раздражители.
- Вставочные (также называются кондукторными или замыкательными). Они служат для того, чтобы перерабатывать и переключать импульсы. Наибольшее их число находится в головном и спинном мозге человека.
- Эффекторные (двигательные). Они получают команды от центральной нервной системы на то, чтобы были совершены определённые действия (при ярком солнце закрыть рукой глаза и так далее).
Каждый нейрон имеет тело клетки и отросток. Путь нервного импульса по телу начинается именно с последнего. Отростки бывают двух типов:
- Дендриты. На них возложена функция восприятия раздражения расположенных на них рецепторов.
- Аксоны. Благодаря им нервные импульсы передаются от клеток к рабочему органу.
Интересный аспект деятельности
О потенциале действия
Как всё работает в мозгу?
Работа нейромедиаторов
Когда они передают нервные импульсы, то существует несколько вариантов, что произойдёт с ними:
- Они будут диффундированы.
- Подвергнутся химическому расщеплению.
- Вернутся назад в свои пузырьки (это называется обратным захватом).
В конце 20-го века сделали поразительное открытие. Ученые узнали, что лекарства, что влияют на нейромедиаторы (а также их выброс и обратный захват), могут изменять психическое состояние человека коренным образом. Так, к примеру, ряд антидепрессантов вроде "Прозака" блокируют обратный захват серотонина. Есть определённые причины считать, что в болезни Паркинсона виноват дефицит в головном мозге нейромедиатора дофамина.
Если кратко, то они могут работать с тысячами нейромедиаторов, которые посылаются их соседями. Детали относительно обработки и интеграции данного типа импульсов нам почти не известны. Хотя над этим работает много исследовательских групп. На данный момент получилось узнать, что все полученные импульсы интегрируются, а нейрон выносит решение – необходимо ли поддерживать потенциал действия и передавать их дальше. На этом фундаментальном процессе базируется функционирование головного мозга человека. Ну что ж, тогда это неудивительно, что мы не знаем ответа на эту загадку.
Некоторые теоретические особенности
Где же создаются нервные импульсы?
Откуда они начинают свой путь? Ответ на этот вопрос может дать любой студент, который прилежно изучал физиологию возбуждения. Есть четыре варианта:
- Рецепторное окончание дендрита. Если оно есть (что не факт), то возможным является наличие адекватного раздражителя, что создаст сначала генераторный потенциал, а потом уже и нервный импульс. Подобным образом работают болевые рецепторы.
- Мембрана возбуждающего синапса. Как правило, такое возможно только при наличии сильного раздражения или их суммирования.
- Триггерная зона дентрида. В этом случае локальные возбуждающие постсинаптические потенциалы формируются как ответ на раздражитель. Если первый перехват Ранвье миелинизирован, то они на нём суммируются. Благодаря наличию там участка мембраны, которая обладает повышенной чувствительностью, здесь возникает нервный импульс.
- Аксонный холмик. Так называют место, где начинается аксон. Холмик – это наиболее частый создать импульсов на нейроне. Во всех остальных местах, которые рассматривались ранее, их возникновение гораздо менее вероятное. Это происходит из-за того, что здесь мембрана имеет повышенную чувствительность, а также пониженный критический уровень деполяризации. Поэтому, когда начинается суммирование многочисленных возбуждающих постсинаптических потенциалов, то раньше всего на них реагирует холмик.
Пример распространяющегося возбуждения
Вспомните сводки из новостей прошлого лета (также это скоро можно будет услышать опять). Пожар распространяется! При этом деревья и кустарники, которые горят, остаются на своих местах. А вот фронт огня идёт всё дальше от места, где был очаг возгорания. Аналогичным образом работает нервная система.
Часто бывает необходимо успокоить начавшееся возбуждение нервной системы. Но это не так легко сделать, как и в случае с огнем. Для этого совершают искусственное вмешательство в работу нейрона (в лечебных целях) или используют различные физиологические средства. Это можно сравнить с заливанием пожара водой.
Генерация и распространение нервного импульса в организмах животных, возбуждение нервных и мышечных клеток — важнейшие мембранные явления. Мембранная теория возбуждения была сформулирована Бернштейном еще в 1902 г. [1]. Согласно его теории, возбуждение определяется электрохимическими процессами, локализованными в мембранах нервных и мышечных клеток. Эти процессы сводятся к перемещению малых ионов. Позднее Лазарев опубликовал монографию об ионной природе нервного возбуждения [2]. Исследования, проведенные Ходжки- ном, Катцем, Хаксли, Тасаки и другими учеными [3—6], раскрыли принципиальные механизмы генерации нервного импульса и его распространения (см. также [7—9]).
Нервное возбуждение распространяется по нервным волокнам— аксонам. Принято разделять нервную систему высших организмов на центральную и периферическую. Периферическая система содержит аксоны, служащие для передачи сигналов, а также ганглии вегетативной нервной системы. Аксоны являются коммуникациями для афферентных сообщений от органов чувств, направляемых в центральную систему, и для эфферентных сигналов, направляющихся от центральной системы к мышцам. Аксоны представляют собой отростки центрально расположенных клеток. Нервная система беспозвоночных имеет иное строение, но она также содержит коммуникационные аксоны.
Исследование генерации нервного импульса и его распространения в аксоне — старая и традиционная проблема биофизики/ Уже Гельмгольц измерял скорость распространения нервного возбуждения/В настоящее время решены важные физические задачи, относящиеся к функции аксона. Напротив, современное состояние науки позволяет лишь формально моделировать работу центральной нервной системы и мы еще далеки от понимания физической природы высших ее функций — памяти и мышления.
На рис. 4.1, заимствованном из [5], представлена схема строения нервной клетки, именуемой иначе нейроном. Нейрон полу
чает сообщения от многих других нейронов через их тонкие ответвления, образующие контакты — синапсы — с телом клетки и его короткими отростками — дендритами. Аксон имеет макроскопические протяженность и сечение. Длины аксонов в теле крупных животных достигают нескольких метров.\
Контакты с другими клетками образуются не только в синапсах.^ Боль - шая часть поверхности нейрона покрыта прилегающими к нему глиальными, или иіванновскими, клетками^ роль которых пока не выяснена. С другой стороны^из мембран шванновских клеток образуется миелиновая оболочка миелинизирован - ных аксонов, показанная ^схематически на рис. 4.1. ^Оболочка прерывается через каждые 1—2 мм длины аксона перехватами Ранвье, имеющими протяженность /Т около 1 мкм. В области перехватов мембрана аксона непосредственно контактирует с окружающей средойЛ^
Немиелинизированные во - ЛОКНа могут быть объеди - Рис. 4.1. Схема строения нервной клетки нены одной шванновской (двигательный нейрон).
К-ТТР. ТКПЙ чтп ППКЯ1ЯНП R гр - і тн
тел0 нейрона,1Ца — аксои1Смо — миелиновая клеткой, ЧТО UOKdJdHU В се
оболочка, перехват Ранвье^д-дендрит,
ЧЄНИИ на рис. 4.2. Погру - X*— ядро,^с —синапсыЛодн—окончание двига о г - г тельного нерва,\мв — мышечное волокно.
Женныи в шванНЬвскую 1
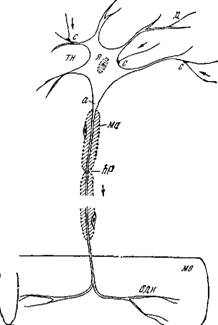
Клетку аксон создает глубокую инвагинацию мембраны шванновской клетки, причем образуется мезаксон — две шванновские мембраны, расположенные рядом^В правом нижнем углу схематически показана двойная мембрана, разделяющая цитоплазму шванновской клетки и цитоплазму аксона, аксоплазму [9].
Герен [10] установила, что миелиновая оболочка аксона образована многократным спиральным закручиванием мембраны шванновской клетки вокруг аксоиаГМиелин представляет собой, - т-а«им
ойразом,/многослойную (доЧїбО слоев) мембрану, строение которой наблюдается с помощью электронного микроскопа. Миелии защищает мембрану аксона от окружающей среды.
6 М. В. Волькеиштейн
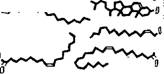
Структура миелина была детально изучена методом рентгенографии с разрешением 10 А [11]. Определены профили электронной плотности миелина седалищного нерва кролика и лягушки и миелина зрительного нерва кролика. На рнс. 4.3 показаны эти кривые для нервных волокон кролика и схема структуры миелина, соответствующая данным кривым. Миелин построен асимметрично, холестерин сосредоточен преимущественно на стороне
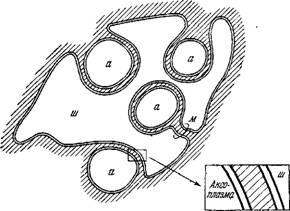
Рис. 4.2. Схема строения немиелинизированнного нервного волокна позвоночного.
А — аксон, м — мезаксон— шванновская клетка.
/Уже давно установлено, что нервы и мышцы способны генерировать электродвижущие силы, биопотенциалы. Электрическая активность клеток проявляется в форме коротких разрядов, каждый из которых длится около 1 мс. Современная экспериментальная техника позволяет усиливать эти сигналы и регистрировать их с помощью осциллографа или другим путем. Установлено, что активность нерва всегда сопровождается электрическими явлениями — и при внешнем возбуждении и при посылке сигнала мозгом в соответствующем аксоне наблюдаются электрические импульсы. На рис. 4.4 показаны импульсы в зрительном нервном волокні краба Limulus, вызванные вспышкой света длительностью 1 с [13]. Частота и характер последовательности импульсов, имеющие несомненное информационное значе
ние, зависят от интенсивности и спектрального состава света [14, 15] (см. гл. 7) .1 Величина и длительность отдельного импульса не зависят отТгрироды и силы раздражения."
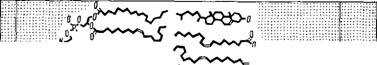

Внугрен^/7о/тярже=Стероид щ^Гибх/гя ^Стероид ^//плярная Внешний
Нийбеяоцт. группы^ +ягесткая цепь +жесткая белок
+еоЗа '— " цепь - цепь — —
Рис. 4.3. Схема миелиновой мембранной структуры.
Вверх* - профили электронной плотности для глазного (пунктир) н седалищного (сплошная
Лнння) нервов кролика.
Электрический сигнал, отвечающий отдельному импульсу, распространяющемуся вдоль аксона, называется потенциалом действия или спайком. Это — основная единица информации, передаваемой по нервному волокну.

Скорость распространения импульса по аксону равна по порядку величины 1—100 м/с, она меньше для немиелинизиро - ванных и больше для миелинизированных аксонов.


Рис. 4.4. Импульсы в зрительном нервном волокне Limulus, вызванные вспышкой света длительностью I с.
Цифры справа указывают относительную интенсивность вспышки. Период освещения указан разрывом верхней белой линии, отметка времени на нижней белой линии 0,2 с.
В табл. 4.1 [3] приведены некоторые данные. Проведение нервного возбуждения есть проведение электрического импульса. Несмотря на то, что аксоплазма представляет
Скорость проведения возбуждения в нервном н мышечном волокнах
У позвоночных животных в процессе эволюции сформировались аксоны, покрытые миелиновой оболочкой, что обеспечило большие скорости проведения нервных импульсов при значительно меньших диаметрах таких аксонов (см. табл. 11.3).
Строение миелинизированного аксона схематично представлено на рис. 11.6, а. Участки аксона длиной в 2-3 мм

На участках аксона, покрытых миелином, мембрана полностью изолирована и не имеет контакта с межклеточной жидкостью, содержащей ионы Na + , вследствие чего потенциал действия здесь сформироваться не может, даже если потенциал аксоплазмы превышает критический потенциал возбуждения Екр. Потенциал действия может сформироваться только в перехватах Ранвье, где мембрана имеет необходимый контакт с межклеточной жидкостью.
Допустим, что один из перехватов Ранвье возбужден, т.е. деполяризован до потенциала фтах. Тогда под действием разности потенциалов (фтах - ф0) между возбужденным и невозбужденным участками в аксоплазме и на наружной стороне мембраны возникают локальные токи, благодаря которым мембранный потенциал распространяется вдоль аксона как по электрическому кабелю и с большой скоростью. Но по мере удаления от возбужденного участка мембранный потенциал аксона экспоненциально уменьшается (рис. 11.6, б), как и любой местный потенциал [см. формулу (11.7)].
Когда этот затухающий мембранный потенциал достигнет ближайшего перехвата Ранвье и повысит там фм до Екр, в мембране этого перехвата открываются Ма + -каналы и генерируется полноценный потенциал действия, что приводит к увеличению мембранного потенциала до максимального значения фтах. Далее весь процесс повторяется, при этом потенциал действия последовательно генерируется лишь в перехватах Ранвье, а на миелинизированных участках аксона происходит его распространение с затуханием, но с большой скоростью, благодаря чему и достигается высокая скорость передачи возбуждения по такому нервному волокну.
Проведение нервного импульса в миелинизированном аксоне называют сапътаторным (скачкообразным), так как потенциал действия генерируется только в перехватах Ранвье и в процессе распространения как бы перескакивает с одного перехвата Ранвье на последующий. Как и в случае безмиели- новых нервных волокон, скорость распространения потенциала действия в миелинизированном аксоне возрастает с увеличением его диаметра, что хорошо видно из табл. 11.3.
Одной из причин высокой скорости проведения потенциала действия в миелинизированных аксонах является высокая плотность Na + -каналов в перехватах Ранвье, где она примерно в 100 раз выше, чем в безмиелиновых волокнах.
Высокая скорость проведения и малый диаметр миелинизированных аксонов обеспечили позвоночным животным возможность создания большого количества параллельных быстропроводящих нервных путей, что способствовало высокой надежности передачи информации по нервной системе.
Если входящий в клетку натриевый ток уменьшить, например, путем снижения концентрации Na + в растворе, окружающем аксон, либо путем инактивации (ухудшения проводимости) Na + -каналов (введением ядов либо воздействием местноанестезирующего препарата), то скорость проведения потенциала действия снижается и в конечном итоге проведение может быть блокировано.
Если с помощью определенных ядов (например, тетродо- токсина) заблокировать натриевые каналы в одном из перехватов Ранвье, то потенциал действия сформируется не на нем, а на следующем перехвате, так как амплитуда затухающего местного потенциала, распространяющегося за счет локальных токов (пунктирная кривая на рис. 11.6, б), еще сможет достичь на нем порогового потенциала возбуждения. Но если заблокировать подряд два или более перехватов Ранвье, то потенциал действия на последующих перехватах Ранвье сформироваться не сможет и проведение нервного импульса будет блокировано.

Нейрон состоит из одного аксона, тела и нескольких дендритов,
Аксон (греч. ἀξον — ось) — нервное волокно, длинная, вытянутая часть нервной клетки (нейрона), отросток или нейрит, элемент, который проводит электрические импульсы далеко от тела нейрона (сомы).
Строение нейрона Править
Нейрон состоит из одного аксона, тела и нескольких дендритов, в зависимости от числа которых нервные клетки делятся на униполярные, биполярные, мультиполярные. Передача нервного импульса происходит от дендритов (или от тела клетки) к аксону. Если аксон в нервной ткани соединяется с телом следующей нервной клетки, такой контакт называется аксо-соматическим, с дендритами — аксо-дендритический, с другим аксоном — аксо-аксональный (редкий тип соединения, встречается в ЦНС, участвует в обеспечении тормозных рефлексов).
В месте соединения аксона с телом нейрона находится аксонный холмик — именно здесь происходит преобразование постсинаптического потенциала нейрона в нервные импульсы, для чего требуется совместная работа натриевых, кальциевых и как минимум трех типов калиевых каналов.
В протоплазме аксона — аксоплазме — имеются тончайшие волоконца — нейрофибриллы, а также микротрубочки, митохондрии и агранулярная(гладкая)эндоплазматическая сеть. В зависимости от того, покрыты ли аксоны миелиновой (мякотной) оболочкой или лишены её, они образуют мякотные или безмякотные нервные волокна.
Концевые участки аксона — терминали — ветвятся и контактируют с другими нервными, мышечными или железистыми клетками. На конце аксона находится синаптическое окончание — концевой участок, контактирующий с клеткой-мишенью. Вместе с постсинаптической мембраной клетки-мишени синаптическое окончание образует синапс. Через синапсы передаётся возбуждение. [1]
Анатомия Править
Аксоны - в действительности первичные линии передачи сигналов нервной системы, и как связки они помогают составлять нервные волокна. Индивидуальные аксоны являются микроскопическими в диаметре (типично 1 мкм в сечении), но могут достигать нескольких метров. Самые длинные аксоны в человеческом теле, например, аксоны седалищного нерва, которые простираются от позвоночника к большому пальцу ноги. Эти волокна единственной ячейки седалищного нерва могут увеличиться до метра или еще длинее. [2]
Физиология Править
Физиология может быть описана Моделью Hodgkin-Huxley, распространённой на позвоночных животных в уравнениях Frankenhaeuser-Huxley. Периферийные волокна нерва могут быть классифицированы на основанные, на аксонально-скоростные проводимости, mylenation, размеры волокна и т.д. Например, есть медленное проведение unmyelinated С fibers волокна и более быстрое проведение myelinated Aδ fibers волокна. Более сложное математическое моделирование проводится сегодня. Есть несколько типов сенсорных — таких как motorfibers. Другие волокна, не упомянутые в матеоиале — например, волокна автономной нервной системы
Двигательная функция Править
В таблице паказаны моторные нейроны, которые имеют два вида волокон:
| Тип | Классификация | Диаметр | Миелин | Скорость проводимости | Связанные мускульные волокна |
|---|---|---|---|---|---|
| α | Aα | 13-20 мкм | Yes | 80-120 m/s | Extrafusal muscle fibers |
| γ | Aγ | 5-8 мкм | Yes | 4-24 m/s [3] [4] | Intrafusal muscle fibers |
Сенсорная функция Править
Различные сенсорные рецепторы возбуждаются различными типами волокон нерва. Proprioceptors возбуждены типом Ia, Ib и II сенсорными волокнами, механорецепторы — типом II и III сенсорными волокнами и типом nociceptors и thermoreceptors.
| Типы | Классификация | Диаметр | Миелин | Скорость проводимости | Связанные сенсорные рецепторы |
|---|---|---|---|---|---|
| Ia | Aα | 13-20 мкм | Yes | 80-120 m/s | Primary receptors of muscle spindle |
| Ib | Aα | 13-20 мкм | Yes | 80-120 m/s | Golgi tendon organ |
| II | Aβ | 6-12 мкм | Yes | 33-75 m/s | Secondary receptors of muscle spindle All cutaneous mechanoreceptors |
| III | Aδ | 1-5 мкм | Thin | 3-30 m/s | Free nerve endings of touch and pressure Nociceptors of neospinothalamic tract Cold thermoreceptors |
| IV | C | 0.2-1.5 мкм | No | 0.5-2.0 m/s | Nociceptors of paleospinothalamic tract Warmth receptors |
Автономная функция Править
Автономная нервная система имеет два вида периферийных волокон:
| Typы | Классификация | Диаметр | Миелин [5] | Скорость проводимости |
|---|---|---|---|---|
| preganglionic fibers | B | 1-5 мкм | Yes | 3-15 m/s |
| postganglionic fibers | C | 0.2-1.5 мкм | No | 0.5-2.0 m/s |
Рост и развитие аксона Править

Ячейки звонковые, ячейки указательного столба помогают в руководстве ростом аксона нейронала. Эти ячейки — типично другой, иногда незрелый, нейроны.
История Править
Часть первой внутриклеточной регистрации в нервной системе была сделана в конце 1930-ых учёными K. Капуста и H. Куртис. Алан Ходгкин и Эндрю Хакслей также использовали аксон гиганта кальмара (1939), и в 1952 они получили полное количественное описание действия ионного основания потенциала, введя формулировку Модели Hodgkin-Huxley. Ходгкину и Хакслей, были предтавлены совместно на паолучение Нобелевской премии по этой работе в 1963. Формулы, детализирующие аксональную проводимость были расширены на позвоночных животных в уравнениях Frankenhaeuser-Huxley. Erlanger и Gasser ранее развивали систему классификации для периферийного[5] волокна нерва, основанные на аксональной скорости проводимости, myelination, размере волокна и т.д. Даже и сейчас наше понимание биохимического процесса распространения действия потенциала продвинулось, и теперь он включает много деталей об индивидуальных каналах иона.
Рана Править
На серьёзном уровне, рана нерва может быть описана как neuropraxia, axonotmesis, или neurotmesis. Сотрясение мозга считают умеренной формой разбросанной аксональной раны [6] .
Читайте также:
