
Нерв строение тканевой состав
От нервных клеток, находящихся в головном и спинном мозгу, отходят отростки, которые и являются нервными волокнами, идущими к периферии. Нервные волокна собираются в пучки разной толщины. Такое скопление нервных волокон называется нервом.
Нервы осуществляют связь между центральной нервной системой и отдельными органами нашего тела. По нервам возбуждение идет либо из центральной нервной системы к рабочему органу, либо от разных участков нашего тела в центральную нервную систему.
Нервы делятся на две группы в зависимости от того, в каком направлении они проводят возбуждение.
Рис. Схема распространения возбуждения при раздражении нерва
Одна группа нервов проводит возбуждение из центральной нервной системы к рабочим органам. Они называются эфферентными (центробежными, или двигательными) нервами. Другая группа проводит возбуждение с разных участков нашего тела и от разных органов в центральную нервную систему. В отличие от предыдущей группы нервов они получили название афферентных (центростремительных, или чувствительных) нервов. Оба рода нервных волокон часто идут в одном стволе, поэтому большинство нервов являются смешанными.
СТРОЕНИЕ НЕРВА
Нервная система состоит из нервных клеток, которые называются нейронами. Нейрон состоит из тела нервной клетки и ее отростков. Различают два вида отростков: а) отростки короткие, ветвистые — дендриты, и б) очень длинный отросток, который тянется от центральной нервной системы до рабочего органа,— а к с о н, который участвует в формировании нервов.
Наконец, имеются еще и особые образования на окончаниях нервов- так называемые концевые аппараты, при помощи которых осуществляется связь нервного волокна с мышцей, железой или другими органами, или рецепторы — окончания центростремительных нервов, воспринимающие раздражение.
Короткие отростки — дендриты — осуществляют связь между отдельными нервными клетками и почти не выходят за пределы центральной нервной системы.
Аксон же тянется из головного или спинного мозга до рабочего органа. Нервы, которые мы встречаем в организме, состоят из аксонов, несущих возбуждение в центральную нервную систему или, наоборот, из центральной нервной системы.
Нормальное протекание обмена веществ во всех отростках нервной клетки связано с ее целостностью. В этом можно убедиться, если перерезать нервное волокно и тем самым нарушить его связь с телом клетки. Деятельность такого волокна нарушается, и та часть, которая отрезана от клетки, отмирает. Совершенно иные явления наблюдаются в той части волокна, которая осталась связанной с телом клетки. Эта часть продолжает жить, нормально функционирует, обмен веществ не нарушен. Более того, такой отрезок растет и через некоторое, время может дойти до мышцы, чем и восстановится целость, нерва. Этим объясняется наблюдающееся иногда восстановле ние движений парализованной конечности через определенный промежуток времени, если паралич был вызван поражением нерва.
Такой особенностью пользуются и хирурги, которые часто производят сшивание нервов с целью восстановления деятельности парализованного органа.
Нервная клетка возбуждается под влиянием тех волн возбуждения, которые поступают с периферии по центростремительным нервам. Однако многие нервные клетки могут возбуждаться даже без поступления импульсов с рецепторов. В этих клетках возбуждение может возникнуть под влиянием гуморальных воздействий. Примером может служить деятельность теплового центра, на функции которого влияет температура крови, и др.
СВОЙСТВА НЕРВНОГО ВОЛОКНА
Нервное волокно обладает возбудимостью и проводимостью. В этом можно убедиться, если нанести электрическое раздражение какому-либо участку нерва нервно-мышечного препарата. Почти тотчас после нанесения раздражения мышца сокращается. Сокращение мышцы стало возможным потому, что при раздражении в нерве возникло возбуждение, которое, пройдя по нерву, поступило к мышце и обусловило ее деятельность.
Для проведения возбуждения необходима анатомическая целость нервного волокна. Перерезка нерва делает невозможной передачу возбуждения. Возбуждение не проводится в случае перевязки, сдавливания или нарушения целости нерва любым иным способом. Однако не только анатомические, но и физиологические нарушения вызывают прекращение про ведения. Нерв может быть целым, но он не будет проводить возбуждения, так как его функ ции нарушены.
Нарушение проведения мож но наблюдать при охлаждении или нагревании нерва, прекращении его кровоснабжения, от равлении и т. д.
Проведение возбуждения по нерву подчиняется двух основ ным законам.
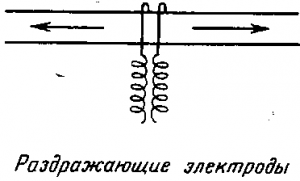
1. Закон двустороннего проведения. Нервное волокно обладает способностью проводить возбуждение по двум направлениям: центростремительно и центробежно. Независимо от того, какое это нервное волок но — центробежное или центростремительное, если ему нане сти раздражение, то возникшее возбуждение будет распространяться в обе стороны от места раздражения (рис.). Это свойство нервного волокна впервые открыл выдающийся русский ученый Р. И. Бабухин (1877).
2. Закон изолированного проведения. Периферический нерв состоит из большого числа отдельных нервных волокон, которые вместе идут в одном и том же нервном стволе. В нервном стволе одновременно могут проходить самые разнообразные центробежные и центростремительные нервные волокна. Однако возбуждение, которое передается по одному нервному волокну, не передается на соседние. Благодаря такому изолированному проведению возбуждения по нервному волокну возможны отдельные весьма тонкие движения человека. Художник может создавать свои полотна, музыкант — исполнять сложные музыкальные произведения, хирург — производить тончайшие операции потому, что каждое волокно изолированно передает импульс мышце, и тем самым центральная нервная система имеет возможность координировать мышечные сокращения. Если бы возбуждение могло переходить на другие волокна, стало бы невозможным отдельное мышечное сокращение, каждое возбуждение сопровождалось бы сокращением самых разнообразных мышц.
Статья на тему Строение нерва
Нервы — состоят из миелиновых и безмиелиновых волокон и соединительнотканных оболочек. В некоторых нервах встречаются одиночные нервные клетки и мелкие ганглии. На поперечном срезе нерва видны сечения осевых цилиндров нервных волокон и одевающие их глиальные оболочки. Между нервными волокнами в составе нервного ствола располагаются тонкие прослойки рыхлой волокнистой соединительной ткани —- эндоневрий. Пучки нервных волокон одеты периневрием. Периневрий состоит из чередующихся слоев плотно расположенных клеток и тонких фибрилл. Таких слоев в периневрий толстых нервов несколько. Фибриллы ориентированы вдоль нерва. Наружная оболочка нервного ствола — эпиневрий — представляет собой плотную волокнистую соединительную ткань, богатую фибробластами, макрофагами и жировыми клетками. Соединительнотканные оболочки нерва содержат кровеносные и лимфатические сосуды и нервные окончания.
Изменения в теле нейрона выражаются в его набухании, тигролизе — растворении глыбокхроматофильной субстанции, и в перемещении ядра на периферию тела клетки. Дегенеративные изменения в центральном отрезке ограничиваются распадом миелинового слоя и осевого цилиндра вблизи травмы.Если существует препятствие для врастания аксонов центрального отрезка нерва в тяжи нейролеммоцитов периферического отрезка, аксоны центрального отрезка растут беспорядочно и могут образовать клубок, называемый ампутационнойневромой. При ее раздражении возникает сильная боль.Поврежденные нервные волокна головного и спинного мозга не регенерируют. Возможно, регенерации нервных волокон в центральной нервной системе не происходит потому, что глиоциты без базальной мембраны лишены хемотаксических факторов, необходимых для проведения регенерирующих аксонов. Однако при малых травмах центральной нервной системы возможно частичное восстановление ее функций, обусловленное пластичностью нервной ткани.
43. Спинной мозг. Общая характеристика строения. Строение серого вещества: виды нейронов и их участие в образование рефлекторных дуг, типы глиоцитов.
Спинной мозг состоит из двух симметричных половин, отграниченных друг от друга спереди глубокой серединной щелью, а сзади — соединительнотканной перегородкой. Внутренняя часть органа темнее — это его серое вещество. На периферии спинного мозга располагается более светлое белое вещество. Выступы серого вещества называют рогами. Различают передние, или вентральные, задние, или дорсальные, и боковые, или латеральные, рога.
В процессе развития спинного мозга из нервной трубки образуются нейроны, группирующиеся в 10 слоях.
Серое вещество спинного мозга состоит из тел нейронов, безмиелиновых и тонких миелиновых волокон и нейроглии. Составной частью, являются мультиполярные нейроны.
Белое вещество представляет собой совокупность продольно ориентированных преимущественно миелиновых волокон. Пучки нервных волокон, осуществляющие связь между различными отделами нервной системы, называются проводящими путями спинного мозга.
Нейроциты. Среди нейронов можно выделить: корешковые клетки, внутренние клетки, и пучковые клетки.
В задних рогах различают губчатый слой, желатинозное вещество, собственное ядро заднего рога и грудное ядро.
Губчатый слой содержится большое количество мелких вставочных нейронов.
Грудное ядро (ядро Кларка) состоит из крупных вставочных нейронов с сильно разветвленными дендритами. Нейроны ядра Кларка получают информацию от рецепторов мышц, сухожилий и суставов.
В передних рогах расположены самые крупные нейроны корешковые клетки.
В сером веществе спинного мозга много рассеянных пучковых нейронов.
Спинномозговой канал выстлан эпендимоцитами, участвующими в выработке спинномозговой жидкости.
Основную часть остова серого вещества составляют протоплазматические и волокнистые астроциты.Олигодендроглиоциты входят в состав оболочек нервных волокон. Микроглияпоступает в спинной мозг распределяется в сером и белом веществе.
Дата публикования: 2015-10-09 ; Прочитано: 2744 | Нарушение авторского права страницы
Структурно-функциональная единица нервной системы – нервная клетка с ее отростками. Трофическим центром клетки является тело (перикарион); воспринимающие (центрипетальные) отростки носят название дендритов. Отросток, по которому нервный импульс идет центрифугально, от тела клетки к рабочему органу, обозначается как аксон (нейрит). Нервное волокно состоит из аксона (нейрита, осевого цилиндра) и окружающих его шванновских клеток (леммоцитов), образующих неврилемму. В мякотных (миелинизированных) нервных волокнах кнаружи от миелинового слоя располагается неврилемма или шванновская оболочка. На относительно правильных промежутках миелиновая обкладка прерывается и нервное волокно разделяется на сегменты. Каждый сегмент образован одним леммоцитом. Между сегментами имеются промежутки, в которых отсутствует миелиновая оболочка (перехваты Ранвье); именно в этих местах активно происходят обменные процессы, способствующие проведению нервного импульса по аксону.
Источником симпатической иннервации на шейно-грудном уровне являются тела нейронов в боковых рогах серого вещества спинного мозга, от которых идут прегангглионарные миелинизированные волокна, покидающие передние корешки и контактирующие затем с паравертебральными симпатическими ганглиями (симпатический ствол) или входящие в состав черепных нервов. Подобно этому, преганглионарные парасимпатические волокна идут из передних спинальных корешков в область таза, а на краниальном уровне входят в состав III, IX и X пар черепных нервов. Парасимпатические ганглии расположены в связанных с ними эффекторных органах или вблизи от них.
При осмотре невооруженным глазом нерв выглядит как белая шнуроподобная структура с довольно гладкой поверхностью, покрытой плотно прилегающей, но не спаянной с нервом, жировой тканью. В наиболее мощных нервах, таких как седалищный, через нее просвечивают крупные нервные пучки – фасцикулы. На поперечном гистологическом срезе наружная поверхность нерва окружена соединительнотканным футляром – периневрием, состоящим из концентрических слоев жировых клеток, разделенных слоями коллагена. Наконец, эндоневрий также представляет собой футляр, содержащий нервные волокна, шванновские клетки (леммоциты), кровеносные сосуды вместе с пучками тонких эндоневральных коллагеновых волокон, ориентированных вдоль нервных пучков. В эндоневрии содержится также небольшое количеств офибробластов.. Эндоневральный коллаген плотно прилегает к поверхности каждого нервного пучка.
Структура нервных волокон неоднородна. Большинство нервов содержит миелинизированные и немиелинизированные или слабо миелинизированные волокна с неодинаковым соотношением их между собой. Клеточный состав эндоневральных пространств отражает уровень миелинизации. В норме 90% обнаруживаемых в этом пространстве клеточных ядер относится к клеткам Шванна (леммоцитам), а остальные принадлежат фибробластам и капилярному эндотелию. При 80% шванновских клеток окружают немиелинизированных аксоны; рядом с миелинизированными волокнами их количество уменьшено в 4 раза. Тотальный диаметр нервного волокна, т. е. аксон-цилиндра (нейрита) и миелинового футляра, вместе взятых, имеет не только морфологический интерес. Миелинизированные волокна большого диаметра проводят импульсы в значительно более быстром темпе, чем слабо миелинизированные или немиелинизированные. Наличие такой корреляции послужило основой для создания ряда морфолого-физиологических классификаций. Так, Warwick R,. Williams P. (1973) выделяют три класса волокон: А, В и С. А-волокна – соматические афферентные и афферентные миелинизированные нервные волокна, В-волокна – миелинизированные преганглионарные вегетативные волокна, С-волокна – немиелинизированные вегетативные и сенсорные волокна. А. Paintal (1973) модифицировал эту кассификацию с учетом функциональных особенностей волокон, их размеров и скорости проведения импульсов.
Класс А (миелинизированные волокна), афферентные, сенсорные.
Группа I. Волокна размером более 20 мкм в диаметре, со скоростью проведения импульса до 100 м/сек. Волокна этой группы несут импульсы от рецепторов мышц (мышечных веретен, интрафузальных мышечных волокон) и рецепторов сухожилий.
Группа II. Волокна размером от 5 до 15 мкм в диаметре, со скоростью проведения импульсов от 20 до 90 м/сек. Эти волокна несут импульсы от механорецепторов и вторичных окончаний на мышечных веретенах интрафузальных мышечных волокон.
Группа III. Волокна размером от 1 до 7 мкм в диаметре, со скоростью проведения импульса от 12 до 30 м/сек. Функцией этих волокон является болевая рецепция, а также иннервация волосяных рецепторов и сосудов.
Класс А (миелинизированные волокна), эфферентные, двигательные.
Альфа-волокна. Более 17 мкм в диаметре, скорость проведения импульса от 50 до 100 м/сек. Они иннервируют экстрафузальные поперечнополосатые мышечные волокна, преимущественно стимулируя быстрые сокращения мышц (мышечные волокна 2-го типа) и крайне незначительно – медленные сокращения (мышц 1-го типа).
Бета-волокна. В отличие от альфа-волокон иннервируют мышечные волокна 1-го типа (медленные и тонические сокращения мышц) и частично интрафузальные волокна мышечного веретена.
Гамма-волокна. Размером 2-10 мкм в диаметре, скорость проведения импульса 10-45 см/сек, иннервирует только интрафузальные волокна, т. е. мышечное веретено, тем самым участвуя в спинальной саморегуляции мышечного тонуса и движений (кольцевая связь гамма-петли).
Класс В – миелинизированные преганглионарные вегетативные.
Это небольшие нервные волокна, около 3 мкм в диаметре, со скоростью проведения импульса от 3 до 15 м/сек.
Класс С – немиелинизированные волокна, размерами от 0,2 до 1,5 мкм в диаметре, со скоростью проведения импульса от 0,3 до 1,6 м/сек. Этот класс волокон состоит из постганглионарных вегетативных и эфферентных волокон, преимущественно воспринимающих (проводящих) болевые импульсы
Очевидно, что эта классификация интересует и клиницистов, помогая понять некоторые особенности эфферентной и сенсорной функций нервного волокна, в том числе закономерности проведения нервных импульсов, как в норме, так и при различных патологических процессах.
Электрофизиологические исследования показывают, что в состоянии покоя существует разница в электрическом потенциале на внутренней и внешней сторонах неврональной и аксональной клеточной мембраны. Внутренняя часть клетки имеет отрицательный разряд 70-100 мВ по отношению к интерстициальной жидкости снаружи клетки. Этот потенциал поддерживается различием в концентрации ионов. Калий (и белки) преобладают внутри клетки, в то время как ионы натрия и хлориды имеют более высокую концентрацию вне клетки. Натрий постоянно диффундирует в клетку, а калий имеет тенденцию выходить из нее. Дифференциал концентрации натрий-калий поддерживается путем энергозависимого насосного механизма в покоящейся клетке, причем это равновесие существует при слегка сниженной концентрации положительно заряженных ионов внутри клетки, чем снаружи от нее. Это приводит к отрицательному внутриклеточному заряду. Ионы кальция также вносят вклад в поддержание равновесия в клеточной мембране, и когда их концентрация снижается, возбудимость нерва нарастает.
Под влиянием естественной или вызванной внешними факторами стимуляции аксона происходит нарушение селективной проницаемости клеточной мембраны, что способствует проникновению ионов натрия в клетку и редукции потенциала покоя. Если мембранный потенциал снижается (деполяризуется) до критического уровня (30-50 мВ), то возникает потенциал действия и импульс начинает распространяться вдоль клеточной мембраны как волна деполяризации. Важно отметить, то в немиелинизированных волокнах скорость распространения импульса прямо пропорциональна диаметру аксона, и возбуждение длительно прямолинейно захватывает соседствующие мембраны.
Большинство нервов имеет смешанный состав нервных волокон по их диаметру, степени миелинизации (миелинизированные и немиелинизированные волокна), включение вегетативных волокон, дистанциям между перехватами Ранвье, и поэтому каждый нерв имеет свой, смешанный (сложный) потенциал действия и суммированную скорость проведения импульса. Например, у здоровых лиц скорость проведения по нервному стволу, измеренная при накожном наложении электродов, варьирует от 58 до 72 м/сек для лучевого нерва и от 47 до 51 м/сек для малоберцового нерва (M. Smorto, J. Basmajian, 1972).
Информация, передаваемая по нерву, распространяется не только стереотипными электрическими сигналами, но и с помощью химических передатчиков нервного возбуждения – медиаторов или трансмиттеров, освобождаемых в местах соединения клеток – синапсах. Синапсы – специализированные контакты, через которые осуществляется поляризованная, опосредованная химически, передача из нейрона возбуждающих или тормозящих влияний на другой клеточный элемент. В дистальной, терминальной части нервное волокно лишено миелина, образуя терминальную арборизацию (телодендрон) и пресинаптический терминальный элемент. Этот элемент морфологически характеризуется расширением окончания аксона, что напоминает булаву и нередко именуется как пресинаптический мешок, терминальная бляшка, бутон, синаптический узелок. Под микроскопом в этой булаве можно увидеть различных размеров (около 500 А) гранулярные пузырьки или синаптические везикулы, содержащие медиаторы (например, ацетилхолин, катехоламины, пептидные гормоны и др.).
Подмечено, что присутствие круглых пузырьков отвечает возбуждению, а плоских – торможению синапса. Под терминальной бляшкой лежит синаптическая щель размерами 0,2-0,5 мкм в поперечнике, в которую из везикул поступают кванты медиатора. Затем следует субсинаптическая (постсинаптическая) мембрана, воздействуя на которую химический передатчик вызывает изменения электрического потенциала в подлежащих клеточных элементах.
Кровеносные сосуды нервов являются ветвями близрасположенных сосудов. Подходящие к нерву артерии разделяются на восходящую и нисходящую ветви, которые распространяются по нерву. Артерии нервов анастомозируют между собой, образуя непрерывную сеть по ходу всего нерва. Наиболее крупные сосуды расположены в наружном эпиневрии. От них отходят ветви в глубину нерва и проходят в нем между пучками в рыхлых прослойках внутреннего эпиневрия. От этих сосудов ветви проходят к отдельным пучкам нерва, располагаясь в толще периневральных влагалищ. Тонкие ветви этих периневральных сосудов расположены внутри пучков нервных волокон в прослойках эндоневрия (эндоневральные сосуды). Артериолы и прекапилляры вытянуты по ходу нервных волокон, располагаясь между ними.
По ходу седалищного и срединного нерва обычно расположены заметные и достаточно длинные артерии (артерия седалищного нерва, артерия срединного нерва). Эти собственные артерии нервов анастомозируют с ветвями близрасположенных сосудов.
Количество источников кровоснабжения каждого нерва индивидуально различно. Большей или меньшей величины артериальные веточки подходят к крупным нервам через каждые 2-10 см. В связи с этим выделение нерва из окружающей его околонервной клетчатки в какой-то мере сопряжено с повреждением подходящих к нерву сосудов.
Микроваскулярное кровоснабжение нерва, исследованное прижизненным микроскопическим методом показало, что обнаруживаются эндоневральные анастомозы между сосудами в различных слоях нерва. При этом преобладает наиболее развитая сеть внутри нерва. Изучение эндоневрального кровотока имеет большое значение как показатель степени повреждения нерва, при этом кровоток претерпевает немедленные изменения даже при слабой компрессии в эксперименте на животных и на людях, производимой на поверхности нерва или же если компремируются экстраневральные сосуды. При такой экспериментальной компрессии только часть глубокорасположенных в нерве сосудов сохраняют нормальный кровоток (Lundborg G,. 1988).
Вены нервов формируются в эндоневрии, периневрии и эпиневрии. Наиболее крупными венами являются эпиневральные. Вены нервов впадают в близрасположенные вены. Следует отметить, что при затруднениях венозного оттока вены нервов могут расширяться, образуя варикозные узлы.
Лимфатические сосуды нерва. В эндоневрии и в периневральных футлярах имеются лимфатические щели. Они находятся в связи с лимфатическими сосудами в эпиневрии. Отток лимфы от нерва происходит по лимфатическим сосудам, тянущимся в эпиневрии вдоль нервного ствола. Лимфатические сосуды нерва впадают в близрасположеные крупные лимфатические протоки, которые идут к регионарным лимфатическим узлам. Межтканевые эндоневральные щели, пространства периневральных влагалищ являются путями перемещения внутритканевой жидкости.
Нервы (нервные стволы) – это структуры, которые связывают центры головного и спинного мозга с рецепторами и рабочими органами. Они состоят из миелиновых и безмиелиновых афферентных и эфферентных волокон и прослоек соединительной ткани. Могут быть отдельные нейроны и отдельные нервные ганглии. Прослойка рыхлой соединительной ткани, окружающая каждое нервное волокно, называется эндоневрий; окружающая пучок нервных волокон — периневрий, который состоит из 5-6 слоев коллагеновых волокон; между этими слоями имеются щелевидные полости, выстланные нейроэпителием, в которых циркулирует жидкость. Весь нерв окружен прослойкой соединительной ткани, которая называется эпиневрий. В периневрии и эпиневрии имеются кровеносные сосуды и нервы нервов.
Регенерация. При повреждении нерва разрываются проходящие в нем нервные волокна. После разрыва волокна в нем образуются 2 конца — конец, который связан с телом нейрона, называется центральным; конец, не связанный с нервной клеткой, называется периферическим.
В периферическом конце происходят 2 процесса: 1) дегенерация и 2) регенерация. Вначале идет процесс дегенерации, заключающийся в том, что начинается набухание нейролеммоцитов, растворяется миелиНовый слой, осевой цилиндр фрагментируется, образуются капли (овоиды), состоящие из миелина и фрагмента осевого цилиндра. К концу 2-й недели происходит рассасывание овоидов, остается только неврилемма оболочки волокна. Нейролеммоциты продолжают размножаться, из них образуются ленты (тяжи).
После рассасывания овоидов осевой цилиндр центрального конца утолщается и образуется колба роста, которая начинает расти, скользя по лентам нейролеммоцитов. К этому времени между разорванными концами нервных волокон образуется нейроглиально-соединительнотканный рубец, являющийся препятствием для продвижения колбы роста. Поэтому не все осевые цилиндры могут пройти на противоположную сторону образовавшегося рубца. Следовательно, после повреждения нервов иннервация органов или тканей полностью не восстанавливается. Между тем часть осевых цилиндров, оснащенных колбами роста, пробивается на противоположную сторону нейроглиального рубца, погружается в тяжи нейролеммоцитов. Затем мезаксон навертывается на эти осевые цилиндры, образуется миелиновый слой оболочки нервного волокна. В том месте, где находится нервное окончание, рост осевого цилиндра приостанавливается, формируются терминали окончания и все его компоненты.
95. Рефлекторная дуга вегетативного симпатического рефлекса
Преганглионарные нейроны симпатической нервной системы расположены в боковых ядрах спинного мозга, начиная с 8-го шейного сегмента и заканчиваясь 2-м поясничным сегментом включительно. В сегментах 8-го шейного, 1 и 2 грудного сегмента находятся нейроны, возбуждение которых вызывает расширение зрачка (сокращение дилататора зрачка), сокращение глазничной части круговой мышцы глаза, а также сокращение одной из мышц верхнего века.
От 1,2,3,4 и 5 грудных сегментов начинаются преганглионарные симпатические волокна, которые направляются к сердцу и бронхам.
I – Преганглионарные волокна,
II – вегетативные ганглии,
III – постганглионарные волокна и клетки-мишени,
IV– иннервируемые органы, в которых заложены клетки-мишени;
В отличие от парасимпатической нервной системы симпатическая иннервирует почти все органы: сердце, сосуды, бронхи, ГМК желудочно-кишечного тракта, ГМК мочеполовой системы, потовые железы, печень, мышцы зрачка, матку, ткани, в которых совершается липолиз, гликогенолиз, надпочечники, ряд других желез внутренней секреции.
симпатическая нервная система способствует значительному повышению работоспособности организма – под ее влиянием возрастает гликогенолиз, глюконеогенез, липолиз, усиливается деятельность сердечно-сосудистой системы, происходит перераспределение массы крови из областей, способных переносить гипоксию, в области, где наличие кислорода и энергетических источников является основой существования. Происходит улучшение вентиляции легких. Одновременно, при активации симпатической нервной системы имеет место торможение деятельности желудочно-кишечного тракта, мочевого пузыря.
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА. СПИННОМОЗГОВЫЕ НЕРВЫ
Нервная система человека подразделяется на центральную, периферическую и автономную части. Периферическая часть нервной системы представляет собой совокупность спинномозговых и черепных нервов. К ней относятся образуемые нервами ганглии и сплетения, а также чувствительные и двигательные окончания нервов. Таким образом, периферическая часть нервной системы объединяет все нервные образования, лежащие вне спинного и головного мозга. Такое объединение в известной мере условно, так как эфферентные волокна, входящие в состав периферических нервов, являются отростками нейронов, тела которых находятся в ядрах спинного и головного мозга. С функциональной точки зрения периферическая часть нервной системы состоит из проводников, соединяющих нервные центры с рецепторами и рабочими органами. Анатомия периферических нервов имеет большое значение для клиники, как основа для диагностики и лечения заболеваний и повреждений этого отдела нервной системы.
Строение нервов
Периферические нервы состоят из волокон, имеющих различное строение и неодинаковых в функциональном отношении. В зависимости от наличия или отсутствия миелиновой оболочки волокна бывают миелиновые (мякотные) или безмиелиновые (безмякотные). По диаметру миелиновые нервные волокна подразделяются на тонкие (1-4 мкм), средние (4-8 мкм) и толстые (более 8 мкм). Существует прямая зависимость между толщиной волокна и скоростью проведения нервных импульсов. В толстых миелиновых волокнах скорость проведения нервного импульса составляет примерно 80-120 м/с, в средних - 30-80 м/с, в тонких - 10-30 м/с. Толстые миелиновые волокна являются преимущественно двигательными и проводниками проприоцептивной чувствительности, средние по диаметру волокна проводят импульсы тактильной и температурной чувствительности, а тонкие - болевой. Безмиелиновые волокна имеют небольшой диаметр - 1-4 мкм и проводят импульсы со скоростью 1-2 м/с. Они являются эфферентными волокнами вегетативной нервной системы.
Таким образом, по составу волокон можно дать функциональную характеристику нерва. Среди нервов верхней конечности наибольшее содержание мелких и средних миелиновых и безмиелиновых волокон имеет срединный нерв, а наименьшее число их входит в состав лучевого нерва, локтевой нерв занимает в этом отношении среднее положение. Поэтому при повреждении срединного нерва бывают особенно выражены болевые ощущения и вегетативные расстройства (нарушения потоотделения, сосудистые изменения, трофические расстройства). Соотношение в нервах миелиновых и безмиелиновых, тонких и толстых волокон индивидуально изменчиво. Например, количество тонких и средних миелиновых волокон в срединном нерве может у разных людей колебаться от 11 до 45%.
Нервные волокна в стволе нерва имеют зигзагообразный (синусоидальный) ход, что предохраняет их от перерастяжения и создает резерв удлинения в 12-15% от их первоначальной длины в молодом возрасте и 7-8% в пожилом возрасте.
Нервы обладают системой собственных оболочек. Наружная оболочка, эпиневрий, покрывает нервный ствол снаружи, отграничивая его от окружающих тканей, и состоит из рыхлой неоформленной соединительной ткани. Рыхлая соединительная ткань эпиневрия выполняет все промежутки между отдельными пучками нервных волокон. Некоторые авторы называют эту соединительную ткань внутренним эпиневрием, в отличие от наружного эпиневрия, окружающего нервный ствол снаружи.
В эпиневрии в большом количестве находятся толстые пучки коллагеновых волокон, идущих преимущественно продольно, клетки фибробластического ряда, гистиоциты и жировые клетки. При изучении седалищного нерва человека и некоторых животных установлено, что эпиневрия состоит из продольных, косых и циркулярных коллагеновых волокон, имеющих зигзагообразный извилистый ход с периодом 37-41 мкм и амплитудой около 4 мкм. Следовательно, эпиневрия - очень динамичная структура, которая защищает нервные волокна при растяжении и изгибе.
Из эпиневрия выделен коллаген I типа, фибриллы которого имеют диаметр 70-85 нм. Однако некоторые авторы сообщают о выделении из зрительного нерва и других типов коллагена, в частности III, IV, V, VI. Нет единого мнения о природе эластических волокон эпиневрия. Одни авторы считают, что в эпиневрии отсутствуют зрелые эластические волокна, но обнаружены два вида близких к эластину волокон: окситалановые и элауниновые, которые располагаются параллельно оси нервного ствола. Другие исследователи считают их эластическими волокнами. Жировая ткань является составной частью эпиневрия. Седалищный нерв содержит обычно значительное количество жира и этим заметно отличается от нервов верхней конечности.
При исследовании черепных нервов и ветвей крестцового сплетения взрослых людей установлено, что толщина эпиневрия колеблется в пределах от 18-30 до 650 мкм, но чаще составляет 70-430 мкм.
Эпиневрий - в основном питающая оболочка. В эпиневрии проходят кровеносные и лимфатические сосуды, vasa nervorum, которые проникают отсюда в толщу нервного ствола.
Следующая оболочка, периневрий, покрывает пучки волокон, из которых состоит нерв. Она является механически наиболее прочной. При световой и электронной микроскопии установлено, что периневрий состоит из нескольких (7-15) слоев плоских клеток (периневрального эпителия, нейротелия) толщиной от 0.1 до 1.0 мкм, между которыми располагаются отдельные фибробласты и пучки коллагеновых волокон. Из периневрия выделен коллаген III типа, фибриллы которого имеют диаметр 50-60 нм. Тонкие пучки коллагеновых волокон расположены в периневрии без особого порядка. Тонкие коллагеновые волокна образуют в периневрии двойную спиральную систему. Причем волокна образуют в периневрии волнистые сети с периодичностью около 6 мкм. Установлено, что пучки коллагеновых волокон имею в периневрии плотное расположение и ориентированы как в продольном, так и концентрическом направлениях. В периневрии найдены элауниновые и окситалановые волокна, ориентированные преимущественно продольно, причем первые в основном локализуются в поверхностном его слое, а вторые - в глубоком слое.
Толщина периневрия в нервах с многопучковой структурой находится в прямой зависимости от величины покрываемого им пучка: вокруг мелких пучков не превышает 3-5 мкм, крупные пучки нервных волокон покрываются периневральным футляром толщиной от 12-16 до 34-70 мкм. Данные электронной микроскопии свидетельствуют, что периневрий имеет гофрированную, складчатую организацию. Периневрию придается большое значение в барьерной функции и обеспечении прочности нервов.
Периневрий, внедряясь в толщу нервного пучка, образует там соединительнотканные перегородки толщиной 0.5-6.0 мкм, которые делят пучок на части. Подобная сегментация пучков чаще наблюдается в поздних периодах онтогенеза.
Периневральные влагалища одного нерва соединяются с периневральными влагалищами соседних нервов, и через эти соединения происходит переход волокон из одного нерва в другой. Если учесть все эти связи, то периферическую нервную систему верхней или нижней конечности можно рассматривать как сложную систему связанных между собой периневральных трубок, по которым осуществляется переход и обмен нервных волокон как между пучками в пределах одного нерва, так и между соседними нервами.
Самая внутренняя оболочка, эндоневрий, покрывает тонким соединительнотканным футляром отдельные нервные волокна. Клетки и внеклеточные структуры эндоневрия вытянуты и ориентированы преимущественно по ходу нервных волокон. Количество эндоневрия внутри периневральных футляров по сравнению с массой нервных волокон невелико. Эндоневрий содержит коллаген III типа с фибриллами диаметром 30-65 нм. Мнения о наличии в эндоневрии эластических волокон весьма противоречивы. Одни авторы считают, что эндоневрий не содержит эластических волокон. Другие обнаружили в эндоневрии близкие по свойствам к эластическим окситалановые волокна с фибриллами диаметром 10-12.5 нм, ориентированные, главным образом, параллельно аксонам.
Известно, что нервные волокна сгруппированы в отдельные пучки различного калибра. У разных авторов существуют различные определения пучка нервных волокон в зависимости от позиции, с которой эти пучки рассматриваются: с точки зрения нейрохирургии и микрохирургии или с точки зрения морфологии. Классическим определением нервного пучка является группа нервных волокон, ограниченная от других образований нервного ствола периневральной оболочкой. И этим определением руководствуются при исследовании морфологи. Однако при микроскопическом исследовании нервов часто наблюдаются такие состояния, когда несколько групп нервных волокон, прилежащих друг к другу, имеют не только собственные периневральные оболочки, но и окружены общим периневрием. Эти группы нервных пучков часто бывают видны при макроскопическом исследовании поперечного среза нерва во время нейрохирургического вмешательства. И эти пучки чаще всего описываются при клинических исследованиях. Из-за различного понимания строения пучка происходят в литературе противоречия при описании внутриствольного строения одних и тех же нервов. В связи с этим ассоциации нервных пучков, окруженные общим периневрием, получили название первичных пучков, а более мелкие, их составляющие, - вторичных пучков.
На поперечном срезе нервов человека соединительнотканные оболочки (эпиневрий, периневрий) занимают значительно больше места (67.03-83.76%), чем пучки нервных волокон. Показано, что количество соединительной ткани зависит от числа пучков в нерве. Ее значительно больше в нервах с большим количеством мелких пучков, чем в нервах с немногими крупными пучками.
Показано, что пучки в нервных стволах могут располагаться относительно редко с промежутками в 170-250 мкм, и более часто - расстояние между пучками менее 85-170 мкм.
В зависимости от строения пучков выделяют две крайние формы нервов: малопучковую и многопучковую. Первая характеризуется небольшим количеством толстых пучков и слабым развитием связей между ними. Вторая состоит их множества тонких пучков с хорошо развитыми межпучковыми соединениями.
Когда количество пучков небольшое, пучки имеют значительные размеры, и наоборот. Малопучковые нервы отличаются сравнительно небольшой толщиной, наличием небольшого количества крупных пучков, слабым развитием межпучковых связей, частым расположением аксонов внутри пучков. Многопучковые нервы отличаются большей толщиной и состоят из большого количества мелких пучков, в них сильно развиты межпучковые связи, аксоны располагаются в эндоневрии рыхло.
Толщина нерва не отражает количества содержащихся в нем волокон, и не существует закономерностей расположения волокон на поперечном срезе нерва. Однако установлено, что в центре нерва пучки всегда тоньше, на периферии - наоборот. Толщина пучка не характеризует количества заключенных в нем волокон.
В строении нервов установлена четко выраженная асимметрия, то есть неодинаковое строение нервных стволов на правой и левой сторонах тела. Например, диафрагмальный нерв имеет слева большее количество пучков, чем справа, а блуждающий нерв - наоборот. У одного человека разница в количестве пучков между правым и левым срединными нервами может варьировать от 0 до 13, но чаще составляет 1-5 пучков. Разница в количестве пучков между срединными нервами разных людей равняется 14-29 и с возрастом увеличивается. В локтевом нерве у одного и того же человека разница между правой и левой сторонами в количестве пучков может колебаться от 0 до 12, но чаще составляет также 1-5 пучков. Различие в количестве пучков между нервами разных людей достигает 13-22.
Разница между отдельными субъектами в количестве нервных волокон колеблется в срединном нерве от 9442 до 21371, в локтевом нерве - от 9542 до 12228. У одного и того же человека разница между правой и левой стороной варьирует в срединном нерве от 99 до 5139, в локтевом нерве - от 90 до 4346 волокон.
Источниками кровоснабжения нервов являются соседние близлежащие артерии и их ветви. К нерву обычно подходят несколько артериальных ветвей, причем интервалы между входящими сосудами варьируют в крупных нервах от 2-3 до 6-7 см, а в седалищном нерве - до 7-9 см. Кроме того, такие крупные нервы, как срединный и седалищный, имеют собственные сопровождающие артерии. В нервах, имеющих большое количество пучков, в эпиневрии содержится много кровеносных сосудов, причем они имеют сравнительно малый калибр. Наоборот, в нервах с небольшим количеством пучков сосуды одиночные, но значительно более крупные. Артерии, питающие нерв, в эпиневрии Т-образно делятся на восходящую и нисходящую ветви. Внутри нервов артерии делятся до ветвей 6-го порядка. Сосуды всех порядков анастомозируют между собой, образуя внутриствольные сети. Эти сосуды играют значительную роль в развитии коллатерального кровообращения при выключении крупных артерий. Каждая артерия нерва сопровождается двумя венами.
Лимфатические сосуды нервов находятся в эпиневрии. В периневрии между его слоями образуются лимфатические щели, сообщающиеся с лимфатическими сосудами эпиневрия и эпиневральными лимфатическими щелями. Таким образом, по ходу нервов может распространяться инфекция. Из больших нервных стволов обычно выходят несколько лимфатических сосудов.
Оболочки нервов иннервируются ветвями, отходящими от данного нерва. Нервы нервов имеют в основном симпатическое происхождение и по функции являются сосудодвигательными.
Читайте также:
