
Нервная регуляция сокращения скелетных мышц
В большинстве движений участвует множество мышц, причем сокращение и расслабление различных групп мышц происходит в определенном порядке и с определенной силой. Такая согласованность движений называется координацией движений. Она осуществляется нервной системой. Скелетные мышцы иннервируются соматическим отделом нервной системы. К каждой мышце подходит один или несколько нервов, проникающих в ее толщу и разветвляющихся на множество мелких отростков, которые достигают мышечных волокон. Посредством нервов осуществляется связь мышц с ЦНС, которая регулирует любые двигательные акты (ходьба, бег, пищевые движения и т. д.) и длительное напряжение мышц — тонус, поддерживающий определенное положение тела в пространстве. Деятельность мышц носит рефлекторный характер. Мышечный рефлекс может запускаться с раздражения рецепторов, находящихся в самой мышце или в сухожилиях, либо с раздражения зрительных, слуховых, обонятельных, осязательных рецепторов.
В регуляции безусловно-рефлекторных движений принимает участие мозжечок. Он осуществляет координацию движения, регуляцию мышечного тонуса, способствует поддержанию равновесия и позы тела. При поражении мозжечка его регуляторные двигательные функции нарушаются.
Сокращаясь, мышца действует на кость как на рычаг и производит механическую работу. На осуществление работы мышцы затрачивается энергия, которая образуется в результате распада и окисления органических веществ, поступивших в мышечную клетку. Основным источником энергии является АТФ. Кровь доставляет мышцам питательные вещества и кислород и уносит образующиеся продукты диссимиляции (углекислый газ и др.). При длительной работе наступает утомление и снижение работоспособности мышцы, возникающее из-за несоответствия между ее кровоснабжением и возросшими потребностями в питательных веществах и кислороде. Систематическая мышечная работа усиливает кровоснабжение мыши и костей, к которым они прикрепляются. Это приводит к увеличению мышечной массы и усиленному росту костей. Сильные мышцы легко справляются с поддержанием туловища в нужном положении, противостоят развитию сутулости, искривлению позвоночника.
ЗАКЛЮЧЕНИЕ
Значение скелета очень велико.Костная система выполняет ряд функций, имеющих или преимущественно механическое, или преимущественно биологическое значение. Рассмотрим функции, имеющие преимущественно механическое значение. Для всех позвоночных характерен внутренний скелет, хотя среди них встречаются виды, которые, наряду с внутренним скелетом, имеют еще и более или менее развитый наружный скелет, возникающий в коже (костная чешуя в коже рыб). В начале своего появления твердый скелет служил для защиты организма от вредных внешних влияний (наружный скелет беспозвоночных). С развитием внутреннего скелета у позвоночных он сначала стал опорой и каркасом для мягких тканей. Отдельные части скелета превратились в рычаги, приводимые в движение мышцами, вследствие чего скелет приобрел локомоторную функцию. В итоге механические функции скелета проявляются в его способности осуществлять защиту, опору и движение.
Опора достигается прикреплением мягких тканей и органов к различным частям скелета. Движение возможно благодаря тому, что кости являются длинными и короткими рычагами, соединенными подвижными сочленениями и приводимыми в движение мышцами, управляемыми нервной системой.
Наконец, защита осуществляется путем образования из отдельных
костей костного канала — позвоночного, защищающего спинной мозг, костной коробки — черепа, защищающего головной мозг; костной клетки — грудной, защищающей жизненно важные органы грудной полости (сердце, легкие, печень, желудок, селезенку, частично почки и др., то есть важнейшие органы разных систем); костного вместилища — таза, защищающего важные для продолжения вида органы размножения, выделения.
Биологическая функция костной системы связана с участием скелета в обмене веществ, особенно в минеральном обмене (скелет является депо минеральных солей — фосфора, кальция, железа и др.). Это важно учитывать для понимания болезней обмена (рахит и др.) и для диагностики с помощью лучи-стой энергии (рентгеновские лучи, радионуклиды). Кроме того, скелет выполняет еще кроветворную функцию. При этом кость является не просто защитным футляром для костного мозга, а последний составляет органическую часть ее. Определенное развитие и деятельность костного мозга отражаются на строении костного вещества, и, наоборот, механические факторы сказываются на функции кроветворения: усиленное движение способствует кроветворению, поэтому при разработке физических упражнений необходимо учитывать единство всех функций скелета.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
В большинстве движений участвует множество мышц, причем сокращение и расслабление различных групп мышц происходит в определенном порядке и с определенной силой. Такая согласованность движений называется координацией движений. Она осуществляется нервной системой. Скелетные мышцы иннервируются соматическим отделом нервной системы. К каждой мышце подходит один или несколько нервов, проникающих в ее толщу и разветвляющихся на множество мелких отростков, которые достигают мышечных волокон. Посредством нервов осуществляется связь мышц с ЦНС, которая регулирует любые двигательные акты (ходьба, бег, пищевые движения и т. д.) и длительное напряжение мышц — тонус, поддерживающий определенное положение тела в пространстве. Деятельность мышц носит рефлекторный характер. Мышечный рефлекс может запускаться с раздражения рецепторов, находящихся в самой мышце или в сухожилиях, либо с раздражения зрительных, слуховых, обонятельных, осязательных рецепторов.
В регуляции безусловно-рефлекторных движений принимает участие мозжечок. Он осуществляет координацию движения, регуляцию мышечного тонуса, способствует поддержанию равновесия и позы тела. При поражении мозжечка его регуляторные двигательные функции нарушаются.
Сокращаясь, мышца действует на кость как на рычаг и производит механическую работу. На осуществление работы мышцы затрачивается энергия, которая образуется в результате распада и окисления органических веществ, поступивших в мышечную клетку. Основным источником энергии является АТФ. Кровь доставляет мышцам питательные вещества и кислород и уносит образующиеся продукты диссимиляции (углекислый газ и др.). При длительной работе наступает утомление и снижение работоспособности мышцы, возникающее из-за несоответствия между ее кровоснабжением и возросшими потребностями в питательных веществах и кислороде. Систематическая мышечная работа усиливает кровоснабжение мыши и костей, к которым они прикрепляются. Это приводит к увеличению мышечной массы и усиленному росту костей. Сильные мышцы легко справляются с поддержанием туловища в нужном положении, противостоят развитию сутулости, искривлению позвоночника.
16. Движения - это элементарные двигательные акты, характерные для того или иного сустава в зависимости от его физиологической подвижности. Фактически данный прием - это ряд физических упражнений, используемых с целью воздействия на отдельные мышечные группы и суставы. Выполняется с медленной скоростью и максимальной амплитудой. Объем движения в суставе зависит от его строения и разности угловых размеров суставных поверхностей: вокруг фронтальной оси - сгибание и разгибание; сагиттальной - приведение и отведение; продольной - вращение, а также пронация и супинация; при комбинированном движении вокруг всех осей - круговое движение. Движения широко применяются в гигиеническом, спортивном и лечебном видах массажа; в ряде случаев им отводится 20-30% времени проводимого сеанса. Они используются при восстановлении работоспособности после физических нагрузок, после травм и заболеваний суставов. В спорте, балете, а также танцевальных коллективах движения в комплексе с растираниями используются при подготовке суставно-связочного аппарата к нагрузке, часто - к нестандартным движениям в суставах.
Физиологическое воздействие движений на организм:
- благотворно влияют на опорно-двигательный аппарат, воздействуя не только на мышцы, связочный аппарат и суставы, но и на реактивность организма в целом;
- увеличивают подвижность в суставах (при ограниченной подвижности);
- улучшают крово- и лимфообращение в области суставов;
- улучшают секрецию синовиальной жидкости;
- улучшают эластичность связок и мышц;
- увеличивают обменные процессы и ускоряют процессы регенерации в тканях;
- под влиянием движений задерживается развитие атрофии, вызванной гиподинамией;
- движения дают хороший эффект при сморщиваниях и укорочениях связочного аппарата, мышечных контрактурах, сращениях и отложениях солей;
- под влиянием движений улучшается трофика внутренних органов, интенсивность пищеварения, увеличиваются защитные свойства и сопротивляемость организма в целом.
Виды движений: 1. Активные. 2. Движения с сопротивлением. 3. Пассивные.
Активные движениявыполняются без участия внешней силы (самим пациентом по команде массажиста) после соответствующего массажа мышц и суставов. Они применяются в том случае, когда необходимо возбудить, усилить работу центральной или периферийной нервной системы, а также для укрепления ослабленного мышечного аппарата (после травм, заболеваний и хирургических вмешательств).

Сокращения мышц контролируются двумя видами проприоцепторов: интрафузальным рецепторами (рецепторы растяжения) и сухожильными рецепторами Гольджи (рецепторы напряжения). Мышечное веретено представляет собой соединительнотканную капсулу, которая окружает группу особых, интрафузальных мышечных волокон. Они тоньше и короче обычных экстрафузальных волокон: всё веретено имеет длину от 4 до 10 мм. В средней части интрафузального волокна сократительных белков нет, там находятся окончания чувствительного нейрона, которые возбуждаются при растяжении волокна. Интрафузальные волокна иннервируются гамма-мотонейронами в отличие от aльфа-мотонейронов, предназначенных лишь для экстрафузальных волокон.
Когда внешняя сила растягивает мышцу, возбуждение от рецепторов интрафузальных волокон увеличивается (рис. 7.3). Чувствительный нейрон двумя коллатералями своего аксона одновременно передаёт возбуждение a- и g-мотонейронам и вслед за этим происходит одновременное возбуждение – это называется альфа-гамма-коактивацией, а затем и сокращение интра- и экстрафузальных волокон: это рефлекс растяжения, который обеспечивает регуляцию длины мышцы.
ЦНС с помощью g-мотонейронов контролирует чувствительность мышечных веретён. Так, например, g-мотонейроны могут возбуждаться двигательными ядрами головного мозга, что приведёт к сокращению концевых участков интрафузальных волокон и растяжению центральной их части, в которой находятся окончания чувствительного нейрона. Это вызовет возбуждение чувствительного нейрона, он подействует на a- и g-мотонейроны, что закончится небольшим сокращением мышцы – повышением её тонуса.

Таким образом, мышечный тонус регулируется с помощью g-мотонейронов. Если двигательные ядра головного мозга повысят их активность, то и мышечный тонус повысится. Но, если влияние головного мозга на g-мотонейроны будет опосредовано тормозными интернейронами спинного мозга, то их активность, а затем и мышечный тонус уменьшатся. В физиологических условиях часто бывает необходимо повышать тонус одних мышц и понижать – тонус других, поэтому происходит и стимуляция, и торможение активности различных g-мотонейронов. Благодаря этому одни мышцы могут находиться в большем тонусе, чем другие – именно это позволяет удерживать определённую позу и переходить от неё к движению. Состояние тревоги, неуверенности, страха обычно сопровождается увеличением тонуса всех мышц, механизм этого явления состоит в усилении активирующих и уменьшении тормозных влияний головного мозга на g-мотонейроны.
Сухожильные рецепторы Гольджи почти не реагируют на изменение длины мышцы, но возбуждаются при изменении мышечного напряжения. Чем оно сильнее, тем больше натягиваются коллагеновые волокна сухожилий, прикрепляющие мышцу к костям. Рецепторы Гольджи, вплетённые в ткань сухожильных волокон, при этом сдавливаются, а распространяющееся от них по чувствительному нейрону возбуждение переключается в спинном мозгу на тормозной интернейрон (рис. 7.5), уменьшающий активность мотонейрона и ограничивающий тем самым дальнейший рост мышечного напряжения.
Дата добавления: 2015-01-21 ; просмотров: 1225 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Ткани и органы. Мышцы
Регуляция сокращения мышечных волокон
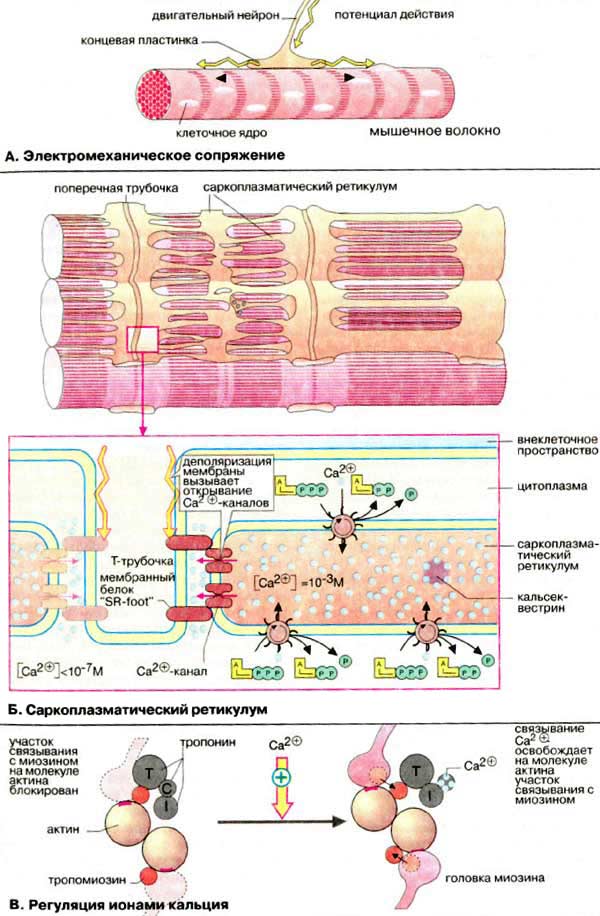
А. Электромеханическое сопряжение
Сокращением мышечного волокна управляют двигательные нейроны , которые выделяют нейромедиатор ацетилхолин в нервно-мышечные соединении (синапсы) (см. рис. 345). Ацетилхолин диффундирует через синаптическую щель и взаимодействуют с ацетилхолиновыми (холинэргическими) рецепторами плазматической мембраны мышечных клеток. Это вызывает открывание трансмембранных ионных каналов и деполяризацию клеточной мембраны (образование потенциала действия). Потенциал действия быстро распространяется по всем направлениям от нервно-мышечного соединения (см. рис. 341, 343), возбуждая все мышечные клетки. В течение нескольких миллисекунд реализуется рассмотренный выше цикл сокращения мышечного волокна.
Б. Саркоплазматический ретикулум
Саркоплазматический ретикулум [СР (SR)] -разветвленная подобная эндоплазматическому ретикулуму органелла, окружающая индивидуальные миофибриллы подобно сетке (в верхней части схемы в качестве примера приведен СР клетки сердечной мышцы). В покоящихся клетках концентрация Са 2+ очень низка (менее 10 -5 М). Однако в саркоплазматическом ретикулуме уровень ионов Са 2+ существенно выше (около 10 -3 М). Высокая концентрация Са 2+ в СР поддерживается Са 2+ -АТФ-азами. Кроме того, в СР имеется специальный белок кальсеквестрин (55 кДа), который благодаря высокому содержанию кислых аминокислот способен прочно связывать ионы Са 2+ .
Переносу потенциала действия на СР индивидуальной миофибриллы способствуют поперечные трубочки Т-системы , представляющие трубчатые впячивания клеточной мембраны и находящиеся в тесном контакте с индивидуальными миофибриллами. Деполяризация плазматической мембраны передается через Т-трубочки на потенци ал-управляемый мембранный белок (так называемый "SR-foot") прилегающей мембраны СР, который открывает Са 2+ -каналы. Результатом является выброс ионов Са 2+ из СР в пространство между филаментами актина и миозина до уровня ≥10 -5 M. В конечном итоге выброс ионов Са 2+ является пусковым механизмом процесса сокращения миофибрилл.
В. Регуляция ионами кальция
В расслабленной скелетной мышце комплекс тропонина (субъединицы = Т, С, I ) с тропомиозином препятствует взаимодействию миозиновых головок с актином .
Быстрое увеличение в цитоплазме концентрации ионов кальция в результате открывания каналов СР приводит к связыванию Са 2+ с С-субъединицей тропонина. Последняя по свойствам близка кальмодулину (см. рис. 375). Связывание ионов Са 2+ вызывает конформационную перестройку в тропонине, тропонинтропомиозиновый комплекс разрушается и освобождает на молекуле актина участок связывания с миозином (на схеме выделен красным цветом). Это инициирует цикл мышечного сокращения (см. с. 324)
В отсутствие последующего стимулирования АТФ-зависимые кальциевые насосы мембраны СР быстро снижают концентрацию ионов Ca 2+ до исходного уровня. Как следствие, комплекс Са 2+ с тропонином С диссоциирует, тропонин восстанавливает исходную конформацию, место связывания миозина на актине блокируется и мышца расслабляется.
Таким образом, при сокращении мышечного волокна скелетных мышц позвоночных происходит следующая последовательность событий. При поступлении сигнала от двигательного нейрона мембрана мышечной клетки деполяризуется, сигнал передается на Сa 2+ -каналы СР. Са 2+ -каналы открываются, внутриклеточный уровень ионов Са 2+ возрастает. Ионы Сa 2+ связывается с тропонином С, вызывая конформационную перестройку в тропонине, что влечет за собой разрушение комплекса тропонин-тропомиозин и дает возможность головкам миозина связываться с актином. Происходит инициация актин-миозинового цикла.
По завершении сокращения уровень ионов Са 2+ снижается за счет активного обратного транспорта Са 2+ в СР, тропонин С отдает Са 2+ , комплекс тропонин-тропомиозин занимает исходное положение на молекуле актина, блокируя актин-миозиновый цикл. Результатом является расслабление мышцы.
Мышцы — главный биохимический преобразователь потенциальной (химической) энергии в кинетическую (механическую). Мышечная ткань занимает первое место по объему среди других тканей человека; на ее долю при рождении приходится чуть меньше 25%, у людей среднего возраста — более 40%, а у пожилых — чуть меньше 30% общей массы тела.
В организме позвоночных существуют три типа мышц: скелетные, сердечные и гладкие. Скелетные и сердечные мышцы при микроскопическом исследовании обнаруживают поперечную исчерченность; в гладких мышцах такая исчерченность отсутствует. В то время как скелетные мышцы находятся под волевым нервным контролем, сердечная и гладкая мышцы функционируют непроизвольно.
Рис. 56.1. Структура поперечнополосатой мышцы. (Drawing by Sylvia Colard Keene. Reproduced, with permission, from Bloom W., Fawcett D. W. A Textbook of Histology, 10th ed. Saunders, 1975.)
Поперечнополосатая мышца состоит из многоядерных клеток (мышечных волокон), окруженных электровозбудимой мембраной — сарколеммой. При микроскопическом исследовании отдельной мышечной клетки, которая может быть вытянута во всю длину мышпы, в ней обнаруживается пучок, состоящий из множества параллельно расположенных миофибрилл; они погружены во внутриклеточную жидкость, называемую саркоплазмой. Эта жидкость содержит гликоген, макроэргические соединения (АТР и фосфокреатин) и ферменты гликолиза.
Саркомер—это функциональная единица мышцы. Саркомеры следуют друг за другом вдоль оси фибриллы, повторяясь через каждые 1500—2300 нм (рис. 56.1). При исследовании миофибриллы в электронном микроскопе выявляется чередование темных и светлых дисков (диски А и I). Центральная область диска А (зона Н) выглядит менее плотной, чем остальная его часть. Диск I делит пополам очень плотная и узкая линия Z. Эти детали мышечной структуры представлены на рис. 56.2.
Исчерченность мышц, видимая под световым микроскопом, - это результат высокой степени их организации, когда большинство мышечных клеток выстраивается таким образом, что их саркомеры располагаются параллельно (рис. 56.1).
Рис. 56.2. Расположение филаментов в поперечнополосатой мышце. А. Расслабленная мышца. Б. Сокращенная мышца.
волокнистым сегментом, который может изгибаться, регулируя пространство между филаментами.
Масса свежих мышечных волокон на 75% состоит из воды и содержит более 20% белка. Два главных мышечных белка — актин и миозин.
В поперечнополосатых мышцах присутствуют еще четыре белка, минорных в плане их вклада в массу мышечной ткани, но выполняющих важную функцию. Тропомиозин представляет собой вытянутую в виде тяжа молекулу, состоящую из двух цепей, а и р, и примыкающую к F-актину в щели между двумя полимерами (рис. 56.3). Этот белок имеется во всех мышцах и подобных им структурах. Характерной особенностью именно поперечнополосатых мышц является наличие в них тропониновой системы, включающей три разных белка. Тропонин Т (ТпТ) так же, как и два других тропониновых компонента, связывается с тропомиозином (рис. 56.3). Тропонин I (Tnl) ингибирует взаимодействие между F-актином и миозином и также связывается с другими компонентами тропонина. Тропонин С (ТпС) -
Рис. 56.3. Схематическое изображение тонкого филамента. Показана пространственная конфигурация трех главных белковых компонентов: актина, тропомиозина и тропонина.
кальций-связывающий белок, первичная и вторичная структура, а также функция которого, совершенно аналогичны соответствующим характеристикам широко распространенного в природе белка — кальмодулина. И тропонин С, и кальмодулин связывают четыре иона кальция на молекулу белка и имеют мол. массу 17000. Тонкий филамент поперечнополосатой мышцы состоит из F-актина, тропомиозина и трех тропониновых компонентов: ТпС, ТпI и ТпТ (рис. 56.3). Тропомиозин и тропониновая система чередуются через каждые 38,5 нм.
Большая часть сведений о миозине получена при изучении продуктов его частичного гидролиза. Обработка миозина трипсином приводит к образованию двух фрагментов — меромиозинов. Легкий меромиозин (ЛММ) состоит из агрегированных нерастворимых а-спиральных фибрилл (рис. 56.5). Он не обладает АТРазной активностью и не связывается с F-актином.
Тяжелый меромиозин (ТММ) представляет собой растворимый белок с мол. массой 340 000, содержащий и фибриллярный, и глобулярный фрагменты (рис. 56.5). Он обладает АТРазной активностью и связывается с F-актином. При гидролизе ТММ
Рис. 56.4. Схема молекулы миозина с двумя переплетенными а-спиралями (фибриллярная часть), глобулярной областью (G) и легкими цепями (L).
Рис. 56.5. Ферментативное расщепление миозина. ТММ — тяжелый меромиозин; ЛММ — легкий меромиозин; S-1-фрагмент 1; S-2-фрагмент 2.
папаином образуются два субфрагмента, S-1 и S-2. S-2 имеет фибриллярную структуру, не проявляет АТРазной активности и не связывает F-актин.
а-Акгинин — это обнаруживаемая в зоне Z-линии белковая молекула, к которой присоединяются концы F-актиновых молекул тонких филаментов (рис. 56.2).
1) Миозиновая головка сама по себе может гидролизовать АТР до ADP и неорганического фосфата, но не обеспечивает освобождение продуктов гидролиза. Следовательно, этот процесс носит скорее стехиометрический, чем каталитический характер.
2) Миозиновая головка, содержащая ADP и неорганический фосфат, может свободно вращаться под большими углами и (при достижении нужного положения) связываться с F-актином, образуя с осью фибриллы угол около 90°.
Рис. 56.7. Гидролиз АТР запускает цикл ассоциации и диссоциации актина и миозина в пяти реакциях, описанных в тексте; Pj-неорганический фосфат. (Modified from Stryer L. Biochemistry, 2nd ed. Freeman, 1981.)
3) Это взаимодействие обеспечивает высвобождение ADP и неорганического фосфата из актин— миозинового комплекса. Поскольку наименьшую энергию актомиозиновая связь имеет при величине угла 45°, миозин изменяет свой угол с осью фибриллы с 90° на примерно 45°, продвигая актин (на 10—15 нм) в направлении центра саркомера.
4) Новая молекула АТР связывается с комплексом миозин-F-актин.
5) Комплекс миозин—АТР обладает низким сродством к актину и поэтому происходит отделение миозиновой (АТР) головки от F-актина. Последняя стадия и есть собственно расслабление, которое таким образом отчетливо зависит от связывания АТР с актин—миозиновым комплексом. АТР вновь гидролизуется миозиновой головкой без высвобождения ADP и неорганического фосфата, и цикл возобновляется.
Таким образом, АТР отсоединяет миозиповую головку от тонкой нити и является движущей силой сокращения. Эффективность такого сокращения — около 50%; эффективность двигателя внутреннего сгорания — менее 20%.
Актиновая регуляция характерна для поперечнополосатых мышц позвоночных—скелетных и сердечной. Согласно общему механизму, рассмотренному выше, единственным потенциально лимитирующим фактором в цикле мышечного сокращения может быть АТР. Скелетные мышцы ингибируются в покое и деингибируются с активацией сокращения. Роль ингибитора в поперечнополосатых мышцах выполняет тропониновая система, связанная в тонких филаментах с тропомиозином и F-актином (рис. 56.3). При отсутствии тропомиозин—тропониновой системы регуляция сокращения поперечнополосатых мышц (или АТРазы как биохимического индикатора сокращения) не осуществляется. Как отмечалось выше, тропомиозин локализуется в щели F-актина, а три компонента тропонина-TnT, ТпI и ТпС-
5 моль/л. Са2+-связывающие сайты на ТпС в тонком филаменте быстро насыщаются Са2+. Комплекс ТпС-4 Са2+ реагирует с ТпI и ТпТ, влияя на их взаимодействие с тропомиозином. Последний в соответствии с этим либо просто отсоединяется, либо изменяет конформацию F-актина таким образом, что появляется возможность взаимодействия ADP Ргмиозиновой головки с F-актином и начинается сократительный цикл.
Как отмечалось выше, во всех мышцах присутствуют актин, миозин и тропомиозин, но тропониновую систему содержат только поперечнополосатые мышцы позвоночных. Следовательно, механизмы регуляции сокращения в разных сократительных системах должны различаться.
Рис. 56.8. Кальциевая регуляция сокращения гладких мышц. (Adapted from Adelstein R. S., Eiscnberg R. Regulation and kinetics of actin myosin ATP interaction. Annu. Rev. Biochem., 1980, 49, 921.)
миозина с F-актином. Таким образом, начинается сократительный цикл (рис. 56.8).
В табл. 56.1 суммируются и сравниваются данные о регуляции актин - миозинового взаимодействия (активации миозиновой АТРазы) в поперечнополосатых и гладких мышцах.
Таблица 56.1. Актин-миозиновые взаимодействия в поперечнополосатых и гладких мышцах
Фосфорилирование легкой цепи миозина гладких мышц снимает ее ингибиторное влияние на взаимодействие актина с миозином и тем самым запускает сократительный цикл. Таким образом, для начала взаимодействия актина с миозином в гладких мышцах требуется фосфорилирование.
Некоторые новые данные свидетельствуют о том, что фосфорилирование тяжелых цепей миозина служит необходимым условием для их сборки в толстые филаменты в скелетных мышцах, гладких мышцах и немышечных клетках (см. ниже).
Фосфагены, такие как креатинфосфат, предотвращают быстрое истощение запасов АТР, поставляя легко используемый макроэргический фосфат, необходимый для ресинтеза АТР из ADP. Креатинфосфат образуется из АТР и креатина в период расслабления мышцы, когда потребность в АТР не столь велика. Фосфорилирование креатина катализируется креатинфосфокиназой (КФК) — специфичным для мышц ферментом, который используется при диагностике осгрых или хронических мышечных нарушений.
АТР в мышечной ткани образуется и в ходе окислительного фосфорилирования — процесса, требующего постоянного притока кислорода. Мышцы, характеризующиеся высокой потребностью в кислороде в связи с длительным состоянием сокращения (например, для поддержания определенной позы), обладают способностью резервировать кислород в миоглобине (см. гл. 6). Поскольку кислород связывается в миоглобине с гемом, мышцы, содержащие миоглобин, окрашены в красный цвет в отличие от не содержащих его белых скелетных мышц. В табл. 56.2 приведены сравнительные данные о некоторых свойствах быстрых (или белых) и медленных (или красных) скелетных мышц.
Миоаденилаткиназа—фермент, присутствующий в мышцах, катализирует образование одной молекулы АТР и одной молекулы АМР из двух молекул ADP. Эта реакция, показанная на рис. 56.9, сопряжена с гидролизом АТР миозиновой АТРазой во время мышечного сокращения. На рисунке, кроме того, отражена связь между различными источниками АТР и его потреблением в ходе сокращения мышцы.
У людей главным после жира источником запасенной энергии служит белок скелетных мышц. Это объясняет очень большую потерю мышечной массы (особенно у взрослых людей), наблюдающуюся при длительной калорической недостаточности.
Изучению распада тканевого белка in vivo препятствует тот факт, что аминокислоты, высвобождающиеся при внутриклеточной деградации белков, могут в значительной степени реутилизироваться для синтеза белка в клетке или переноситься к другим органам и вступать там в анаболические процессы.
Таблица 56.2. Характеристика быстрых и медленных скелетных мышц
Однако актин и миозин после синтеза их пептидных связей метилируются с образованием З-метнлгаспщшш. В ходе внутриклеточного распада актина и миозина 3-метилгистидин высвобождается и выделяется с мочой. При введении метки крысам или человеку было показано, что экскреция с мочой метилированной аминокислоты служит надежным показателем скорости деградации белка миофибрилл в мышцах. Фракционная скорость распада мышечного белка у пожилых людей мало отличается от этого показателя у молодых, но, поскольку масса мышц при старении уменьшается, снижается и вклад этой ткани в общее возрастное увеличение распада белков в организме.
Мышцы также синтезируют и высвобождают большие количества аланина и глутамина. В синтезе этих соединений используются аминогруппы, которые образуются при распаде аминокислот с разветвленной цепью и затем переносятся на а-кетоглутарат и пируват в ходе реакций трансаминирования. Источником почти всего пирувата, идущего на синтез аланина, является гликолиз из экзогенной глюкозы. Эти реакции формируют так называемый глюкозо-аланиновый цикл, в котором аланин мышц используется в процессе печеночного глюконеогенеза и в то же время доставляет в печень аминогруппы, удаляемые в виде мочевины.
Углеродный скелет аминокислот, подвергшихся деградации и включившихся в цикл трикарбоновых кислот в мышечной ткани, превращается главным образом в глутамин и пируват, который далее окисляется или превращается в лактат. Таким образом, при голодании или в период после всасывания большая часть образующихся в процессе распада мышечного белка аминокислот покидает мышцы; исключением являются изолейцин, валин, глутамат, аспартат и аспарагин: они участвуют в образовании глутамина, который высвобождается мышцами и используется другими тканями.
Давно известно, что работающая мышца высвобождает аммиак. Как установлено в настоящее время, непосредственным источником аммиака в скелетной
Рис. 56.9. Множественные источники АТР в мышце.
мышце служит АМР, который дезаминируется в IMP под действием аденилатдезаминазы. IMP может вновь превращаться в АМР в ходе реакций, использующих аспартат и катализируемых аденил-сукцинатсинтетазой и аденилсукциназой (см. гл. 35).
СИЛА, РАБОТА И УТОМЛЕНИЕ СКЕЛЕТНЫХ МЫШЦ
Сила мышц может быть максимальной, относительной и абсолютной. Максимальной силой мышцы называют то напряжение мышцы, которое она может развить в условиях изометрического сокращения (развитие напряжения без изменения длины) или поднятия максимально возможного груза. Относительная сила мышцы — это максимальная сила мышцы, разделенная на площадь ее поперечного сечения. Абсолютная сила мышц — это отношение максимальной силы мышцы к сумме поперечных сечений всех ее волокон. Сила сокращения скелетных мышц зависит от силы раздражения.
Скелетные мышцы способны выполнять определенную работу, которая может быть динамической и статической. При динамической работе происходит сдвиг костей относительно друг друга и мышца перемещает груз. Динамическая работа мышцы определяется путем умножения массы груза на высоту его подъема. При статической работе груз не перемещается, не происходит сдвига костей относительно друг друга, мышца при этом развивает напряжение без изменения длины. В результате этой работы происходит активная фиксация органов относительно друг друга и тело занимает определенное положение в пространстве. Максимальную работу мышца производит при средних нагрузках и среднем ритме работы.
Утомлением называется временное понижение или прекращение работоспособности мышцы в результате ее деятельности. При этом снижаются ее физиологические свойства: возбудимость, лабильность, проводимость, сократимость и др. После отдыха мышца снова сокращается нормально.
Причина утомления еще полностью не выяснена, и существует несколько гипотез, объясняющих возникновение этого явления. Первая гипотеза предполагает, что в результате накопления кислых продуктов обмена веществ (угольной, молочной и фосфорной кислот) наступает отравление мышцы. Согласно второй гипотезе в мышце происходит истощение запасов энергетических ресурсов (гликогена, креатинфосфата, АТФ). Тем не менее в первую очередь утомление наступает в местах контактов нервов и мышц (синапсах). Для того чтобы мышца работала без утомления, необходима ее тренировка, которая способствует лучшему кровоснабжению мышцы, экономному расходованию энергетических запасов и нарастанию массы мышцы.
Деятельность мышц регулируется нервной системой рефлекторно. По чувствительным нейронам импульсы от различных рецепторов передаются в центральную нервную систему, где происходит их анализ и синтез; в результате формируется программа действия, которая по двигательным нейронам поступает к скелетным мышцам. Аксон двигательного нейрона разветвляется, и каждая его веточка иннервирует одно мышечное волокно. Следовательно, один мотонейрон иннервирует группу мышечных волокон. Этот комплекс называется моторной единицей. В мышце имеется большое число моторных единиц. Импульсы к ним подходят не одновременно, поэтому их сокращения по времени не совпадают (одни волокна сокращаются раньше, другие — позже). Этим объясняется плавность сокращения всей мышцы.
В регуляции деятельности мышц участвует головной мозг. Мозжечок осуществляет безусловно-рефлекторную координацию движений (согласованную работу мышц и движения частей тела). Средний и продолговатый мозг обеспечивают перераспределение тонуса мышц при изменении позы. Кора больших полушарий обеспечивает произвольные движения, которые приобретаются в процессе всей жизни человека. Двигательная зона коры расположена в лобной доле впереди от центральной борозды. Из центральной нервной системы к мышцам постоянно поступают импульсы, которые обеспечивают их тонус, т. е. незначительное постоянное напряжение.
1. Данилова Н.Н., Крылова А.Л. Физиология высшей нервной деятельности. – М., 1997. – 432 с.
2. Федюкович Н.И. Анатомия и физиология человека. – Ростов- на- Дону, 2000. – 416 с.
3. Фомин Н.А. Физиология человека. – М.: Просвещение, 1982. – 320 с.
4. Основы физиологии. – М., 1984. - 556 с.
5. Российский физиологический журнал им. И. М. Сеченова. - СПб. М. : Наука.
Читайте также:
