
Нервное волокно его структура и функции перерождение после перерезки
Нервн. волокна (НВ) - это отростки нейронов, с помощью кот. осуще-ся связь между нейронами, а также нейронов с исполнит-ми кл. В состав НВ входят осевой цилиндр (нервный отросток) и глиальная оболочка. По взаимоотнош. осевых цилиндров с глиальными кл выделяют два типа нервных волокон: безмиелиновые и миелиновые. Оболочку безмиелиновых волокон образ шванновские клетки (леммоциты). При этом осевые цилиндры прогибают клеточную оболочку леммоцитов и погружаются в них. Клеточная мембрана обычно полностью окруж. каждый осевой цилиндр и смыкается над ним, образуя сдвоенную мембрану (мезаксон).
Оболочку миелиновых волокон образуют в периферич. нервной системе также шванновские клетки, а в ЦНС — олигодендроциты. В отличие от безмиелиновых волокон в миелиновых волокнах мезаксон удлиняется и спирально закручивается вокруг осевого цилиндра, образуя слой миелина (липидный футляр) вокруг осевого цилиндра. Миелиновая оболочка через равные уч-ки прерыв-ся, образ. свободн. от миелина небольшие уч-ки — узловые перехваты Ранвье. Участки волокон между перехватами наз-ся межузловыми сегментами, они образованы слоем миелина.Миелин сост. на 78 % из липидов (фосфолипид 42 %, цереброзидов — 28 %, холестерина — 25 %.) НО миелин требует затраты энергии и обеспеч. О2 и пит. в-вами(или - деструкция :( ). Нервные волокна имеют две основные ф-ции — изолирующ. ф-ция, способствующ. лучшему проведению биопотенциалов по отросткам нейронов. и транспорт в-в, обеспеч. трофическую ф-цию.
Проведение возбужд. по нервам подчиняется следующим законам: 1. Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезе, вторая - действии веществ блокирующих проведение, например новокаина. 2. Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаше всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным - от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное или антидромное распространение возбуждения. 3. Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола. 4. Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются, проходя по ним. 5. Скорость проведения прямопропорциональна диаметру нерва. (Нервные волокна обладают свойствами центрического кабеля, у которого не очень хорошая изоляция.
40. Механизм проведения потенциала действия в мякотных и безмякотных волокнах. Неутомляемость нервного волокна. Скорость проведения в различных нервах.
Функцию быстрой передачи возбуждения к нервной клетке и от нее выполняют ее отростки - дендриты и аксоны, т.е нервные волокна. В зависимости от структуры их делят на мякотные, имеющие миелиновую оболочку, и безмякотные. Эта оболочка формируется шванновскими клетками, являющиеся видоизмененными глиальными клетками. Они содержат миелин, который в основном состоит из липидов. Он выполняет изолирующую и трофическую функции. Участки, где оболочка прерывается, т.е. не покрыты миелином называют перехватами Ранвье. Функционально все нервные волокна делят на три группы:
1 Волокна типа А - это толстые волокна, имеющие миелиновую оболочку. В эту группу входят 4 подтипа: (альфа) двигательные волокна скелетных мышц и афферентные нервы (рецепторов растяжения). Скорость проведения по ним максимальна (бета) - афферентные волокна, идущие от рецепторов давления и прикосновения кожи. (гамма)- эфферентные волокна, идущие к мышечным веретенам (15-30 м/сек). (сигма) - афферентные волокна от температурных и болевых рецепторов кожи (12-30 м/сек).
2 Волокна группы В - тонкие миелинизированные волокна, являющиеся преганглионарными волокнами вегетативных эфферентных путей.
3 Волокна группы С, безмиелиновые постганглионарные волокна вегетативной нервной системы.
Проведение возбуждения по нервам подчиняется следующим законам: 1. Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезе, вторая - действии веществ, блокирующих проведение, например новокаина. 2. Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаше всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным - от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное или антидромное распространение возбуждения. 3. Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола. 4. Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются, проходя по ним.
5. Скорость проведения прямопропорциональна диаметру нерва. (Нервные волокна обладают свойствами центрического кабеля, у которого не очень хорошая изоляция. В основе механизма проведения возбуждения лежит возникновение местных токов: В результате генерации ПД в аксоном холмике и реверсии мембранного потенциала, мембрана аксона приобретает противоположный заряд. Снаружи она становится отрицательной, внутри положительной. Мембрана нижележащего, невозбужденного участка аксона заряжена противоположным образом. ''Поэтому между этими участками, по наружной и внутренней поверхностям мембраны начинают проходить местные, токи. Эти токи деполяризуют мембрану нижележащего невозбужденного участка нерва до критического уровня к в нем также генерируется ПД. Затем процесс повторяется и возбуждается более отдаленный участок нерва и т.д. Т.к. по мембране безмякотного волокна местные токи текут не прерываясь, поэтому такое проведение называется непрерывным. При непрерывном проведении местные токи захватывают большую поверхность волокна, поэтому им " требуется, длительное время для прохождения по участку волокна. В результате дальность, и. скорость проведения возбуждения по безмякотным волокнам небольшая. В мякотных волокнах участки, покрытые миелином обладают большим электрическим сопротивлением. Поэтому непрерывное проведение ПД Невозможно. При генерации ПД местные токи текут лишь между соседними, перехватами По закону “все или ничего" возбуждается ближайший к аксонному холмику перехват Ранвье, затем соседний нижележащий перехват и т.д. Такое проведение называется сальтаторным (прыжком). При этом механизме ослабления местных токов не происходит, и нервные импульсы распространяются на большое расстояние и с большой скоростью.
Скорость проведения потенциала действия в разных нервах. Скорость потенциала действия нервного ствола. Химические изменения в нерве при потенциале действия. Теплопродукция и утомление.
Энергозатраты нервного волокна на проведение ПД относительно невелики, поскольку возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% общей поверхности мембраны аксона. Поэтому даже после длительных ритмических пачек ПД трансмембранный градиент концентраций ионов практически не изменяется.
В физиологических условиях ПД движутся в одном направлении от места раздражения (ортодромное проведение). ПД, проходящий по нервному волокну, возбуждает следующий, но не предыдущий участок мембраны. Это связано с рефрактерностью предыдущего участка после возбуждения. Проведение в противоположном направлении (антидромное проведение) возможно при травматическом поражении нервных волокон и в редких случаях (аксон–рефлекс).
В периферической нервной системе волокна объединены с помощью соединительнотканных оболочек в нервные стволы (нервы). В одном нерве могут быть тысячи нервных волокон: например, в срединном и мышеч-но-кожном нервах имеется 27—37 тыс. нервных волокон. Волокна в нервах могут быть миелиновыми и безмиелиновыми, афферентными и эфферентными. В естественных условиях каждое волокно нерва возбуждается от своего источника (например, эфферентное — от аксонного холмика, афферентное — от рецептора), и ПД в них проводятся асинхронно. Кроме того, чувствительные и двигательные волокна проводят импульсы в противоположных направлениях. Суммарная электрическая активность нерва создается электрической активностью составляющих его волокон и зависит от числа возбужденных волокон, степени шунтирования местных токов невозбужденными волокнами, синхронности проведения ПД в волокнах. В связи с этим анализ суммарной электрической активности нерва (нейрограммы) представляет трудную задачу.
3. Большая скорость проведения возбуждения. Скорость проведения ПД в различных типах волокон нерва равна 0,5—120 м/с (см. табл. 5.2). Она значительно выше в миелиновых волокнах в связи с сальтаторным типом проведения ПД (см. раздел 5.2.3), а среди ми-елиновых волокон прямо пропорциональна диаметру волокна. Скорость проведения возбуждения в миелиновых нервных волокнах значительно выше, чем в других удлиненных возбудимых структурах, — в гладких миоци-тах (0,02—0,10 м/с), рабочих кардиомиоцитах (около 1 м/с), и только в миоцитах проводящей системы сердца и скелетных миоцитах скорость проведения ПД (2—5 м/с) достигает величин распространения ПД в низкоскоростных нервных волокнах (тип С и В). Передача возбуждения по нервным волокнам является наиболее скоростным из известных способов передачи информации на значительные расстояния в организме. Для сравнения отметим, что скорость передачи гуморальных влияний ограничена скоростью кровотока, которая равна от 0,5 мм/с в капиллярах до 0,25 м/с в аорте (средняя скорость).
Дата добавления: 2015-12-15 | Просмотры: 2168 | Нарушение авторских прав
Нервные волокна не могут существовать вне связи с телом нервной клетки: перерезка нерва ведет к гибели тех волокон, которые оказались отделенными от тела клеток. У теплокровных животных уже через 2—3 сут после перерезки нерва периферический его отросток утрачивает способность к проведению нервных импульсов. Вслед за этим начинается дегенерация нервных волокон, причем миёлиновая оболочка претерпевает жировое перерождение, Это выражается в том, что мякотная оболочка теряет миелин, который скапливается в виде капель; распавшиеся волокна и их миелин рассасываются и на месте нервных волокон остаются тяжи, образованные леммоцитом (шванновской клеткой). Все эти изменения впервые были описаны английским врачом Валлером и названы по его имени валлеровским перерождением.
Регенерация нерва происходит очень медленно. Леммоциты, оставшиеся на месте дегенерировавших нервных волокон, начинают разрастаться вблизи места перерезки по направлению к центральному отрезку нерва. Одновременно перерезанные концы аксонов центрального отрезка образуют так называемые колбы роста — утолщения, которые растут в направлении периферического отрезка. Часть этих веточек попадает в старое ложе перерезанного нерва и продолжает расти в этом ложе со скоростью 0,5—4,5 мм в сутки до тех пор, пока не дойдет до соответствующей
периферической ткани или органа, где волокна образуют нервные окончания. С этого времени восстанавливается нормальная иннервация органа или ткани.
В различных органах восстановление функции после перерезки нерва наступает в разные сроки. В мышцах первые признаки восстановления функций могут появиться через 5—6 нед; окончательное восстановление происходит много позднее, иногда через год.
ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ
При "изучении проведения возбуждения по нерву было установлено несколько необходимых условий и правил (законов) протекания этого процесса.
Анатомическая и физиологическая непрерывность волокна. Проведение импульсов возможно лишь при условии анатомической целостности волокна, поэтому как перерезка нервных волокон, так и любая травма поверхностной мембраны нарушают проводимость. Непроводимость наблюдается также при нарушении физиологической целостности волокна (блокада натриевых каналов возбудимой мембраны тетродотоксином или местными анестетиками, резкое охлаждение и т. п.). Проведение нарушается и при стойкой деполяризации мембраны нервного волокна ионами К, накапливающимися при ишемии в межклеточных щелях. Механическая травма, сдавливание нерва при воспалительном отеке тканей могут сопровождаться частичным или полным нарушением функции проведения.
Двустороннее проведение. При раздражении нервного волокна возбуждение распространяется по нему и в центробежном, и в центростремительном направлениях. Это доказывается следующим опытом.
К нервному волокну, двигательному или чувствительному, прикладывают две пары электродов, связанных с двумя электроизмерительными приборами А и Б (рис. 45). Раздражение наносят между этими электродами. В результате двустороннего проведения возбуждения приборы зарегистрируют прохождение импульса как под электродом А, так и под электродом Б.
Двустороннее проведение не является только лабораторным феноменом. В естественных условиях потенциал действия нервной клетки возникает в той ее части, где тело переходит в ее отросток — аксон (так называемый начальный сегмент). Из начального сегмента потенциал действия распространяется двусторонне: в аксоне по направлению к нервным окончаниям и в тело клетки по направлению к ее дендритам.
Изолированное проведение. В периферическом нерве импульсы распространяются по каждому волокну изолированно, т. е. не переходя с одного волокна на другое и оказывая действие только на те клетки, с которыми контактируют окончания данного нервного волокна. Это имеет очень важное значение в связи с тем, что всякий периферический нервный ствол содержит большое число нервных волокон — двигательных, чувствительных и вегетативных, которые иннервируют разные, иногда далеко отстоящие друг от друга и разнородные по структуре и функциям клетки и ткани. Например, блуждающий нерв иннервирует все органы грудной полости и значительную часть органов брюшной полости, седалищный нерв — всю мускулатуру, костный аппарат, сосуды и кожу нижней конечности. Если бы возбуждение переходило внутри нервного ствола с одного волокна на другое, то в этом случае нормальное функционирование периферических органов и тканей было бы невозможно.
Изолированное проведение в отдельных волокнах смешанного нерва может быть доказано простым опытом на скелетной мышце, иннервированной смешанным нервом, в образовании которого участвует несколько спинномозговых корешков. Если раздражать один из этих корешков, сокращается не вся мышца, как это было бы в случае перехода возбуждения с одних нервных волокон на другие, а только те группы мышечных волокон, которые иннервированы раздражаемым корешком. Еще более строгое доказательство изолированного проведения возбуждения может быть получено при отведении потенциалов действия от различных нервных волокон нервного ствола.
Изолированное проведение нервного импульса обусловлено тем, что сопротивление жидкости, заполняющей межклеточные щели, значительно ниже сопротивления мем-
Рис. 45. Схематическое изображений опыта для доказательства двустороннего проведения импульса и нерве. Объяснение и тексте.
браны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным (деполяризованным) и покоящимися участками возбудимой мембраны, проходит по межклеточным щелям, не заходя в соседние волокна.
Нервн. волокна (НВ) - это отростки нейронов, с помощью кот. осуще-ся связь между нейронами, а также нейронов с исполнит-ми кл. В состав НВ входят осевой цилиндр (нервный отросток) и глиальная оболочка. По взаимоотнош. осевых цилиндров с глиальными кл выделяют два типа нервных волокон: безмиелиновые и миелиновые. Оболочку безмиелиновых волокон образ шванновские клетки (леммоциты). При этом осевые цилиндры прогибают клеточную оболочку леммоцитов и погружаются в них. Клеточная мембрана обычно полностью окруж. каждый осевой цилиндр и смыкается над ним, образуя сдвоенную мембрану (мезаксон).
Оболочку миелиновых волокон образуют в периферич. нервной системе также шванновские клетки, а в ЦНС — олигодендроциты. В отличие от безмиелиновых волокон в миелиновых волокнах мезаксон удлиняется и спирально закручивается вокруг осевого цилиндра, образуя слой миелина (липидный футляр) вокруг осевого цилиндра. Миелиновая оболочка через равные уч-ки прерыв-ся, образ. свободн. от миелина небольшие уч-ки — узловые перехваты Ранвье. Участки волокон между перехватами наз-ся межузловыми сегментами, они образованы слоем миелина.Миелин сост. на 78 % из липидов (фосфолипид 42 %, цереброзидов — 28 %, холестерина — 25 %.) НО миелин требует затраты энергии и обеспеч. О2 и пит. в-вами(или - деструкция :( ). Нервные волокна имеют две основные ф-ции — изолирующ. ф-ция, способствующ. лучшему проведению биопотенциалов по отросткам нейронов. и транспорт в-в, обеспеч. трофическую ф-цию.
Проведение возбужд. по нервам подчиняется следующим законам: 1. Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезе, вторая - действии веществ блокирующих проведение, например новокаина. 2. Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаше всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным - от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное или антидромное распространение возбуждения. 3. Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола. 4. Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются, проходя по ним. 5. Скорость проведения прямопропорциональна диаметру нерва. (Нервные волокна обладают свойствами центрического кабеля, у которого не очень хорошая изоляция.
Механизм проведения потенциала действия в мякотных и безмякотных волокнах. Неутомляемость нервного волокна. Скорость проведения в различных нервах.
Функцию быстрой передачи возбуждения к нервной клетке и от нее выполняют ее отростки - дендриты и аксоны, т.е нервные волокна. В зависимости от структуры их делят на мякотные, имеющие миелиновую оболочку, и безмякотные. Эта оболочка формируется шванновскими клетками, являющиеся видоизмененными глиальными клетками. Они содержат миелин, который в основном состоит из липидов. Он выполняет изолирующую и трофическую функции. Участки, где оболочка прерывается, т.е. не покрыты миелином называют перехватами Ранвье. Функционально все нервные волокна делят на три группы:
1 Волокна типа А - это толстые волокна, имеющие миелиновую оболочку. В эту группу входят 4 подтипа: (альфа) двигательные волокна скелетных мышц и афферентные нервы (рецепторов растяжения). Скорость проведения по ним максимальна (бета) - афферентные волокна, идущие от рецепторов давления и прикосновения кожи. (гамма)- эфферентные волокна, идущие к мышечным веретенам (15-30 м/сек). (сигма) - афферентные волокна от температурных и болевых рецепторов кожи (12-30 м/сек).
2 Волокна группы В - тонкие миелинизированные волокна, являющиеся преганглионарными волокнами вегетативных эфферентных путей.
3 Волокна группы С, безмиелиновые постганглионарные волокна вегетативной нервной системы.
Проведение возбуждения по нервам подчиняется следующим законам: 1. Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезе, вторая - действии веществ, блокирующих проведение, например новокаина. 2. Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаше всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным - от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное или антидромное распространение возбуждения. 3. Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола. 4. Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются, проходя по ним.
5. Скорость проведения прямопропорциональна диаметру нерва. (Нервные волокна обладают свойствами центрического кабеля, у которого не очень хорошая изоляция. В основе механизма проведения возбуждения лежит возникновение местных токов: В результате генерации ПД в аксоном холмике и реверсии мембранного потенциала, мембрана аксона приобретает противоположный заряд. Снаружи она становится отрицательной, внутри положительной. Мембрана нижележащего, невозбужденного участка аксона заряжена противоположным образом. ''Поэтому между этими участками, по наружной и внутренней поверхностям мембраны начинают проходить местные, токи. Эти токи деполяризуют мембрану нижележащего невозбужденного участка нерва до критического уровня к в нем также генерируется ПД. Затем процесс повторяется и возбуждается более отдаленный участок нерва и т.д. Т.к. по мембране безмякотного волокна местные токи текут не прерываясь, поэтому такое проведение называется непрерывным. При непрерывном проведении местные токи захватывают большую поверхность волокна, поэтому им " требуется, длительное время для прохождения по участку волокна. В результате дальность, и. скорость проведения возбуждения по безмякотным волокнам небольшая. В мякотных волокнах участки, покрытые миелином обладают большим электрическим сопротивлением. Поэтому непрерывное проведение ПД Невозможно. При генерации ПД местные токи текут лишь между соседними, перехватами По закону “все или ничего" возбуждается ближайший к аксонному холмику перехват Ранвье, затем соседний нижележащий перехват и т.д. Такое проведение называется сальтаторным (прыжком). При этом механизме ослабления местных токов не происходит, и нервные импульсы распространяются на большое расстояние и с большой скоростью.

Нервное волокно — это отросток нейрона, который покрыт специальной оболочкой (глиального типа). Благодаря их присутствию у нервной системы человека появилась способность передавать и воспринимать импульсы. При повреждении миелиновой оболочки происходит процесс димиелинизации, который сопровождается тяжелыми заболеваниями. В данной статье пойдет речь о строении этой структуры, их основных функция и значимости для ЦНС человека.
Общие сведения
Все нервы включают в себя огромное количество волокон, которые окружаются соединительной тканью. Само волокно состоит из особого отростка — аксона, который покрыт эктодермальной оболочкой. Они собираются в определенные пучки, таким образом, создаются тракты в головном, спинном мозгу и периферической нервной системе. Стоит отметить, что отростки бывают мякотными и безмякотными (например, нервные окончания кожных покровов).
Все они отличаются по характеру своего покрытия, а также принадлежностью к определенной нервной системе. Разделяются на две основные группы: покрытые миелином и лишенные его. В целом в организме человека преобладает именно первая группа.
Рассмотрим подробнее строение миелинового волокна.
Его основными компонентами являются:
- цилиндр, которой проходит по центральной оси;
- непосредственно оболочка миелиновой природы, которая покрывает осевой цилиндр;
- шванновская оболочка.

Классификация нервных волокон
Все они по классификации разделяются на три основные группы:
- по скорости передачи импульса;
- по поперечному диаметру;
- по продолжительности потенциала действия.
Стоит отметить, что, чем больше будет их диаметр и миелинизация, тем быстрее по нему проходит импульс. Выделяют три разновидности:
- Группа А. Все они покрыты оболочкой, потенциал действия у них самый низкий. В свою очередь они разделяются на 4 подвида: альфа, бета, гамма и дельта. К ним относятся все рецепторы соматической нервной системы, чувствительные волокна кожи, терморегуляции, проприорецепторов. Все эти отростки отвечают за тактильные чувства человека.
- Группа В. Отростки не полностью покрыты миелиновой оболочкой, к ним относятся составляющие вегетативной нервной системы. Сюда относятся медиаторы болевых ощущений и сигнализаторы работы внутренних органов.
- Группа С. Оболочка полностью отсутствует, скорость проведения импульса низкая. К ним относятся клетки ВНС, а также болевые и температурные соматические.
В состав миелина входят фосфолипиды, холестерол, основное белковое вещество и другие полезные компоненты. Таки образом оболочка является уникальной мембраной, благодаря которой в нервной системе появляется возможность быстрой передачи импульсов.
Все нервные отростки делят на две основные группы: афферентные (проводят импульсы от тканей до ЦНС) и эфферентные (действуют наоборот).

Миелинизация нервных волокон и демиелинизация
Как описывалось выше, каждый отросток имеет в своем составе осевой цилиндр, который покрыт специальной миелиновой оболочкой. Этот процесс носит название миелинизации. Благодаря присутствию участков Ранвье происходит передача импульса от одного к другому. Именно это и обеспечивает высокую передачу возбуждения по отростку в направлении к нерву.
В промежутках Ранвье происходит генерация и ускорение импульсных реакций. Их функции в вегетативной нервной системе берут на себя олигодендроглии.
Ткани безмякотного характера не имеют миелиновой оболочки в своем составе, поэтому характеризуются низкой изоляционной способностью. В данном случае скорость передачи импульса значительно снижается из-за того, что при его передаче от нейронов, он напрямую контактирует с окружающей средой. Передача импульсов для них требует больших энергетических затрат организма (в отличие от волокон мякотного типа).
Из этих двух групп волокон в дальнейшем формируется крупный нерв, который имеет на своем окончании в виде мелких пучков. Они отличаются по своим основным функциям. Важно отметить, что данные участки являются конечными при формировании межнейронной системы.
При нарушении функционирования миелиновой оболочки или ее повреждении происходит процесс димиелинизации. Данная патология может быть вызвана наличием воспалительного или инфекционного процесса в организме, нарушениями метаболизма, ишемическими процессами в тканях или распространением нейроинфекции. В результате этого процесса происходит замена миелина в оболочке на фиброзные бляшки. Проводимость импульсных реакций в таком случае значительно снижается.
Существует два вида димиелинизации:
- миелинопатия, которая является результатом аутоиммунных нарушений в организме;
- миелинокластия появляется при генетической предрасположенности к процессу димиелинизации.
Данный процесс считается достаточно опасным, так как несет серьезные нарушения в работе ЦНС. Очень важно диагностировать заболевание на ранней стадии, чтобы провести эффективную терапию.
Функции нервных волокон
Основной функцией нервных отростков является передача импульсной реакции от нейрона к нейрону. Существует два вида такой передачи:
- импульсная. В ее основе лежат электролитные и нейтротрансмиттерные механизмы. Как описывалось выше, в волокнах, покрытых миелиновой оболочкой скорость передачи намного выше;
- безимпульсная. Все реакции происходят за счет тока аксоплазмы с использованием микротрубочек аксона. Последние содержат в своем составе специальное вещество, которое оказывает трофическое воздействие на иннервирующий орган.
Во время передачи импульса происходит трансформация электрических потенциалов, в результате которых образуется уникальные молекулы — нейромедиаторы.
Все данное образование обладают уникальными свойствами:
- лабильность (за определенное время может проводиться ограниченное количество импульсов);
- возбудимость;
- проводимость.
Считается, что нервное волокно неутомлямо. Это связано с низкими затратами АТФ при передаче импульсной реакции. В случае безмиелиновых волокон энергии требуется в разы больше, поэтому и скорость передачи значительно снижается.

Заключение
Итак, нервное волокно — это отдельный отросток нейрона, бывают с миелиновой оболочкой или без нее. Основной их функцией является передача импульса по нейронам к основному нерву. Основными составляющими ПНС и ЦНС являются именно миелиновые волокна, в ВНС преобладают безмиелиновые. В зависимости от сигнала, который проходит по волокну различают чувствительные, двигательные вегетативные и соматические. В случае нарушения функционирования миелина или повреждения оболочки у человека диагностируют серьезные патологии. Они требует своевременной диагностики и лечения.
При повреждении нервов дегенеративные процессы аксона происходят дистальнее места поражения. Это связано с тем, что аксон является отростком нейрона, и его питание и рост зависят от сомы. Регенеративные процессы периферической нервной системы характеризуются высокой интенсивностью и в большинстве случае обеспечивают полное восстановление структур. В ЦНС, напротив, интенсивность процессов восстановления не высока, и регенерация не является полной.
а) Валлерова (антероградная) дегенерация периферических нервов. Основные этапы процесса дегенерации и их описание представлены на рисунке ниже и в подписи к нему. После разрушения или перерезки нерва в течение первых 48 ч аксоны и миелиновая оболочка под действием протеаз, высвобождающихся из шванновских клеток под влиянием ионов Са 2+ , распадаются на фрагменты элипсоидной формы дистальнее места нарушения целостности нерва. Продукты распада удаляют моноциты, которые перемещаются из кровотока в эндоневрий и превращаются в макрофаги.
Помимо осуществления фагоцитоза, макрофаги стимулируют митотическую активность шванновских клеток. Макрофаги и шванновские клетки выполняют трофическую (питательную) и направляющую функции для регенерирующих аксонов.
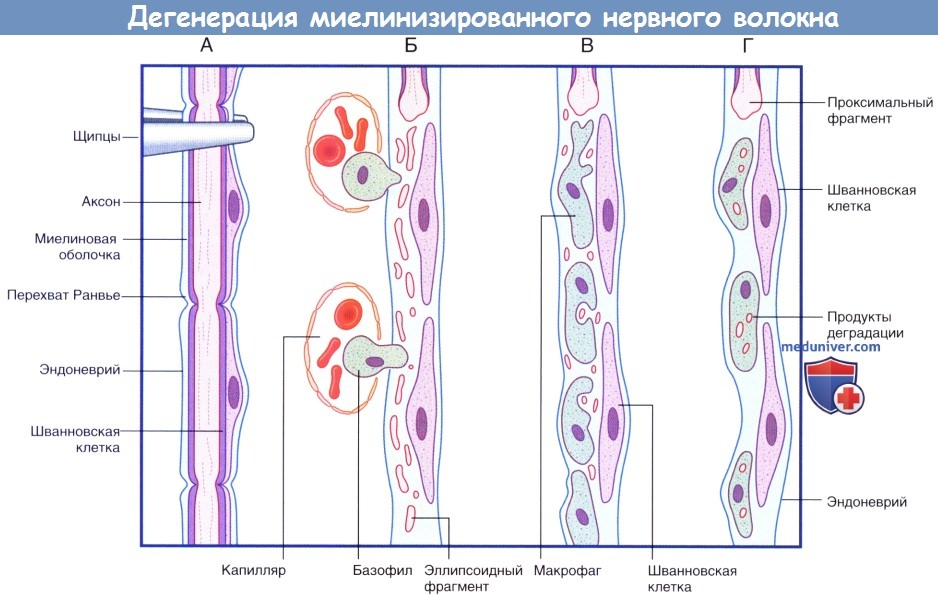
Последовательность процессов при дегенерации миелинизированного нервного волокна.
(А) Сохранное нервное волокно; продемонстрированы четыре сегмента. Выполняют пережатие нервного волокна в верхней части.
(Б) Миелиновая оболочка и аксон распадаются на мелкие частицы и фрагменты эллипсоидной формы. Моноциты проникают в эндоневрий из кровеносного русла.
(В) Мелкие продукты дегенерации поглощают моноциты.
(Г) Происходит практически полное удаление всех продуктов дегенерации. Шванновские клетки и эндоневрий сохранны. Последовательность процессов при регенерации миелинизированного нервного волокна. 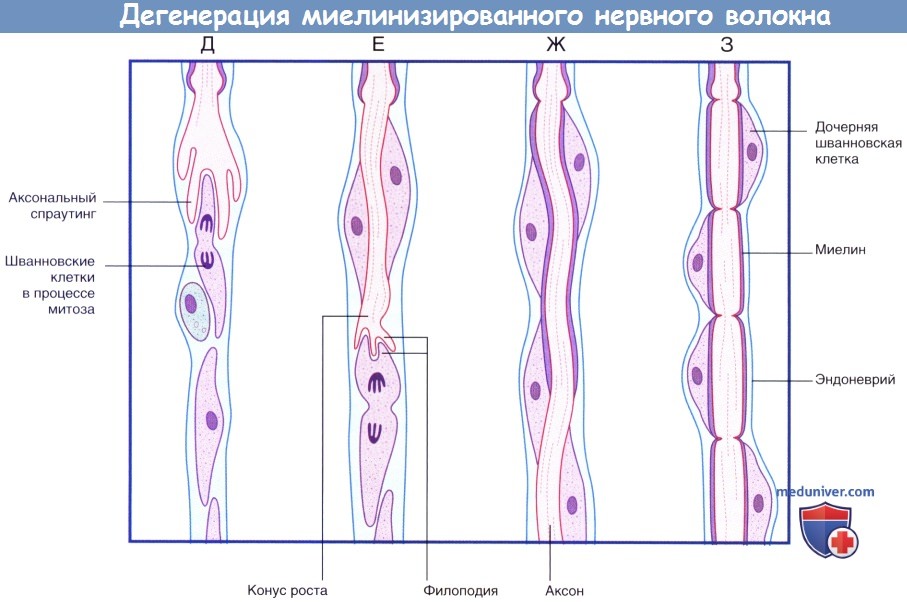
(продолжение) Последовательность процессов при дегенерации миелинизированного нервного волокна.
(Д) Аксональный спраутинг начинается с дистального участка аксона. Спраутинг оказывает митогенное дейсвтие на окружающие шванновские клетки.
(Е) Конус роста продвигается дистально вдоль поверхности шванновских клеток.
(Ж) Миелинизация начинается вдоль проксимального участка регенерирующего аксона.
(З) Общее строение вновь миелинизированного нервного волокна соответствует тем же принципам, однако миелинизированные сегменты характеризуются меньшей длиной.
б) Регенеративные процессы в периферической нервной системе. Основные этапы процесса регенерации представлены на рисунке ниже. После ровного среза нерва спраутинг (разрастание новых ветвей) на конце проксимального отрезка аксона начинается уже спустя несколько часов. Однако в клинической практике повреждения нерва часто происходят при раздавливании или разрыве. В этих случаях происходит отмирание участка нерва длиной 1 см и более, за счет чего спраутинг может продолжаться в течение недели. В случае удачной регенерации происходит тесное соприкосновение проксимального конца аксона со шванновской клеткой дистального конца пересеченного нейрона.
При нарушении формирования этой связи в месте первичного повреждения образуется псевдоневрома, представляющая собой извитые регенерирующие аксоны, погруженные в рубцовую ткань. Ампутационные псевдоневромы — источники сильных болей после ампутаций конечностей.
Регенерация нейронов при повреждении происходит двумя путями в течение нескольких часов после повреждения. На проксимальном конце пересеченного аксона появляются множественные отростки, на конце которых образуются утолщения — конусы роста. На дистальном конце шванновские клетки формируют отростки, направляющиеся навстречу конусам роста. На концах конусов роста формируются напоминающие антенны филоподии, где располагаются поверхностные рецепторы, временно связывающиеся с соответствующими поверхностными молекулами адгезии базальных мембран шванновских клеток. Актиновые филаменты филоподий прикрепляются к поверхностным рецепторам и относительно этих соединений осуществляют дальнейшее продвижение конусов роста.
Конусы роста стимулируют митотическую активность шванновских клеток. Шванновские клетки делятся и миелинизируют наиболее крупные аксоны.
При полном разрыве нервных стволов перед попыткой их восстановления, как правило, выжидают около трех недель, поскольку сразу после повреждения их соединительнотканные оболочки отечны, а в течение этого промежутка времени они становятся немного толще, что позволяет шовному материалу лучше закрепиться. Кроме того, обрезание нервов перед наложением швов приводит к развитию вторичной аксотомии проксимальной части пересеченного аксона. В ходе экспериментальных исследований на животных показано, что вторичная аксотомия стимулирует более интенсивную и длительную регенерацию.
Влияния тела нейрона на пересеченный участок нерва заключаются в следующем:
• Вследствие осмотических изменений перикариона ядро ориентируется эксцентрически.
• Клетки нейроглии заполняют все синаптические щели, изолируя исходный двигательный нейрон от синаптических контактов в сером веществе.
• В исследованиях, проведенных на обезьянах, показано, что после перерезки чувствительных нейронов 30-40 % окончаний дорсальных нервных корешков подвергается валлеровской дегенерации. Регенерация этих нейронов не происходит, поскольку их окончания располагаются в сером веществе головного мозга. Однако некоторые рецепторы восстанавливаются за счет спраутинга сохранных прилежащих нейронов. Данное наблюдение позволяет объяснить явление неполного восстановления чувствительности у таких пациентов.
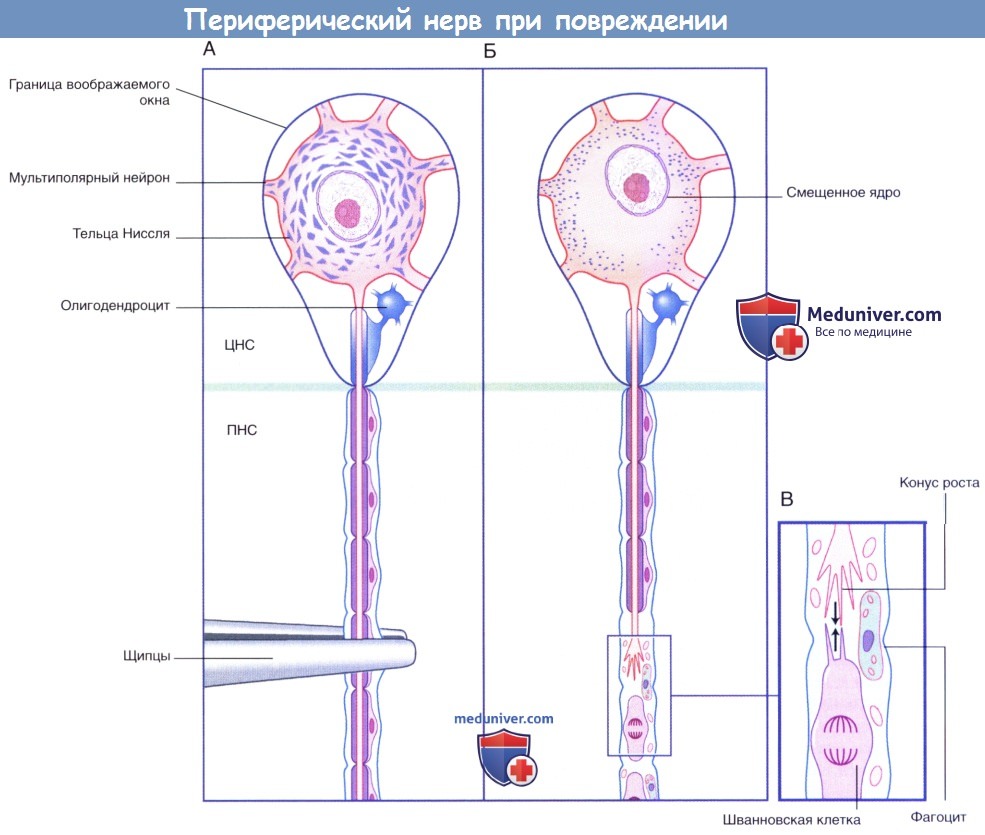
Схематическое изображение процессов, происходящих в периферическом нерве после повреждения.
(А) Двигательный нейрон ЦНС, видимый через воображаемое окно.
(Б) Хроматолизис проявляется фрагментированием и рассеиванием телец Ниссля, а также смещением ядра.
(В) В зоне повреждения в условиях удаления продуктов деградации происходит образование контакта филоподий конуса роста с проксимальными выростами шванновских клеток (указано стрелками).
ЦНС — центральная нервная система; ПНС — периферическая нервная система.
Редактор: Искандер Милевски. Дата публикации: 12.11.2018
Читайте также:
