
Нервные центры и нервные волокна основные свойства нервных центров и нервных волокон
Нервный центр – это совокупность нейронов, обеспечивающих регуляцию какого-либо конкретного физиологического процесса или функции.
Нервный центр в узком смысле – это совокупность нейронов, без которых данная конкретная функция не может регулироваться. Например, без нейронов дыхательного центра продолговатого мозга дыхание прекращается. Нервный центр в широком смысле — это совокупность нейронов, которые участвуют в регуляции конкретной физиологической функции, но не являются строго обязательными для ее осуществления! Например, в регуляции дыхания кроме нейронов продолговатого мозга участвуют нейроны пневмотаксического центра варолиевого моста, отдельные ядра гипоталамуса, кора больших полушарий и другие образования головного мозга.
Все нейроны нервного центра разделяют на 2 неравные по количеству и качеству группы.
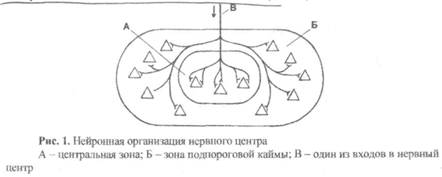
Первая группа – нейроны центральной зоны. Это наиболее возбудимые нейроны, которые возбуждаются в ответ на поступление порогового (для нервного центра) сигнала. Таких нейронов около 15-20%, и они не обязательно располагаются в середине нервного центра, как это изображено на рис.1. Особенностью их является то, что они имеют на своем теле больше синаптических терминалей от сенсорных и вставочных нейронов.
Вторая группа – нейроны подпороговой каймы. Это менее возбудимые нейроны, которые не возбуждаются в ответ на поступление пороговых им-пульсов, но при действии более сильных раздражителей они возбуждаются и включаются в работу нервного центра, обеспечивая ее усиление. Таких нейронов большинство (80-85%), и они не обязательно располагаются на периферии нервного центра, но все имеют значительно меньше синаптических терминалей от сенсорных и вставочных нейронов по сравнению с нейронами центральной зоны.
На рис. 1 нейроны центральной зоны условно поставлены в центр внутреннего круга (А), а нейроны подпороговой каймы – в пространство между внутренним и наружным кругами (Б). Таким образом, если к нервному центру по афферентному входу (В) придет пороговый импульс, то возбудятся три нейрона центральной зоны, а на десяти нейронах подпороговой каймы потенциалы действия не возникнут, но появится местная деполяризация – возбуждающий постсинаптический потенциал (ВПСП).
От структуры нервного центра зависят его свойства, а они, в свою очередь, влияют на процесс проведения возбуждения через нервный центр, на его скорость и степень выраженности. От свойств нервных центров во многом зависит процесс распространения возбуждения по ЦНС, что имеет важное значение в интегративной деятельности организма.
Свойства нервных центров обусловлены описанной выше нейронной организацией нервного центра, а также химическим способом передачи возбуждения в синапсах. При электрическом способе передачи возбуждения нервные центры не имели бы подобных свойств.
Свойства нервных центров: 1 одностороннее проведение возбуждения; 2 задержка проведения возбуждения; 3 суммация; 4 облегчение; 5 окклюзия; 6 мультипликация; 7 трансформация; 8 последействие; 9 посттетаническая потенциация; 10 утомление; 11 тонус; 12 высокая чувствительность к изменению состояния внутренней среды организма; 13 пластичность.
3) Суммация – это возникновение возбуждения в нервном центре при поступлении к нему нескольких допороговых импульсов, каждый из которых в отдельности не может возбуждения (рис. 2). Фактически этот процесс происходит на нейронах подпороговой каймы. Различают два вида суммации: пространственную и временною.
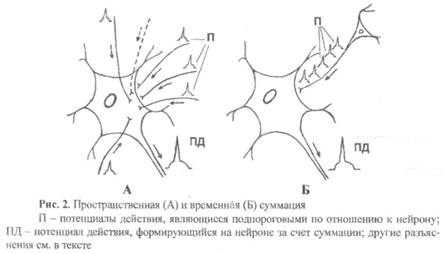
Пространственная суммация возникает в том случае, когда к нервному центру (к его нейронам) приходят одновременно, несколько допороговых импульсов. На рисунке 2А видно, что к нейрону подпороговой каймы, имеющему пороговый потенциал 30 мВ одновременно по пяти различным афферентным входам (их аксоны обозначены сплошной линией) приходят пять импульсов, каждый из которых деполяризует мембрану нейрона на 5 мВ (то есть возникают пять отдельных ВПСП). В этом случае возбуждение нейрона не наступает, так как суммарная деполяризация мембраны нейрона составляет лишь 25 мВ (суммированный ВПСП мал для достижения КУД). Но если к нейрону придет еще один подобный импульс по шестому входу (его аксон обозначен пунктирной линией), то суммированный ВПСП будет достаточен по величине и мембрана нейрона в зоне аксонного холмика деполяризуется до критического уровня, в результате чего нейрон из состояния покоя перейдет в состояние возбуждения. На постсинаптической мембране происходит суммация ВПСП в пространстве.
Временная (последовательная) суммация возникает в том случае, когда к нейронам нервного центра по одному афферентному входу приходит не один, а серия импульсов с очень небольшими по времени межимпульсными промежутками (рис. 2Б). Два механизма временной суммации:
1) интервалы между отдельными импульсами настолько малы, что за это время медиатор, выделившийся в синаптическую щель, не успевает полностью разрушиться и вернуться в пресинаптический аппарат. В этом случае возникает постепенное накопление медиатора до критического объема, необходимого для возникновения достаточного по амплитуде ВПСП, а значит, и для возникновения ПД;
2) интервалы между отдельными импульсами настолько малы, что возникший за это время на постсинаптической мембране ВПСП не успевает исчезнуть и усиливается за счет новой порции медиатора – суммируется. На постсинаптической мембране происходит суммация ВПСП во времени.
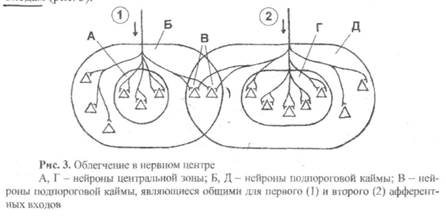
4) Облегчение – это увеличение количества возбужденных нейронов в нервном центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному, а по двум или более афферентным входам. На рис. 3 рассмотрен случай, когда при отдельном раздражении первого афферентного входа возбуждается только три нейрона центральной зоны (А), а на пяти нейронах подпороговой каймы (Б) возникают ВПСП. Если раздражать отдельно только второй афферентный вход, то возбуждены будут пять нейронов (Г), а четыре нейрона подпороговой каймы (Д) не возбудятся. Раздражая и первый, и второй афферентные входы одновременно (!), мы ожидаем вовлечения в процесс возбуждения восьми нейронов. И они, естественно, будут возбуждаться, но кроме них (сверх ожидания!) могут возбуждаться еще некоторые нейроны подпороговой каймы. Это произойдет потому, что один или несколько нейронов подпороговой каймы являются общими как для первого, так и для второго афферентных входов (в нашем случае это два нейрона, обозначенные буквой В), и при одновременном поступлении возбуждения к этим нейронам дни возбудятся за счет возникновения пространственной суммации.
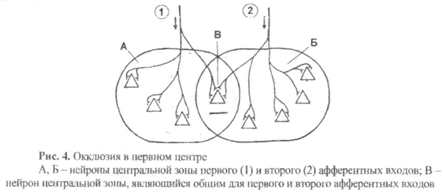
5) Окклюзия – это уменьшение количества возбужденных нейронов в нервной центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному. а по двум или более афферентным входам (рис. 4).
6) Мультипликационное возбуждение (мультипликация) заключается в том, что по разветвлениям аксона вставочного нейрона возбуждение поступает одновременно не на один, а на несколько моторных нейронов (рис. 6). В связи с этим эффект на рабочем органе усиливается в несколько раз, или в работу вовлекаются не одна, а несколько рабочих структур, Это свойство особенно ярко проявляется в ганглиях автономной (вегетативной) нервной системы.
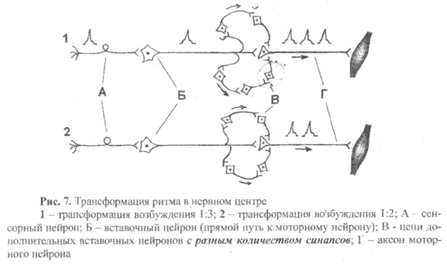
7) Трансформация ритма возбуждения – это изменение частоты импульсов на выходе из нервного центра по сравнению с частотой импульсов на входе в нервный центр.
а) наличием дублирующих цепей вставочных нейронов, связывающих сенсорные и моторные нейроны;
б) разным количеством синапсов в каждой из этих цепей.


Например, на рис.7 представлены два варианта трансформации, которые, на первый взгляд, не отличаются друг от друга, так как в том и в другом случае показаны две дополнительные цепи вставочных нейронов (кроме прямого пути), с помощью которых возбуждение может передаваться по цепи нейронов А—Б—В. Рассмотрим эти схемы.
Вариант 1. Верхняя цепь состоит из двух дополнительных вставочных нейронов, а значит, по сравнению с прямым путем передачи возбуждения с нейрона Б на нейрон В, имеет два дополнительных синапса. Поэтому возбуждение, проходя по верхней цепи, задержится на 2 мс (время синаптической задержки в одном синапсе составляет
1 мс) и придет на нейрон В после того, как пройдет возбуждение по прямому пути. В нижней цепи три дополнительных вставочных нейрона (то есть три дополнительных синапса), значит, возбуждение будет доходить до нейрона В еще дольше, чем по верхней цепи (задержка составит 3 мс). Следовательно, по нижней цепи возбуждение на нейрон В придет после того, как пройдет возбуждение по верхней цепи. В результате на один импульс, пришедший по сенсорному нейрону А, на моторном нейроне В возникнет три потенциала действия (трансформация 1:3).
Вариант 2. В этом случае и верхняя и нижняя цепи вставочных нейронов состоят из двух дополнительных нейронов. Возбуждение по обеим цепям придет к нейрону В одновременно в виде одного потенциала действие, который появится на нейроне В только после прохождения возбуждения к нему от нейрона Б по прямому пути. В этом варианте мы тоже получим трансформацию ритма, но уже в соотношении 1:2.
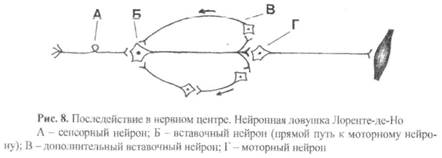
8) Последействие – это продолжение возбуждения моторного нейрона в течение некоторого времени после прекращения действия раздражителя.
Другими причинами последействия могут быть:
а) возникновение высокоамплитудного ВПСП, в результате которого возникает не один, а несколько потенциалов действия то есть ответ длится большее время;
б) длительная следовая деполяризация постсинаптической мембраны, в результате чего возникают несколько потенциалов действия, вместо одного.
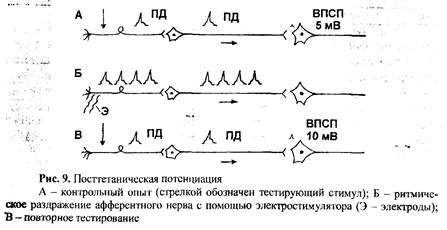
9) Посттетаническая потенциация (синаптическое облегчение) – это улучшение проведения в синапсах после короткого раздражения афферентных путей.
Если в качестве контроля вызвать одиночное раздражение афферентного нерва тестирующим раздражителем (рис. 9А), то на моторном нейроне мы получим ВПСП вполне определенной амплитуды (в нашем случае 5 мВ). Если после этого тот же афферентный нерв раздражать некоторое время серией частых импульсов (рис. 9Б), а потом вновь подействовать тестирующим раздражителем (рис. 9В), то величина ВПСП будет больше (в нашем случае 10 мВ). Причем она будет тем больше, чем более частыми импульсами мы раздражали афферентный нерв.
Длительность синаптического облегчения зависит от свойств синапса и характера раздражения: после одиночных стимулов оно выражено слабо, после раздражающей серии потенциация (облегчение) может продолжаться от нескольких минут до нескольких часов. Объясняется он тем, что при частом раздражении афферентного волокна в его пресинаптической терминали (окончании) накапливаются ионы кальция, а значит, улучшается выделение медиатора. Кроме того, показано, что частое раздражение нерва приводит к усилению синтеза медиатора, мобилизации пузырьков медиатора, к усилению синтеза белков-рецепторов на постсинаптической мембране и увеличению их чувтствительности. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах.
10) Утомление нервного центра (посттетаническая депрессия, синаптическая депрессия) – это уменьшение или прекращение импульсной активности нервного центра в результате длительной стимуляции его афферентными импульсами (или произвольного вовлечения его в процесс возбуждения посредством импульсов, идущих из коры больших полушарий). Причинами утомления нервного центра могут быть:
• истощение запасов медиатора в афферентном или вставочном нейроне;
• снижение возбудимости постсинаптической мембраны (то есть мембраны моторного или вставочного нейрона) из-за накопления, например, продуктов метаболизма.
Утомляемость нервных центров продемонстрировал Н.Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения п. tibialis и п. peroneus. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги. Синаптическая депрессия при длительной активации центра выражается в снижении постсинаптических потенциалов.
11) Тонус нервного центра – это длительное, умеренное возбуждение нервного центра без видимо утомления Причинами тонуса могут быть:
• потоки афферентных импульсов, постоянно поступающие с неадаптирующихся рецепторов;
• гуморальные факторы, постоянно присутствующие в плазме крови;
• спонтанная биоэлектрическая активность нейронов (автоматия);
• циркуляция (реверберация) импульсов в ЦНС.
12) Нервный центр состоит из нейронов, а они очень чувствительны к изменению состава внутренней среды организма, что и отражается на свойствах нервных центров. Наиболее важными факторами, влияющими на работу нервных центров, являются: гипоксия; недостаток питательных веществ (например, глюкозы); изменение температуры; воздействие продуктов метаболизма; воздействие различных токсических и фармакологических препаратов.
Разные нервные центры имеют неодинаковую чувствительность к воздействию названных факторов. Так, нейроны коры больших полушарий наиболее чувствительны к гипоксии, недостатку глюкозы, продуктам метаболизма; клетки гипоталамуса – к изменению температуры, содержанию глюкозы, аминокислот, жирных кислот и др.; различные участки ретикулярной формации выключаются разными фармакологическими препаратами, различные нервные центры избирательно активируются или тормозятся разными медиаторами.
13) Пластичность нервного центра означает его способность изменять при определенных обстоятельствах свои функциональные свойства. В основе этого явления лежит поливалентность нейронов нервных центров. Особенно ярко проявляется это свойство при всевозможных повреждениях ЦНС, когда организм компенсирует утраченные функции за счет сохранившихся нервных центров. Особенно хорошо свойство пластичности выражено в коре больших полушарий. Например, центральные параличи, связанные с патологией двигательных центров коры, иногда полностью компенсируются, и ранее утраченные двигательные функции восстанавливаются.
Дата добавления: 2015-01-21 ; просмотров: 21333 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
ЦЕНТРЫ НЕРВНОЙ СИСТЕМЫ
Функционально связанная совокупность нейронов, расположенных в одной или нескольких структурах ЦНС и обеспечивающих регуляцию той или иной функции или осуществление целостной реакции организма, называется центром нервной системы. Физиологическое понятие центра нервной системы отличается от анатомического представления о ядре, где близко расположенные нейроны объединяются общими морфологическими особенностями.
Классификация нервных центров. В основу классификации центров нервной системы положены морфологический и функциональный критерий. По локализации в структурах нервной системы различают корковые, подкорковые и спинальные центры. В головном мозге также выделяют центры диэнцефальные, мезэнцефальные, бульбарные, гипоталамические, таламические. На функциональной основе центры нервной системы разделяют по регулируемой функции (например, сосудодвигательный центр, центр
теплообразования, дыхательный центр и др.) или по афферентному восприятию (например, центры зрения, слуха, обоняния и др.). Выделяют также центры нервной системы, которые формируют мотивационные состояния организма, являясь пейсмекерами мотивационных возбуждений (центры голода,
жажды, насыщения и др.).
Доминанта. При высоком уровне тонического возбуждения центр может доминировать в деятельности ЦНС (принцип доминанты А.А. Ухтомского). В доминантном центре нервной системы нейроны характеризуются
длительной повышенной возбудимостью, возрастает эффективность временной и пространственной суммации возбуждений. Доминирующий центр, подавляя деятельность других центров, может стать главенствующим в формировании целенаправленного поведенческого акта животных и человека. Примером
доминанты служит классический опыт А.А. Ухтомского, в котором раздражение сенсомоторной коры на фоне механического раздувания прямой кишки вызывало реакцию дефекации вместо сокращения лапы животного.
Особенности распространения возбуждения в ЦНС в основном определяются свойствами нервных центров:
1. Одностороннее проведение возбуждения. В ЦНС возбуждение может распространяться только в одном направлении: от рецепторного нейрона через вставочный к эфферентному нейрону,
что обусловлено наличием синапсов.
2. Более медленное проведение возбуждения по сравнению с нервными волокнами. Промежуток времени от момента нанесения раздражения на рецептор до ответной реакции исполнительного органа называется временем рефлекса. Большая его часть тратится на проведение возбуждения в нервных центрах, где возбуждение проходит через синапсы. На выделение и диффузию медиатора в синапсе требуется промежуток времени в 1,5-2 мс (синоптическая задержка). Чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса.
3. Суммация возбуждений (или торможения). Нервные центры могут суммировать афферентные импульсы, что проявляется в усилении рефлекса при увеличении частоты раздражений или числа раздражаемых рецепторов. Различают два вида суммации: временная суммация — если импульсы приходят к нейрону по одному и тому же пути через один синапс с коротким интервалом, то происходит суммирование ВПСП на постсинаптической мембране и она деполяризуется до уровня, достаточного для генерации ПД; пространственная суммация связана с суммированием ВПСП, возникающих одновременно в разных синапсах одного нейрона. Оба вида суммации происходят в области аксонного холмика, где и генерируется ПД.
4. Конвергенция. В нервном центре несколько клеток могут передавать импульсы к одному нейрону, т. е. возбуждения конвергируют на нем. Конвергенция может быть результатом прихода возбуждающих или тормозных входных сигналов от различных источников. Так, моторные нейроны спинного мозга могут получать импульсы: от периферических нервных волокон, входящих в спинной мозг; волокон, соединяющих сегменты спинного мозга; кортикоспинальных волокон от коры мозга; тормозных путей от
ретикулярной формации. В результате конвергенции происходит суммация импульсов от этих источников и возникает ответ, являющийся суммарным эффектом разнородной информации.
5. Дивергенция и иррадиация. Возбуждение даже единственного нервного волокна, по которому импульсы поступают в нервный центр, может послужить причиной возбуждения множества выходящих из центра нервных волокон. Морфологическим субстратом широкого распространения импульсов (иррадиации) возбуждения является ветвление аксонов и наличие большого числа вставочных нейронов в пределах центра.
6. Облегчение и окклюзия. На нейронах нервных центров оканчиваются не только волокна их собственных афферентных
входов. Каждый из них получает веточки от афферентов соседне-
го центра, что может обусловливать развитие окклюзии (закупор-
ки) или, наоборот, облегчения.
Феномен окклюзии состоит в том, что количество возбужден-
ных нейронов при одновременном раздражении афферентных входов обоих нервных центров оказывается меньше, чем арифметическая сумма возбужденных нейронов при раздельном раздражении каждого афферентного входа в отдельности. Явление окклюзии приводит к снижению силы суммарной ответной реакции. Феномен центрального облегчения характеризуется противоположным эффектом. На облегчении основано проторение пути — распространение возбуждения не по той цепи нейронов, по
которой информация не поступала ранее, а через нейроны, уже облегченные первым раздражением.
7. Трансформация ритма возбуждений. Если сопоставить частоту импульсов в задних (чувствительных) и передних (двигательных) корешках спинного мозга при раздражении рецепторов, то обычно их ритм не совпадает. Центры способны как снижать, так и повышать ритмы возбуждений, поступающих от рецепторов.
8. Рефлекторное последействие. Продолжительность рефлекса всегда больше, чем время раздражения, так как возбуждение в нервных центрах сохраняется в течение некоторого времени после прекращения действия раздражителя. Это объясняется тем, что вставочные нейроны в центре образуют замкнутые цепи («нейрон-
9. Высокая чувствительность к недостатку кислорода.
Уменьшение доставки к клеткам мозга кислорода быстро ведет к тяжелым расстройствам деятельности ЦНС и гибели нейронов. Кратковременное нарушение кровоснабжения мозга вследствие временного спазма его сосудов или падения давления крови приводит к потере сознания — обмороку. Своевременно принятые
меры по восстановлению кровоснабжения мозга (нашатырный спирт, кофеин, горизонтальное положение тела и др.) выводят больного из обморока.
10. Высокая чувствительность к химическим веществам объясняется большим числом синапсов. На одном нейроне могут располагаться синапсы, обладающие чувствительностью к различным химическим веществам. Подбирая фармакологические препараты, которые избирательно блокируют одни синапсы, оставляя другие в рабочем состоянии, можно корректировать реакции организма.
11. Низкая функциональная подвижность (лабильность) и высокая утомляемость. Нервные центры, как и синапсы, обладают низкой функциональной подвижностью и быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми и имеют высокую лабильность.
12. Посттетаническая потенциация — явление усиления рефлекторного ответа после длительного ритмического раздражения нервного центра. Это связано с сохранением ВПСП на нейронах центра в течение некоторого времени, что облегчает проведение последующих возбуждений через синапсы.
13. Тонус нервных центров. Регистрация биоэлектрической активности ЦНС даже при отсутствии раздражений показывает, что многие нервные центры (составляющие их нейроны) постоянно генерируют импульсы. Эта импульсация поступает к рабочим органам и свидетельствует о существовании некоторого постоянного тонического возбуждения нервных центров.
14. Пластичность. Нервные центры обладают способностью изменять собственное функциональное назначение и расширять свои функциональные возможности, т.е. существенно модифицировать картину осуществляемых рефлекторных реакций. Пластичность нервных центров тесно связана с изменением эффек-
тивности или направленности связей между нейронами.
Свойства нервных волокон. Все нервные волокна характеризуются общими, присущими всем возбудимым тканям, свойствами — порогом возбуждения, лабильностью, циклическими изменениями возбудимости,
раздражения, так как неповрежденное нервное волокно в любом из своих участков на всем протяжении имеет одинаковые кабельные свойства.
• В норме возбуждение всегда распространяется ортодромно (прямо) — от тела нервной клетки по аксону вплоть до его концевых разветвлений.
• В эксперименте — при искусственной стимуляции участка нервного волокна — возбуждение может направиться антидромно — в направлении, противоположном естественному.
• Нервные волокна практически неутомляемы, так как проведение возбуждения связано только с их электрическими свойствами и не затрагивает сложных нейрохимических процессов.
• Скорость проведения в различных типах нервных волокон различна.
• По нервным волокнам передаются серии импульсов, которые имеют разные частоты и распределение во времени (рисунок, паттерн); доказано, что в паттерне закодирована информация о параметрах
раздражения, вызвавшего возбуждение.
• Из всех возбудимых образований нервные волокна обладают самой высокой функциональной лабильностью, т.е. проводят очень высокие частоты импульсации без трансформации ритма.
• В том случае, если частота импульсации превышает функциональную лабильность нервного волокна, возникает явление пессимума.
промежутки. Сравнивая время прохождения тока по оголенному проводнику и по унизанному бусами, он установил, что в последнем случае скорость проведения намного выше, чем в первом.
Безмиелиновые волокна на всем протяжении имеют одинаковую электропроводность и сопротивление. Вследствие деполяризации участка мембраны возникающий в нем локальный (местный) ток распространяется только на рядом расположенный невозбужденный. Волна деполяризации
идет последовательно, не имея возможности миновать ни один из невозбужденных участков волокна (рис. 5.2, Б).
Миелиновые волокна, как уже упоминалось, имеют изолирующий слой, резко уменьшающий емкость мембраны нервного волокна и практически полностью предотвращающий утечку тока из него. Перехваты узла шириной около 0,5 мкм, лишенные миелина, в отличие от миелиновых участков, имеют очень низкое сопротивление и поэтому являются центрами электрической активности. Практически все натриевые каналы сосредоточены в области перехватов — до нескольких тысяч на 1 мкм2, тогда как в
миелиновых участках их вообще нет. Невозбужденный участок волокна в области перехвата
электроположителен по отношению к аксоплазме, а возбужденный — электроотрицателен.
Итак, миелиновые волокна имеют очевидные преимущества:
исходного градиента 10:1 тратится значительно меньше энергии, чем
для реполяризации безмиелинового волокна;
• быстро, точно и дифференцированно проводят различные виды
чувствительности, обеспечивая максимально быстрые, адекватные реакции.
В процессе эволюции высших организмов скачок в развитии нервной
системы был, по-видимому, связан с началом миелинизации нервных
волокон. В онтогенезе, особенно у человека, отмечена корреляция между ми-
елинизацией некоторых проводящих путей и усложнением рефлекторного
и целостного приспособительного поведения.
Последнее изменение этой страницы: 2016-04-07; Нарушение авторского права страницы
Нервный центр — это совокупность связанных между собой нейронов, совместно выполняющих определённую функцию путём преобразования входящего возбуждения в выходящее с изменёнными характеристиками.
Анатомическое понятие "нервный центр" - это совокупность нейронов, располагающихся в строго определенных отделах центральной нервной системы и осуществляющих один рефлекс. Например: центр коленного рефлекса - в передних рогах 2-4 поясничных сегментов спинного мозга; центр глотания - на уровне продолговатого мозга: 5, 7, 9 пары черепно-мозговых нервов.
Здесь используется морфологический подход, т.е. нервные центры определяются по строению. Нервные клетки, образующие такой нервный центр, соединены в локальные компактные структуры: нервные узлы (ганглии) в периферической нервной системе или мозговые ядра в центральной нервной системе.
Физиологическое понятие "нервный центр" - это совокупность нейронов, расположенных на различных уровнях центральной нервной системы и регулирующих сложный рефлекторный процесс. Например: центр глотания входит в состав пищевого центра.
Такое определение созвучно понятию "функциональной системы", которое было предложено П.К. Анохиным. Но если функциональная система подразумевает временное объединение нейронов, то нервный центр - это обычно устойчивое образование. По П.К. Анохину различные анатомические нервные центры при необходимости могут на время объединяться в функциональную систему для получения определенного полезного результата.
Основные свойства нервных центров
Нейроны ЦНС для осуществления сложных и многообразных функций объединяются в нервные центры. Нервный центр —это совокупность нейронов, принимающих участие в осуществлении конкретного рефлекса (мигания, глотания, кашля и т.д.). В целом организме при формировании сложных адаптивных процессов происходит функциональное объединение нейронов, расположенных на различных уровнях ЦНС. Такое объединение (нервный центр в широком смысле слова) позволяет осуществлять наиболее адекватное для конкретных условий осуществления рефлекторной деятельности.
Нервные центры обладают рядом функциональных свойств и особенностей. К основным свойствам нервных центров относятся:
1) одностороннее проведение возбуждения —от рецептора к рабочему органу, обусловливается свойством синапсов односторонне проводить возбуждение.
2) Возбуждение в нервных центрах проводится медленнее, чем по нервному волокну. Это обусловлено замедленным проведением возбуждения через синапс (синаптическая задержка).
3) В нервных центрах происходит суммация возбуждений. Суммация может быть временная или последовательная, если импульсы возбуждения приходят к нейрону по одному и тему же пути через один синапс с интервалом меньше, чем время полной реполяризации постсинаптической мембраны и пространственная или одновременная суммация наблюдается в том случае, когда импульсы возбуждения поступают к нейрону одновременно через разные синапсы
4) Трансформация ритма возбуждения — изменение количества импульсов, выходящих из нервного центра, по сравнению с числом импульсов, приходящих к нему. Трансформация может быть в виде понижения, когда частота, поступающих импульсов превышает лабильность центральных синапсов или в виде повышения при однократном (но сильном) или редких импульсах, но высокой возбудимости центров.
5) Рефлекторное последействие — реакция заканчивается позже прекращения действия раздражителя. Это явление обусловлено либо длительной следовой деполяризацией мембраны нейрона, на фоне которой могут возникать несколько потенциалов действия, обеспечивающих кратковременное рефлекторное последействие, либо в результате циркуляции (реверберации) возбуждения в нейронной сети.. Возбуждение, попадая в такую сеть, может длительное время циркулировать в ней, обеспечивая длительное рефлекторное последействие. Возбуждение в такой цепочке может циркулировать до тех пор, пока какое-либо внешнее воздействие затормозит этот процесс или в ней наступит утомление.
6) Нервные центры обладают высокой чувствительностью к недостатку кислорода и к химическим веществам.
7) Нервные центры, как и синапсы, обладают быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми. Они обладают низкой лабильностью.
8) В нервных центрах легко возникает процесс торможения.
9) Нервные центры обладают пластичностью — способностью изменять собственное функциональное назначение, частично восстанавливать утраченные функции.
Процессы возбуждения и торможения тесно связаны между собой, протекают одновременно и являются различными проявлениями единого процесса. Очаги возбуждения и торможения подвижны, охватывают большие или меньшие области нейронных популяций и могут быть более или менее выраженными. Возбуждение непременно сменяется торможением, и наоборот, т. е. между торможением и возбуждением существуют индукционные отношения.
Торможение лежит в основе координации движений, обеспечивает защиту центральных нейронов от перевозбуждения. Торможение в ЦНС может возникать при одновременном поступлении в спинной мозг нервных импульсов различной силы с нескольких раздражителей. Более сильное раздражение тормозит рефлексы, которые должны были наступать в ответ на более слабые.
Читайте также:
