
Нервные клетки произошли от секреторных клеток факты
Факт №1: Гибель нервных клеток в нас изначально запрограммирована.
Факт №2: Наиболее интенсивно нейроны гибнут у детей
70% нейронов, заложенных еще при эмбриогенезе, гибнут ещё до рождения ребёнка. И это нормально – именно в детском возрасте способность к обучению должна быть максимальна, а потому и мозг должен обладать самыми значительными резервами. Резервами, которые прямо в ходе обучения апробируются и сокращаются по мере возможности, чтобы снизить нагрузку на весь организм. Иными словами, можно сказать, что избыточное количество клеток нервной системы – необходимое условие для обучения и то, что обеспечивает многообразие возможных вариантов развития человека – то есть нашу индивидуальность.
Смысл пластичности в том, что функции погибших нервных клеток берут на себя их оставшиеся в живых „коллеги“, которые увеличиваются в размерах и формируют новые связи, компенсируя утраченные функции. Одна живая нервная клетка может заменить девять погибших.
В зрелом возрасте процесс гибели клеток продолжается, но уже не так сильно. Однако, если не нагружать мозг новой информацией, то он будет оттачивать старые навыки, сокращая количество нервных клеток, необходимых для их реализации. Клеток будет становиться меньше, а связей у них с другими клетками – больше. И это, опять-таки, совершенно нормальный процесс.
У пожилого человека нейронов в мозгу существенно меньше, чем у младенца или молодого человека. Однако соображать он может намного быстрее, не говоря уж о том, что знает существенно больше. Всё это – благодаря отлично простроенной в процессе обучения архитектуре связей между нейронами.

А вот в старости, если нет обучения, то мозг человека, как и весь организм, начинает программу свёртывания – то есть старения, приводящего к смерти. И чем меньше показатели востребованности у разных систем организма, то есть чем меньше физическая и интеллектуальная нагрузка, чем меньше человек двигается и общается с другими людьми – тем процесс быстрее. Вот почему требуется постоянно осваивать новую информацию.
Факт №3: Нервные клетки восстанавливаются – причем генерируются в трех местах человеческого организма
Новые нервные клетки не появляются в ходе деления, как это происходит в других органах и тканях организма, а образуются в ходе нейрогенеза, который наиболее активен во время внутриутробного развития. Нейрогенез начинается с деления предшественников нейронов - нейронных стволовых клеток, которые затем мигрируют, дифференцируются и образуют полностью функционирующий нейрон.
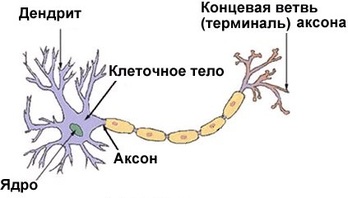
Нейрон — особенная клетка, у неё имеются отростки: длинные называются аксонами, а короткие разветвлённые —дендритами. Нейроны генерируют нервные импульсы, передавая их соседним нервным клеткам. Средний диаметр тела нейрона составляет около 0,01 мм, а общее количество нейронов в головном мозге достигает 100 миллиардов. Если тела всех нейронов головного мозга выстроить в одну линию, то её длина составит 1000 километров. Нейроны отличаются друг от друга по размеру, разветвлённости дендритов и длине аксонов. Самые длинные аксоны достигают метра. Это — аксоны гигантских пирамидных клеток коры больших полушарий. Они тянутся к нейронам нижних отделов спинного мозга, контролирующим двигательную активность мышц туловища и конечностей.
Впервые сообщение об образовании новых нервных клеток во взрослом организме млекопитающих появилось ещё в 1962 году. Но тогда результаты работы Джозефа Олтмана, опубликованные в журнале Science, не были восприняты всерьёз, и признание нейрогенеза отложилось почти на двадцать лет. С тех пор неоспоримые доказательства существования этого процесса во взрослом организме были получены для певчих птиц, грызунов, амфибий и некоторых других животных. И только в 1998 году нейробиологам во главе с Питером Эрикссоном и Фредом Гейгом удалось продемонстрировать образование новых нейронов в гиппокампе человека и доказать существование нейрогенеза в головном мозге взрослых людей. Сейчас исследование нейрогенеза является одним из самых приоритетных направлений в нейробиологии. Учёные и медики видят в нём большой потенциал для лечения дегенеративных заболеваний нервной системы, таких как болезнь Альцгеймера или болезнь Паркинсона.
Факт №4: Стволовые нейрональные клетки мигрируют в организме
Так же, как у птиц и низших позвоночных, у млекопитающих нейрональные стволовые клетки располагаются поблизости от боковых желудочков мозга. Их перерождение в нейроны идёт очень интенсивно. Так, у взрослых крыс за месяц из стволовых клеток образуется около 250 000 нейронов, замещая 3% всех нейронов гиппокампа. Продолжительность жизни таких нейронов очень высока — до 112 дней. Более того, стволовые нейрональные клетки мигрируют! И в среднем преодолевают путь длиной около 2 см. И если они попадают в обонятельную луковицу, то превращаются там в нейроны.
Также стволовые клетки можно извлечь из мозга и пересадить в другой участок нервной системы, где они превратятся в нейроны. Недавно проведённые исследования показали, что новые нейроны во взрослом мозге могут образовываться не только из нейрональных стволовых клеток, но из стволовых клеток крови. Однако последние не превращаются в нейроны, а сливаются с ними, образуя двуядерные клетки. Затем „старое“ ядро нейрона разрушается, а его замещает „новое“ ядро стволовой клетки крови.
Факт №5: Нервные клетки неспособны гибнуть от стресса, они просто тормозят свою деятельность и отдыхают
При стрессе клетки гибнут не от избыточного напряжения. Нейроны вообще не способны погибнуть от перегрузки - они просто тормозят свою деятельность и отдыхают. Нейроны гибнут от возникающего недостатка питательных веществ, особенно витаминов и из-за нарушения кровоснабжения тканей, приводящих к интоксикации продуктами жизнедеятельности и гипоксии, употреблении разнообразных лекарств, крепкого кофе и чая, наркотиков, табака и алкоголя, значительных физических нагрузках, и инфекционных болезнях. А сберечь их очень просто. Достаточно постоянно учиться чему-то новому, а также развивать уверенность в себе и крепкие эмоциональные связи с близкими людьми.
Глава II. Возникновение нервных клеток и мозга
Причиной возникновения нервной системы стала низкая скорость получения информации о внешнем и внутреннем мире организма с донервной организацией. Его ткани состояли из клеток со сходной химической, электромагнитной и механической чувствительностью. Такой организм не мог оперативно получить дифференцированные сведения из внешнего мира и от собственного тела (Lipps, Signor, 1992). Решение проблемы без участия нервной системы было найдено растениями и одноклеточными животными, что разбиралось в главе I. Этот путь оказался биологически очень успешным и привёл к появлению процветающих групп организмов. Можно сказать, что растения и одноклеточные животные выиграли ничуть не меньше, чем животные с развитой нервной системой. Тем не менее нервная система возникла. Она давала одно, но огромное преимущество, которого невозможно достичь при помощи универсальных свойств неспециализированных клеток. Это увеличение скорости реакции организма на изменяющиеся внутренние и внешние условия. Как только нервная система позволила животным быстро адаптироваться к изменяющимся условиям, активно разыскивать пищу и половых партнёров, наступило время динамичной эволюции животного мира. Скорость решала почти все проблемы. Конкуренция за источники питания привела к появлению бесконечного разнообразия растительноядных организмов, но они сами были питательнее растений. Ещё более быстрые и эффективные хищники должны были обладать более совершенной нервной системой, чем травоядные. Это касалось как позвоночных, так и беспозвоночных животных. Кто обладал эффективной нервной системой, тот быстрее поднимался на вершину пищевой цепочки, питаясь не грубой растительной пищей, а калорийными родственниками. В самом общем виде эта ситуация сохранилась и в социальных структурах высших приматов.
Преимущества и достоинства нервной системы вполне понятны. Причины её появления вполне логичны и не вызывают сомнения. Совершенно иная ситуация с вопросом о том, как это произошло. Нет ясного представления о механизмах появления первых нервных клеток. Почему они дифференцировались от других тканей и от чего возникли их уникальные свойства? Почему мозг позвоночных имеет 5 гомологичных отделов? Почему так различно поведение животных из одной систематической группы и так сходно у отдалённых видов? Как возникли и строились ассоциативные центры на различных структурных принципах? Эти вопросы пока остаются без ответа. Не менее принципиальны проблемы становления мозга при появлении первых хордовых, выходе позвоночных на сушу, появлении птиц и млекопитающих. Эти глобальные эволюционные события требовали перестройки нервной системы и органов чувств. Как это могло произойти с такой консервативной и инертной системой, как мозг? Его же нельзя частично выключить из работы или рудиментировать, как задние конечности у китообразных. В нервной системе происходили процессы, совершенно не похожие на эволюцию конечностей, позвоночника, сердца, лёгких или хвоста. Эволюционные преобразования в нервной системе не укладываются полностью в рамки традиционной морфологической методологии. В связи с этим мне пришлось несколько адаптировать приёмы эволюционной морфологии к реконструкции событий, произошедших в непроверяемом прошлом.
Происхождение нервных клеток
Происхождение нервных клеток в процессе эволюции животных довольно спорно. Существует несколько основных точек зрения на эту проблему. Наибольшее распространение получила точка зрения братьев Гертвигов (Hertwig, 1878). Их гипотеза состоит в том, что нервные и мышечные клетки происходят из эктодермального эпителия. Они возникают независимо друг от друга и вступают во взаимодействия уже вторично (рис. II-1, а-в). В соответствии с гипотезой Гертвигов источником всех нервных клеток является первичная чувствительная клетка, возникшая из эктодермы и получившая возможность воспринимать раздражения, генерировать и проводить возбуждение. Специализированные отростки этих клеток вступают в связь с независимо возникшими мышечными клетками. По мнению авторов этой гипотезы, мышечные волокна возникли из клеток, сходных с эпителиально-мышечными клетками кишечнополостных. Такой простейший нервно-мышечный комплекс развивается по пути дифференциации нервного аппарата, что приводит к возникновению вставочных нейронов или ганглиев (см. рис. II-1, в). Нервная система подразделяется на чувствительные клетки, лежащие в эктодерме, и вставочные нейроны, обладающие интегративной функцией. Вставочные нейроны являются предвестниками централизации обработки сенсорной информации и возникновения мозга. На выходе расположены эффекторные нейроны, иннервирующие мышцы и железы.

а-в — гипотеза братьев Гертвигов; г-е — гипотеза Клейненберга-Заварзина. По гипотезе Гертвигов нервные и мышечные клетки происходят независимо из эктодермы, а затем объединяются в нервномышечный комплекс. По представлениям Клейненберга-Заварзина, из эктодермы возникает клетка одновременно с сенсорными и моторными функциями. Затем она дифференцируется на нервную и мышечную системы. Красные — мышечные клетки или волокна, синие и фиолетовые — чувствительные нервные клетки.
Считается, что обе системы возникли одновременно в виде образования, которое не имело дифференцированных мышечных и нервных компонентов. В соответствии с этой гипотезой первичными были клетки, выделившиеся из эктодермы и обладавшие как нервными, так и мышечными функциями (см. рис. II-1, г, д). Часть клетки была способна к чувствительности, а другая — к сокращению. В гипотезе подчёркивается первичность морфофункциональной связи между нервными и мышечными клетками. В дальнейшем, по мнению авторов, произошла дифференциация чувствительно-сократимых клеток на чисто сенсорные и моторные, а общность их происхождения стала основой нервно-мышечных взаимодействий (см. рис. II-1, е). Обе теории роднит то, что они ничего не объясняют. Непонятно, зачем вообще понадобились нервные, да и мышечные клетки, почему они возникли одновременно и откуда взялись отростки нервных клеток вместе с особыми свойствами рецептировать, генерировать и проводить сигналы. В дискуссии о природе нервномышечных взаимодействий как-то сам собой исчез биологический смысл происхождения нервных клеток. Этот вопрос можно рассмотреть и с других позиций.

У гребневиков имеется уникальный тип клеток, которые получили название коллобласты. Они располагаются на щупальцах и выделяют липкий секрет, с помощью которого гребневик ловит свою добычу. В августе 2018 года в журнале Molecular Biology and Evolution появилось сообщение, что в ходе развития коллобласты образуются от тех же клеток-предшественников, что и нейроны! Более того, выяснилось, в ряде давно забытых работ ещё четверть века назад была показана общность происхождения нейронов и стрекательных клеток у медуз, а также секреторных клеток и нейронов у плодовой мушки-дрозофилы. Об этом свидетельствует и доклад на только что закончившейся конференции Society for Integrative and Comparative Biology (SICB), который и стал информационным поводом заметки в Science.
Напрашивается идея, что общность происхождения секреторных и нервных клеток – это универсальная черта всех многоклеточных организмов, имеющих нервную систему, и в этом отношении гребневики вовсе не являются исключением. Вероятно, секреторные и нервные клетки, которые в ходе индивидуального развития берут начало от одних и тех же клеток-предшественников, имелись уже у последнего общего предка настоящих многоклеточных животных (то есть всех животных, за исключением губок и пластинчатых).
Впрочем, убеждённые сторонники уникальности нервной системы гребневиков и тут нашли что возразить: теоретически возможно, что нейроны возникали в ходе эволюции несколько раз, и каждый раз от стволовых клеток одной и той же популяции. Так что остаётся лишь надеяться, что дальнейшие исследования прольют свет на происхождение нейронов и помогут разгадать загадку необычной нервной системы гребневиков.
Текст: Елизавета Минина
Читайте материалы нашего сайта в Facebook, ВКонтакте, Яндекс-Дзен и канале в Telegram, а также следите за новыми картинками дня в Instagram.

Российские морские биологи изучили строение нервной системы необычной круглоротой мшанки, построили ее компьютерную модель и сравнили с другими мшанками. Исследование показало, что вид Crisia eburnea…

Эта странная картинка показывает одну анатомическую особенность беспозвоночного, которая некогда изменила нейронауки. Слева вы видите сечение гигантского аксона нейрона кальмара. Справа, для сравнения, — нервное волокно…

Посмотреть, что происходит в нервной системе животного – беспозвоночного или позвоночного – в момент движения по понятным причинам очень непросто. Чтобы понять, какие цепочки нейронов…

Даже у плодовой мушки дрозофилы, не говоря уж про нас с вами, мозг подразделен на множество функциональных участков, в которых находятся различные виды нейронов и…
Один из лидеров нейронаук в нашей стране – Филипп Хайтович, профессор Сколковского института науки и технологий (Сколтех) и руководитель группы сравнительной биологии в Институте Вычислительной…
Секреторные нейроны
Особенности гистофизиологии нервных клеток не исчерпываются тем, что о них было сказано выше. Установлена способность нервных клеток секретировать биологически активные вещества, сходные с гормонами по механизму их выделения из клетки и действия на мишени. Этот феномен получил название нейросекреции. Идея о том, что нервные клетки наделены секреторной функцией, была впервые сформулирована Э. Шаррером в 1928 г. на основе морфологических исследований нейронов гипоталамуса у рыб. Позднее он и его коллеги описали аналогичный феномен в нервных клетках гипоталамуса млекопитающих. Было показано, что морфологический вид нейросекреторных клеток зависит от состояния гидратации организма, и установлено, что экстракты, выделенные из гипоталамуса, обладают антидиуретической активностью. Эти данные позволили предположить, что антидиуретический гормон, выделяемый в кровь из задней главной части нейрогипофиза, в действительности синтезируется не клетками нейрогипофиза, а нейронами гипоталамуса. Фактами, которые подтвердили правильность этого предположения, были, с одной стороны, наблюдения о существовании в нервных клетках тока аксоплазмы, то есть транспорта составных частей цитоплазмы от тела клетки к окончанию его аксона, с другой – демонстрации того, что нейросекрет в гипоталамо-гипофизарной системе накапливается выше места перерезки или перевязки стебля гипофиза.
Существуют различные модели структурной организации нейросекреторной системы. Классической моделью, с которой началось развитие наших знаний о нейросекреции, может служить гипоталамо-нейрогипофизарная система, которая будет подробно освещена в разделе частной гистологии. Здесь же уместно лишь упомянуть, что нейросекрет (вазопрессин и окситоцин) вырабатываются в клеточных телах нейронов супраоптических и паравентрикулярных ядер гипоталамуса, транспортируется далее по аксонам последних в заднюю, главную часть нейрогипофиза, где депонируется в нервных терминалях и выделяется из них в кровь. Подобно истинным гормонам, биологически активные пептиды, заключенные в окрашенном нейросекрете, регулируют функции органов, удаленных от места секреции этих пептидов.


Позднее были установлены альтернативные модели нейросекреторной активности. Их примерами могут быть выделение нейросекреторных пептидов в ликвор желудочков мозга или в синаптическую щель. Последняя из упомянутых возможностей привлекает к себе наибольшее внимание, так как в этом случае нейросекреторные пептиды рассматриваются в качестве агентов, влияющих на транссинаптическую передачу, то есть нейромедиаторов или нейромодуляторов. Допускается, что отличия между ними не являются абсолютными, а скорее сводятся к тому, что нейромодуляторы обладают более длительным латентным периодом и более длительным эффектом действия. Общепринято, что их основной функцией является изменение чувствительности мишени к действию нейромедиаторов. Поскольку передача в нервной системе осуществляется преимущественно через посредство химических мессенджеров, нейросекрецию следует рассматривать как фундаментальное свойство всех нейронов. Окончательный маршрут нейрогормона в аксонах и места его биологического действия определяются топографией межнейронных взаимодействий.
Внутриклеточные механизмы синтеза нейрогормонов сходны с таковыми в типичных эндокринных железистых клетках, например, в клетках панкреатических островков. Пептидные гормоны синтезируются в канальцах ГЭС, а упаковка в секреторные гранулы происходит на мембранах комплекса Гольджи. Выделение секреторного продукта совершается путем обратного пиноцитоза в ответ на распространение потенциала действия.
Нейросекреторные клетки, при всем их сходстве с железистыми, в отношении генетической программы синтеза пептидных гормонов и внутриклеточного аппарата его реализации сохраняют структурные и функциональные признаки нейронов. Им присущи типичные для нейронов электрофизиологические характеристики и свойственные нейронам органеллы. Сходным образом они подвергаются воздействию со стороны других нейронов через синаптический аппарат и отвечают на действие нейротрансмиттеров. Единственное отличие заключается в том, что их ответы наряду с деполяризацией электровозбудимых мембран и освобождением нейромедиатора включают в себя также выделение пептидных нейрогормонов, которые вызывают соответствующие изменения со стороны эндокринных или гомеостатических функций.
Согласно этой концепции, эмбриональные клетки, из которых детерминируются нейроны, изначально наделены способностью синтезировать не только нейромедиаторы, но и гормоны. В зависимости от того, какие локусы их генов экспрессируются, они дифференцируются либо в классические нейроны, либо в нейросекреторные клетки, или в эндокринные клетки. Отсюда Пирс рассматривает эндокринологию как частный случай нейроэндокринологии, а в нервной системе выделяет три компонента: соматическую нервную систему, вегетативную и эндокринную. Исходя из этой концепции, можно понять, почему в значительном числе нейронов мозга обнаруживаются пептидные гормоны, секрецию которых считали прежде исключительной прерогативой эндокринных желез.
Взгляды Пирса получили широкое распространение. Какова же роль нейропептидов мозга, если подходить к их оценке с позиций нейрофизиологов? Уже упоминалось мнение о том, что они могут выполнять нейромедиаторную функцию. В переживающих срезах мозга нейропептиды подобно нейромедиаторам выделяются в ответ на деполяризацию мембраны. Этот процесс является кальцийзависимым. При центрифугировании в градиенте плотности нейропептиды так же, как и нейромедиаторы, обнаруживаются во фракции синаптосом. Они способны изменять активность нейронов. Наконец, для многих из них выявлены рецепторы на постсинаптических мембранах. Для многих нейропептидов установлены корреляты нейрофизиологической активности, что позволяет предполагать их важную роль в механизмах мотиваций, обучения и памяти.
Хотя феномен нейросекреции стал уже привычным понятием, все еще не угасают споры среди нейрофизиологов в его трактовке. Это относится, в частности, к попыткам рассматривать роль нейропептидов в качестве нейромедиаторов.
APUD-концепция позволяет трактовать этот постулат более компромиссно: один нейрон – несколько медиаторов. Вполне возможно, что новые факты вносят коррекцию в правило Дейла, однако нельзя исключить и того, что в каждом нейроне один из медиаторов является профилирующим, и по содержанию которого можно судить о принадлежности нейрона к определенному типу, тогда как нейропептиды функционируют в роли модуляторов транссинаптической передачи. Хотя окончательное решение вопроса еще ждет своих исследователей, на сегодня очевидно одно: появление и творческое преломление в нейробиологии APUD-концепции существенно приблизило к пониманию не только обширного потока новой информации, но и некоторых белых пятен в наших прежних знаниях. В этой связи интересен такой пример. Агентом, влияющим на потоотделение, является ацетилхолин, выделяемый окончаниями парасимпатических нейронов. Этот агент – атропинзависимый. Между тем сопутствующая потоотделению вазодилятация является атропинрезистентной, что давно озадачивало физиологов. Исследования последних лет показали, что наряду с ацетилхолином в тех же нервных окончаниях содержится нейропептид VIP (вазоактивный интестинальный пептид), который является медиатором атропинрезистентной вазодилятации. Отсюда следовало, что в ответ на адекватный стимул из одного и того же нервного окончания выделяются два различающихся по природе, но синнергических по конечному результату медиатора, каждый из которых связывается со своим собственным рецептором.
Такого рода факты привлекают внимание не только нейрофизиологов, но и клиницистов. В этой связи заслуживает упоминания факт сосуществования в одних и тех же нейронах нейропептида холецистокинина и традиционного медиатора дофаминовых систем мозга. Установлено, что метаболиты холецистокинина (короткоживущего пептида) тормозят выделение дофамина из нервных терминалей, откуда допускается, что в дофаминовых нейронах холецистокинин вовлекается в механизм тормозящей обратной связи, который в норме модулирует выделение медиатора. При отсутствии холецистокинина или резком снижении его содержании избыточно активируется дофаминовая система, что характерно для параноидной формы шизофрении, протекающей с галлюцинациями и нарушением высшей нервной деятельности. Принимая во внимание, что некоторые дофаминовые системы мозга, например мезолимбическая, тесно связаны с процессами высшей нервной деятельности, высказывается предположение, что нарушение естественного баланса между содержанием в нервных клетках холецистокинина (или его метаболитов) и дофамина может служить патогенетическим фактором в развитии шизофрении.
Наконец уместно упомянуть о том, что холецистокинин и ВИП являются мощными стимуляторами активности нейронов коры больших полушарий мозга, холецистокинин к тому же известен как пептид, вызывающий чувство насыщения; ангиотензин-II вызывает чувство жажды; люлиберин влияет на половое поведение. Болевые ощущения связывают с рядом пептидов: субстанцией P, нейротензином, брадикинином, тогда как обезболивающий эффект связывают с опиоидными пептидами семейства энкефалинов и эндорфинов. Дельта-пептид, то есть пептид, индуцирующий дельта-фазу сна, называют пептидом сна, а вазопрессин и его фрагменты играют важную роль в механизмах обучения и памяти. Перечисленные нейроактивные соединения далеко не исчерпывают известных к настоящему времени нейропептидов мозга. Сходным образом вышеизложенные сведения о секреторной функции нейронов не исчерпывают прогрессивно накапливаемых знаний об этой замечательной и ранее неизвестной особенности гистофизиологии нервных клеток.
Функциональные возможности организма обеспечивают взаимодействие 2-х систем: нервной и гуморальной. Возможности таких взаимоотношений этих 2-х систем могут осуществляться благодаря наличию в межуточном мозге нейросекреторных клеток. Последние обладают способностью выполнять функции нервных клеток и секреторных клеток.
Будучи нервными клетками, они воспринимают сигнал, обрабатывают его и передают другим клеточным структурам. Однако, в отличие от нервных клеток, нейросекреторные клетки способны синтезировать и секретировать различные гормоны – нейрогормоны; они являются веществами белковой природы, и работа нейросекреторных клеток осуществляется циклично. Поленов выделил в функции нейросекреторных клеток 3 фазы:
Эти фазы меняют друг друга, после последней фазы, гранулы нейрогормонов выводятся в кровь и в ликвор (спинномозговую жидкость). Нейрогормоны регулируют функции эндокринных желез, которые, в свою очередь, выбрасывают гормоны в кровь и осуществляют регуляцию активности различных органов и систем.
Под действием тропных гормонов активируется синтез гормонов периферическими железами. Однако между гипоталамусом, гипофизом и периферическими железами существует не только прямая, но и обратная связь. К примеру: под действием тириотропных гормонов (ТГ), активируется щитовидная железа, которая синтезирует и секретирует в кровь тироксин. Уровень тироксина в крови анализируется специальными клетками гипоталамуса, которые, в свою очередь, превышают секрецию либиринов и статинов.
Нейроглия
В отличие от нервных клеток, глиальные клетки обладают большим разнообразием. Их количество в десятки раз превышает количество нервных клеток. В отличие от нервных клеток, глиальные способны делиться, их диаметр значительно меньше диаметра нервной клетки и составляет 1,5-4 микрона.
Долгое время считали, что функция глиоцитов несущественна, и они выполняют лишь опорную функцию в нервной системе. Благодаря современным методам исследования, установлено, что глиоциты выполняют ряд важных для нервной системы функций:
- опорная
- разграничительная
- трофическая
- секреторная
- защитная
Среди глиоцитов, по морфологической организации, выделяют ряд типов:
- эпендимоциты
- астроциты
Эпендимоциты образуют плотный слой клеток, элементов, выстилающих спинномозговой канал и желудочки мозга. В процессе онтогенезе, эпендимоциты образовывались из спонгиобластов. Эпендимоциты представляют собой слегка вытянутые клетки с ветвящимися отростками. Некоторые эпендимоциты выполняют секреторную функцию, выделяя биологически активные вещества в кровь и в желудочки мозга. Эпендимоциты образуют скопления на капиллярной цепи желудочков мозга; при введении в кровь красителя, он накапливается эпендимоцитах, это свидетельствует о том, что последние выполняют функцию гематоэнцефалического барьера.
Астроциты выполняют опорную функцию. Это огромное количество глиальных клеток, имеющих множество коротких отростков. Среди астроцитов выделяют 2 группы:
- плазматические клетки
- волокнистые астроциты
Олигодендроциты – крупные глиальные клетки, часто сконцентрированы вокруг нервной клетки и поэтому называются сатиллитными глиацитами. Их функция очень важна для трофики нервной клетки. При функциональных перенапряжениях нервной клетки, глиоциты способны прореферировать вещества поступающие путем пиноцитоза в нервную клетку. При функциональных нагрузках, вначале происходит истощение синтетического аппарата глиальных клеток, а затем нервных. При восстановлении (репарации), вначале восстанавливаются функции нейронов, а затем – глиальных клеток. Таким образом, глиоциты принимают участи1е в обеспечении функций нейронов. Глиальные клетки существенным образом способны влиять на трофику мозга, а также на функциональный статус нервной клетки.
Функционально морфологическая организация синаптических структур. Типы синапсов. Медиаторы и их функциональное значение
Передача сигнала от одной клетки к другой или от нервной клетки к эффекторной клетки осуществляется через синапсы. Синапс состоит из 3-х элементов: пресинаптическая мембрана (1); постсинаптическая мембрана (2); синаптическая щель (3).
Проходящий по аксону импульс достигает пресинаптической мембраны, что приводит к её деполяризации. При этом пресинаптическая мембрана открывает кальциевые каналы, кальций мигрирует через эти каналы и связывается с белком. У пресинаптической мембраны находятся визикулы (шарики, пузырьки) с медиатором. Комплекс белка и кальция (кальмодулин), приводит к слиянию везикул с пресинаптической мембраной и кванты медиатора экзоцитозом выбрасываются в синаптическую щель, достигает постсинаптической мембраны, на которой находятся рецепторы. Медиатор связывается с рецептором и возникает деполяризация постсинаптической мембраны. Так работает возбуждающий синапс, медиатором у которого может быть ацетилхолин.
Однако передача сигнала по нервному волокну осуществляется импульсами и к каждому последующему импульсу на постсинаптической мембране должен восстановиться мембранный потенциал. Это возможно благодаря тому, что в синаптической щели находится фермент, разрушающий медиатор, благодаря чему восстанавливается мембранный потенциал. Для каждого медиатора имеются специфические ферменты. Таким образом, мы описали работу возбуждающего синапса.
Однако кроме возбуждающих синапсов имеются тормозные, которые имеют специфические медиаторы (допомин, ГАМК и др.). В катехоламинергических синапсах, где медиатором является норадреналин, серонин, дофомин. После каждого импульса часть медиатора разрушается ферментом, а остальная – захватывается обратно через пресинаптическую мембрану.
В возбуждающих синапсах медиатор вызывает деполяризацию постсинаптической мембраны, а в тормозных – гиперполяризацию (т.е. увеличивается величина мембранного потенциала).
Синапсы бывают химические и электрические, выше мы рассматривали работы химических синапсов. В химических синапсах величина синаптической щели составляет от 100 до 40 ангстрем. В электрических синапсах эта величина составляет от 20 до 59 ангстрем. У человека в ЦНС находятся химические синапсы.
Рефлекс как основной факт нервной деятельности. Рефлекторная дуга и рефлекторное кольцо
Деятельность нервной системы осуществляется с помощью рефлекторных актов. Благодаря рефлексам возникает или исчезает, усиливается или ослабевают функции тех или иных органов. Путь, по которому осуществляется рефлекс, называется рефлекторной дугой.
Рефлекторная дуга состоит из ряда звений:
2. Афферентное звено
3. Чувствительные спинномозговой ганглии
4. Переключающие структуры мозга
5. Исполнительный орган
Спинальная рефлекторная дуга осуществляется при участии определённых отделов спинного мозга.
При раздражении рецепторов (1), сигнал поступает к чувствительным ганглиям (2), аксоны этих ганглиев формируют задние корешки спинного мозга, они переключаются на нейронах задних рогов спинного мозга (4). Затем через ряд вставочных нейронов (5) сигнал поступает к мотонейронам передних рогов спинного мозга (6), из передних рогов спинного мозга выходят передние корешки спинного мозга (7), которые иннервируют эффекторный орган (8).
Для коррекции выраженности рефлекторного фактора в организме имеется другая структура – морфологическая организация, которая называется рефлекторным кольцом.
Суть работы рефлекторного кольца заключается в том, что сигналы, поступающие через задние рога спинного мозга, переключаются не только на мотонейронах передних рогов, но и по восходящим путям спинного мозга попадает в стволовую структуру головного, подкорковые структуры и в кору. Здесь происходит анализ и синтез поступившей информации и далее она поступает по нисходящим структурам и мотонейронам передних рогов. Эти сигналы могут усилить работу нейронов, либо затормозить.
Для восприятия сигналов из внешней среды на теле человека существуют определённые структуры – рецепторы.
Для раздражителя различной природы (светового, звукового, теплового_ существуют специфические рецепторы, которые воспринимают адекватные раздражители только одной природы. Чаще всего для каждого раздражителя существуют определённые параметры восприятия. К примеру: человеческое ухо воспринимает сигналы частотой от 20 до 20 тыс. Гц.
Кроме того, рецепторы обладают способностью адаптироваться к воздействию внешних раздражителей. При действии сильных раздражителей, порог чувствительности повышается. Например: если человек вышел из тепла сразу в холод – то человеку вначале холодно, но затем рецепторы повышают свой порог и человеку становится тепло.
Также рецепторы кодируют силу действующего раздражителя и транспортируют его в определённую частоту электрических импульсов. Эти импульсы воспринимают соответствующие структуры головного мозга, и они формируют адекватную реакцию организма на раздражитель.
По расположению рецепторов в человеческом теле их делят на:
- экстерорецепторы (расположены на поверхности тела)
- интрорецепторы (направляют сигнал от внутренних органов)
- проприорецепторы (сигнализируют ЦНС о степени напряжения мышц).
В зависимости от того, какие отделы мозга принимают участие в осуществлении того или иного рефлекса, существуют следующие типы рефлексов:
- бульбарные (для их осуществления необходима целостность продолговатого мозга)
- кортикальные
по биологическому значению классифицируют:
- пищевые рефлексы
- половые рефлексы
- локомоторные рефлексы
Рефлекторные дуги бывают:
- моносинаптические. Имеют один синапс при переключении от мотонейронов задних рогов к мотонейронам передних рогов.
- полисимпатические. Имеют много синапсов и переключение осуществляется через ряд вставочных нейронов.
Читайте также:
