
Нервные сети это физиология
Нервные сети
Нейроны неррного центра за счет структурно-функциональных связей (ветвления отростков и установления множества синапсов между разными клетками) объединяются в нервные сети. Связи между нервными клетками являются генетически детерминированными.
Основные типы нервных сетей
Дивергентные сети с одним входом представляют собой нейронные ансамбли, в которых один нейрон образует выходные связи с большим количеством других клеток разных иерархических уровней и, главное, разных нервных центров. Максимально выраженная дивергенция связей разных нервных центров свидетельствует о том, что эти нервные сети не являются специфическими для реализации определенных рефлексов, а обеспечивают интеграцию разных рефлекторных актов и общее состояние активности многочисленных нейронов разных отделов мозга.
Поскольку в нервных сетях включено огромное число вставочных нейронов, они могут, передавая импульсацию, реализовывать как возбуждение, так и торможение.
Типы тормозных процессов в нейронных сетях
Реципрокное торможение (рис.3.9-1), т.е. процесс, основанный на том, что сигналы по одним и тем же афферентным путям обеспечивают возбуждение одной группы нейронов, а через вставочные тормозные клетки вызывают торможение другой группы нейронов. Этот тип торможения называют также сопряженным. Проявляется реципрокное торможение, например, на уровне мотонейронов спинного мозга, иннервируюших мышцы-антагонисты (сгибатели-разгибатели) конечностей. Передача возбуждения к мотонейронам мышц-сгибателей одновременно через вставочные тормозные нейроны тормозит мотонейроны мышц-разгибателей, благодаря чему только и возможно осуществление сгибательного рефлекса;
Возвратное торможение (рис.3.9-2), т.е. процесс торможения эфферентных нейронов сигналами, возвращающимися к ним по коллатералям аксонов через вставочные тормозные клетки.

Рис.3.9. Тормозные процессы в нервных сетях центра.
Возбуждение от одних нейронов поступает через ответвления на аксонах к вставочным тормозным нейронам, а аксоны этих клеток в свою очередь образуют тормозные синапсы на телах первых нейронов. Примером является торможение Реншоу в спинном мозге, где импульсы от двигательных нейронов посылаются по аксонам не только к скелетной мышце, но поступают через аксонные коллатерали к тормозным нейронам Реншоу, а от них назад к двигательным клеткам, приводя к их торможению. Таким образом, осуществляется короткая отрицательная обратная связь, не позволяющая возникнуть избыточному возбуждению мотонейронов спинного мозга.
1 — реципрокное торможение в центрах спинного мозга, управляющих движением нижних конечностей;
2 — возвратное торможение (Реншоу) в спинном мозге,
3 — латеральное торможение,
4 — окружающая тормозная зона
Латеральное торможение, т.е. процесс торможения группы нейронов, расположенной рядом с группой возбужденных клеток. Этот вид торможения распространен в сенсорных системах, где ответвле ния аксонов возбужденных нейронов образуют синапсы на вставочных тормозных нейронах, а аксоны последних оканчиваются тормозными синапсами на соседних, рядом расположенных клетках, получающих информацию от другого афферентного входа (рис.3.9-3);
Тормозная зона, т.е. процесс торможения нескольких групп нейронов, расположенных рядом с возбужденными клетками. Сигналы от возбужденных нейронов по коллатералям аксонов поступают к вставочным нейронам, а аксоны последних образуют тормозные синапсы на группах клеток, получающих информацию по другим афферентным входам (рис.3.9-4). Если эта группа заторможенных клеток располагается вокруг возбужденных нейронов, тормозную зону называют окружающей.
Нервная система человека и животных может быть представлена как система нейронных цепочек, передающих возбуждающие и тормозные сигналы (нервная сеть).
Существуют значительные количественные различия нервных сетей у разных видов позвоночных и беспозвоночных. Так, у человека нервная система включает 10 элементов, у примитивных беспозвоночных - около 10 нейронов (их нервная система называется малонеиронной). Однако в строении и функционировании всех нервных систем имеются общие черты
В нервной системе позвоночных и беспозвоночных можно выделить следующие категории нейронов
чувствительные (сенсорные) нейроны, формирующие чувствительные нервные окончания - рецепторы,
двигательные, возбуждающие (а у беспозвоночных также и тормозящие) мускулатуру нейроны, сюда же можно отнести нейроны, запускающие работу электрических органов (рыб) и желез, все эти нейроны называют эффекторными,
вставочные нейроны, передающие сигналы от сенсорных к эффекторным
Особую категорию составляют так называемые командные вставочные клетки, которые благодаря связям со множеством эффекторных нейронов запускают более или менее сложные поведенческие акты.
Все эти клетки наиболее подробно изучены в малонейронной нервной системе моллюсков (например, у аплизии) Там осуществлена идентификация многих крупных элементов, которые имеют стандартное положение в ганглиях и стандартные функции у всех особей. Предполагают, что такую же жесткую регламентацию структуры и свойств могут иметь и некоторые нейроны высших животных.
Во всех изученных нервных системах (сетях) обнаружены дивергенция путей, передающих те или иные сигналы, конвергенция путей, передающих информацию в высшие, а также в исполнительные центры, различные варианты тормозных связей между элементами нервных цепочек.
Дивергенция и конвергенция путей. Дивергенция пути - это контактирование одного нейрона со множеством нейронов более высоких порядков.. Так, у позвоночных существует разделение аксона чувствительного нейрона, входящего в спинной мозг, на множество веточек (коллатералей), которые направляются к разным сегментам спинного мозга и в головной мозг, где происходит передача сигнала на вставочные и далее - на моторные нервные клетки.
Дивергенция пути сигнала наблюдается и у вставочных, например командных, клеток, а также выходных нервных клеток (мотонейронов и других эффекторных нейронов). Так, у человека один большой мотонейрон (группы А) иннервирует, т. с. возбуждает десятки мышечных волокон (во внешних глазных мышцах) и даже тысячи (в мышцах конечностей).
Дивергенция пути обеспечивает расширение сферы действия сигнала. Это называют иррадиацией возбуждения (или торможения).
Конвергенция - это схождение многих нервных путей к одним и тем же нейронам.
Например, у позвоночных на каждом мотонейроне спинного мозга и ствола головного мозга образуют синоптические окончания тысячи сенсорных, а также возбуждающих и тормозных вставочных нейронов разных уровней. Мощная конвергенция обнаруживается и на нейронах ретикулярной формации ствола мозга, на многих корковых нейронах у позвоночных и, видимо, на командных нейронах.
Конвергенция многих нервных путей к одному нейрону делает этот нейрон интегратором соответствующих сигналов. Его состояние, т. е. импульсация или "молчание" в каждый данный момент определяются алгебраическим сложением массы возбуждающих и тормозных входов, иными словами, интегралом всех его ВПСП и ТПСП. Такое сложение ПСП называютпространственной или одновременной суммацией. При этом нейрон-интегратор называют общим путем для конвергирующих на него нервных сигналов, и если речь идет о мотонейроне, т. е. конечном звене нервного пути к мускулатуре, говорят об общем конечном пути.
Наличие конвергенции множества путей, т. с. нервных цепочек, на одной группе мотонейронов лежит в основе феноменов пространственного облегчения и окклюзии.
Пространственное облегчение - это превышение эффекта одновременного действия двух относительно слабых афферентных возбуждающих входов в ЦНС над суммой их раздельных эффектов. Феномен объясняется суммацией ВПСП до КУД в группе мотонейронов, имеющих подпороговые ВПСП при раздельной активации входов.
Окклюзия - это явление, противоположное пространственному облегчению. Здесь два сильных (т. с. реализуемых большим числом элементов) афферентных входа вместе возбуждают группу мотонейронов, меньшую той, которая образуется при арифметическом сложении групп мотонейронов, возбуждаемых от этих входов, когда они действуют порознь.
Причина окклюзии состоит в том, что эти афферентные входы в силу конвергенции отчасти адресуются к одним и тем же мотонейронам и каждый может создать в них такое же сверхпороговое возбуждение, как и оба входа вместе.
Использование торможения в нервных сетях. Рассмотрим основные направления тормозных действий в нервных сетях: реципрокное торможение, возвратное торможение, латеральное торможение и прямое взаимное торможение.
Реципрокиое торможение - это взаимное (сопряженное) торможение центров антагонистических рефлексов, обеспечивающее координацию этих рефлексов.
Классический пример реципрокного торможения - это торможение мотонейронов мышц-антагонистов у позвоночных. Торможение осуществляется с помощью специальных тормозных вставочных нейронов. При активации путей, возбуждающих, например, мотонейроны мышц-сгибателей, мотонейроны мышц-разгибателей тормозятся импульсами вставочных клеток. Аналогичное по смыслу торможение, но направленное прямо на мышцы, наблюдается у ракообразных .
Возвратное торможение - это торможение нейронов собственными импульсами, поступающими по возвратным коллатералям к тормозным клеткам.
Возвратное торможение наблюдается, например, в мотонейронах спинного мозга позвоночных. Эти клетки отдают возвратные коллатерали в мозг к тормозным вставочным клеткам Реншоу, которые имеют синапсы на этих же мотонейронах. Торможение обеспечивает ограничение ритма мотонейронов, что важно для нормальной работы двигательного аппарата. Такую же роль играет возвратное торможение и в других нервных сетях .
Латеральное торможение - это торможение элементов соседних нервных цепочек в конкурирующих сенсорных каналах связи .
Латеральное торможение, видимо, также осуществляется с помощью тормозных вставочных клеток. Такое торможение четко показано у мечехвоста в сенсорных цепочках, идущих от фасеточного глаза к ЦНС. Оно наблюдается у соседних элементов сетчатки позвоночных, а также в их зрительных, слуховых и других сенсорных центрах.
Во всех случаях латеральное торможение обеспечивает контраст, т е выделение существенных сигналов из фона.
Прямым взаимным торможением можно назвать тормозное взаимодействие двух (или большего числа) командных нейронов, осуществляющееся без специальных вставочных клеток.
Такое торможение предполагается у командных клеток L-10 и интернейрона II у моллюска аплизии. Прямое взаимное торможение в паре клеток делает эту пару склонной функционировать по принципу "или - или". Иначе говоря, такая "метастабильная" сеть разрешает работу либо клетке L-10, либо клетке II в силу того обстоятельства, что активная клетка прямо тормозит конкурента, чем освобождает себя от торможения с его стороны.
Предполагают, что подобные отношения существуют и между командными нейронами мозга высших животных и человека, обеспечивающими поведенческие и эмоциональные установки по принципу доминанты .
Самовозбуждающиеся нервные цепи. Некоторые данные свидетельствуют о том, что в мозгу животных и человека существуют самовозбуждающиеся цепочки нейронов, т. е. такие замкнутые цепочки, в которых нейроны соединены синапсами возбуждающего действия . Возникнув в ответ на внешний сигнал, возбуждение в такой цепочке циркулирует, иначе реверберирует, до тех пор, пока или какой-либо внешний тормоз не выключит одно из звеньев цепи, или в ней не наступит утомление. Выходные пути от такой цепочки (ответвляющиеся по коллатералям аксонов нервных клеток - участников цепи) во время работы передают равномерный поток импульсов, создающий ту или иную настройку в нервных клетках-мишенях.
Таким образом, самовозбуждающаяся цепочка, пока она работает, как бы "помнит" тот краткий сигнал, который включил в ней циркуляцию (реверберацию) импульсов. Считают, что это возможный механизм (или один из механизмов) краткосрочной памяти.
Наши сведения о мозге растут быстро. Однако физиолог не может чувствовать полного удовлетворения, так как основной вопрос, как же устроен мозг, остается открытым. Главное в работе мозга — это переработка информации. Переработка информации составляет основу таких процессов, как мышление, узнавание, обучение. Но именно механизмы этих явлений наиболее трудны для исследования.
В основе активного состояния и сна человека, в основе его эмоций лежит одновременное возбуждение или торможение целых отделов мозга, включающих многие тысячи нейронов. Эти процессы можно исследовать электрофизиологически, путем удаления или раздражения участков мозга. Поэтому в изучении этих явлений достигнуты успехи. В основе же переработки информации лежит работа сложнейших конструкций нейронов — нервных сетей, где в доли секунды возникают и исчезают сложнейшие мозаики из возбужденных и заторможенных нервных клеток, которые очень трудно уловить и проанализировать.
Так же как школьник не сможет понять устройства вычислительной машины, наблюдая за вспыхиванием отдельных ламп, так же трудно без знания общей теории переработки информации разобраться в работе нервных сетей головного мозга.
В настоящее время появился очень хороший критерий нашего понимания механизмов физиологических процессов. Этот критерий — возможность моделирования. Директор Института кибернетики в Киеве академик В. М. Глушков подчеркивает, что если исследователи действительно понимают механизмы физиологических процессов, то их уже сейчас можно воспроизвести в вычислительных машинах и воспроизвести явление в модели. Однако современные знания о головном мозге не удовлетворяют этим условиям. Как же подойти к разгадке тайны работы нервных сетей?
Но если ученые не могут соединять живые нервные клетки, то они могут при современном уровне науки сделать то же самое с их электронными моделями. Можно создать модели нейронов и, собирая из них сети, изучать свойства и законы функционирования этих сетей. Так и поступают многие ученые.
Сейчас в нейрокибернетике делается еще более глубокий заход с тыла. Дело в том, что с теоретической точки зрения нервная клетка — это далеко не элементарная единица, перерабатывающая информацию. Это скорее узел, равноценный по сложности устройству телевизора или радиоприемника. Для теории же важно иметь дело с простейшими компонентами. Таких компонентов немного.
Развивается теория автоматов — паука, во многих чертах подобная геометрии. Так же, как геометрия, она оперирует абстрактными понятиями и развивает теорию, казалось бы, независимо от изучения мозга. Однако так же, как выводы из геометрии оказываются применимыми и необходимыми в весьма различных областях практической деятельности человека, так же и кибернетическая теория оказывается применимой при изучении всех систем, перерабатывающих информацию. А мозг — именно такая система.
У ученых возникла мысль: нельзя ли, используя все то, что сейчас известно о мозге, и опираясь на кибернетическую теорию работы систем, перерабатывающих информацию, попытаться представить, как должна быть организована нервная сеть, способная к различным сложным формам деятельности?
Оказалось, что это вполне возможно. Так родилась теория нервной сети. Эта теория имеет такое же соотношение с экспериментальным изучением мозга, какое существует между теоретической и экспериментальной физикой.
Первые работы в этом направлении были осуществлены американским ученым Н. Рашевским, который построил интересные нервные сети и математически доказал, что эти системы способны к довольно сложным формам деятельности.
Доказано, что нервные сети, состоящие из простых элементов, в целом оказываются способными к обучению, решению проблем, узнаванию образов и выработке понятий. Например, в Московском энергетическом институте под руководством Ю. Н. Кушелева, а затем в Германии профессором Штайнбухом была создана теория самообучающихся матриц. Эта теория объясняет, как должны быть организованы структуры, которые способны к приобретению опыта и к формированию целесообразного поведения в новых условиях.
На основе этой теории уже созданы самообучающиеся автоматы, успешно – управляющие работой химических заводов.
Итак, сведения ученых об организации мозга сосредоточиваются в настоящее время вокруг двух полюсов. С одной стороны, известно, какие именно структуры могут самообучаться, решать проблемы и так далее. На другом полюсе собираются экспериментальные факты о структурах мозга.
Следующим этапом науки о мозге должен стать, очевидно; более тесный синтез этих двух направлений. Кибернетическая теория должна стать базой для проведения специальных экспериментов, подтверждающих те или иные гипотезы. Может быть, на первых этапах и не будет детального совпадения структур мозга и теоретически рассчитанных структур. Однако будут подтверждены и найдены некоторые общие принципы.
Следует подчеркнуть важность взаимного обогащения обоих направлений. Каждое новое открытие в физиологии обогащает теорию нервных сетей. Сейчас уже ясно, что необходимо создать более сложную и совершенную теорию, чем та, которую создали в свое время, например, Мак Каллок и Пите. Большое значение при этом имеет изучение тех клеток, которые находятся между нейронными сетями (нейроглии). Роль этих клеток, видимо, необходимо учитывать при построении любой теории. Интересны данные об объемных соотношениях компонентов коры головного мозга: 30% объема коры составляют тела нейронов с дендритами, и 70% объема занимают клетки глин и кровеносные сосуды.
По происхождению все элементы нейроглии (греч. neuron — нейрон, gli — клей) делятся на два вида: макроглию и микроглию.
Но нас в основном будут интересовать клетки макроглии, которые уже в период своего возникновения тесно связаны с нейронами, одновременно с ними развиваясь из эмбрионального зачатка нервной системы — нервной трубки. И именно взаимодействие между нейронами и макроглией в течение последних лет интересует многих исследователей.
Тела нейронов, их дендриты и аксоны как бы взвешены в массе глиальных клеток. Отростки глиальных клеток, густо оплетая нейрон, образуют вокруг него своего рода капсулы, которые, очевидно, изолируют нейрон от прямого контакта с кровеносной системой. Вступая во внутриклеточный контакт с телом нейрона и капиллярами кровеносной системы, клетки глии являются транспортной или опосредующей системой между капиллярами и нервными клетками. Но клетки глии не только доставляют необходимые вещества нейронам — макроглия подвергает эти вещества соответствующей обработке, так что на долю нейронов остаются только последние этапы, включая синтез непосредственных донаторов энергии.
В какой же степени клетки глии, а не нейроны ответственны за изученные физиологические процессы? Существует много экспериментальных данных, для объяснения которых можно было бы прибегнуть к рассмотрению взаимодействия между глией и нейронами. Так, например, интересны опыты по охлаждению или данные о зимней спячке животных, во время которой активность нервной системы приближается к нулю, но после пробуждения сложное поведение подверженных охлаждению животных полностью восстанавливается.
Но, видимо, следовало бы ученым поинтересоваться, в какой мере клетки глии, а не нейроны реагируют на электрическое раздражение, на введение в организм различных фармакологических веществ, на появление в крови всевозможных гормонов. И есть ли необходимость рассматривать функционирование нейронных сетей, а не взаимодействие между нейронами и глией? Нам не удается сейчас в должной мере объяснить многие акты поведения, которые формируются и длятся иногда часы, годы, поколения, исходя из наших знаний о нейронной активности, длящейся миллисекунды, секунды, минуты.
Временные характеристики глиальной активности отличаются от нейронной. Потенциалы, отведенные от одного из видов глиальных клеток (астроглии), имели примерно в тысячу раз большую длительность, чем потенциалы нейронов, и вполне вероятно, что нейрон, окруженный ажурными капсулами из глии, испытывает на себе влияние медленно изменяющейся электрической активности нейроглии, которая направляет активность нервных клеток.
Роль глии как программирующего устройства в деятельности нервных клеток частично была вскрыта в экспериментах по изучению регенерации периферических нейронов. Полагают, что глия служит главной направляющей структурой для роста регенерирующих аксонов. В массе клеток среди множества различных биохимических комплексов клетки глии великолепно узнают и прокладывают для растущего аксона нужные пути, направляют установление необходимых связей.
Об этом, например, говорит и то, что попытки моделирования нервных сетей оказались менее эффективными, чем предполагалось ранее. Вряд ли удастся добиться сложного поведения от системы, сконструированной из элементов одного уровня, из однотипных, универсальных единиц. Дело, по всей вероятности, заключается не только в способах соединений этих элементов в системы.
Развитие теории нервных сетей идет в настоящее время по различным направлениям. Так, еврейский ученый А. Б. Коган считает, что трудно представить себе, что в нервных сетях, функционирующих в головном мозге, предопределено, как именно каждый нейрон связан с другими соседними, то есть предопределена вся схема (структура) строения. А. Б. Коган думает, что взаимодействие нейронов носит вероятностный характер.
Эти идеи подтверждены экспериментами на живых организмах. Помимо того, были созданы электронные модели, на которых было доказано, что действительно такие нервные сети обладают в целом многими свойствами, например, способностью к узнаванию, способностью к выработке условных рефлексов.
Американский ученый Розенблат также считает, что важно выяснить какой-то общий принцип организации и соединения нейронов, а не создавать схемы, в которых предопределена каждая связь между нервными клетками.
Авторы: А. Напалков, А. Туров.
P. S. О чем еще говорят британские ученые: о том, что научные достижения в сфере нервных сетей вполне могли бы войти в ТОПы и интересные факты из сферы науки.

Нервная система регулирует деятельность всех органов и систем, обусловливая их функциональное единство, и обеспечивает связь организма как целого с внешней средой (увеличить рисунок)
Структурной единицей нервной системы является нервная клетка с отростками - нейрон. Bся нервная система представляет собой совокупность нейронов, которые контактируют друг с другом при помощи специальных аппаратов - синапсов. По структуре и функции различают три типа нейронов:
- рецепторные, или чувствительные;
- вставочные, замыкательные (кондукторные);
- эффекторные, двигательные нейроны, от которых импульс направляется к рабочим органам (мышцам, железам).
Нервная система условно подразделяется на два больших отдела - соматическую, или анимальную, нервную систему и вегетативную, или автономную, нервную систему. Соматическая нервная система осуществляет преимущественно функции связи организма с внешней средой, обеспечивая чувствительность и движение вызывая сокращение скелетной мускулатуры. Так как функции движения и чувствования свойственны животным и отличают их от растений, эта часть нервной системы получила название анимальной (животной).
В нервной системе выделяют центральную часть - головной и спинной мозг - центральная нервная система и переферическую, представленную отходящими от головного и спинного мозга нервами, - переферическая нервная система. На разрезе мозга видно, что он состоит из серого и белого вещества.
Серое вещество образуется скоплениями нервных клеток ( с начальными отделами отходящих от их тел отростков). Отдельные ограниченные скопления серого вещества носят названия ядер.
Белое вещество образуют нервные волокна, покрытые миелиновой оболочкой (отростки нервных клеток, образующих серое вещество). Нервные волокна в головном и спинном мозге образуют проводящие пути.
И.П. Павлов показал, что центральная нервная система может оказывать три рода воздействий на органы:
- 1) пусковое, вызывающее либо прекращающее функцию органа (сокращение мышцы, секрецию железы);
- 2) сосудодвигательное, изменяющее ширину просвета сосудов и тем самым регулирующее приток к органу крови;
- 3) трофическое, повышающее или понижающее обмен веществ и, следовательно потребление питательных веществ и кислорода. Благодаря этому постоянно согласуется функциональное состояние ргана и его потребность в питательных веществах и кислороде. Когда к работающей скелетной мышце по двигательным волокнам направляются импульсы, вызывающие ее сокращение, то одновременно по вегетативным нервным волокнам поступают импульсы, расширяющие сосуды и у силивающие обмен веществ. Тем самым обеспечивается энергетическая возможность выполнения мышечной работы.
Центральная нервная система воспринимает афферентную (чувствительную) информацию, возникающую при раздражении спецефических рецепторов и в ответ на это формирует соответствующие эфферентные импульсы, вызывающие изменения в деятельности определнных органов и систем организма.
В основе организации нейронных сетей лежат процессы дивергенции (от лат. diverge — отхожу) и конвергенции (от лат. converge — схожусь). Так, разветвления аксона одного нейрона могут подходить к нескольким нейронам, передавая им свои сигналы одновременно (дивергенция), или к одному нейрону подходить разветвления аксонов нескольких нейронов (конвергенция). Переработка информации в нервных се-

Рис. 4.17. Типы нейронных сетей:
а — пример сети нейронов; 6 — нейроэндокринная; в — локальная сеть; г — сети с отдаленными связями; д — распределительные системы тях может происходить также за счет процессов интеграции (от лат. integer — целый), или объединения, различного рода потоков нервных импульсов.
Наиболее простые сети нервной системы — локальные и диффузные. Локальные, или микросети (см. рис. 4.17), объединяют небольшое число близко расположенных нейронов, лежащих в пределах одного ядра или его части, и обеспечивают местную обработку информации. Диффузные сети образованы нейронами, не собранными в ядра, но лежащими вблизи друг от друга (сетчатая, или ретикулярная, формация). Простые нейронные сети могут входить в состав более сложных.
Нейроны могут лежать на большом расстоянии друг от друга, но в пределах одной области мозга. Функции таких сетей — распространение информации за пределы небольшого локального участка или обеспечение реципрокных (противоположных: возбуждение — торможение) отношений между нервными клетками в пределах одной области. Таким образом взаимодействуют нейроны ядер черепно-мозговых нервов в стволе головного мозга или нейроны коры больших полушарий (рис. 4.18, а).
Нейроны, расположенные в разных структурах мозга, объединяются в сети с отдаленными связями, для которых характерна более сложная организация: каждая такая сеть име-

Рис. 4.18. Сети нейронов коры головного мозга человека (я); распределительная система с участием нейронов коры больших полушарий, базальных ядер, таламуса, черной субстанции (б)
ет несколько входов и выходов. Примером могут служить многочисленные связи мозжечка со стволовыми структурами. Различного рода информация обрабатывается в таких сетях по тем же принципам конвергенции, дивергенции и интеграции. Разновидностью этих сетей являются последовательно соединенные нейроны нескольких структур мозга. Такие сети образуют восходящие и нисходящие проводящие пути или целые системы мозга (сенсорные, моторные, центральные). Их функция — передача и обработка информации, поступающей в мозг (сенсорные системы) или из мозга на периферию (моторные системы). Самые сложно организованные нервные сети — распределительные системы (рис. 4.18, б). Они образованы нейронами, относящимися к функционально разным структурам мозга (сенсорным, моторным, центральным). Их предназначение — организация целенаправленного поведения, в котором участвует весь организм.

Здравствуй, Geektimes! В ранее опубликованной статье, была представлена модель нервной системы, опишу теорию и принципы, которые легли в её основу.
Теория основана на анализе имеющейся информации о биологическом нейроне и нервной системе из современной нейробиологии и физиологии мозга.
Сначала приведу краткую информацию об объекте моделирования, вся информация изложена далее, учтена и использована в модели.
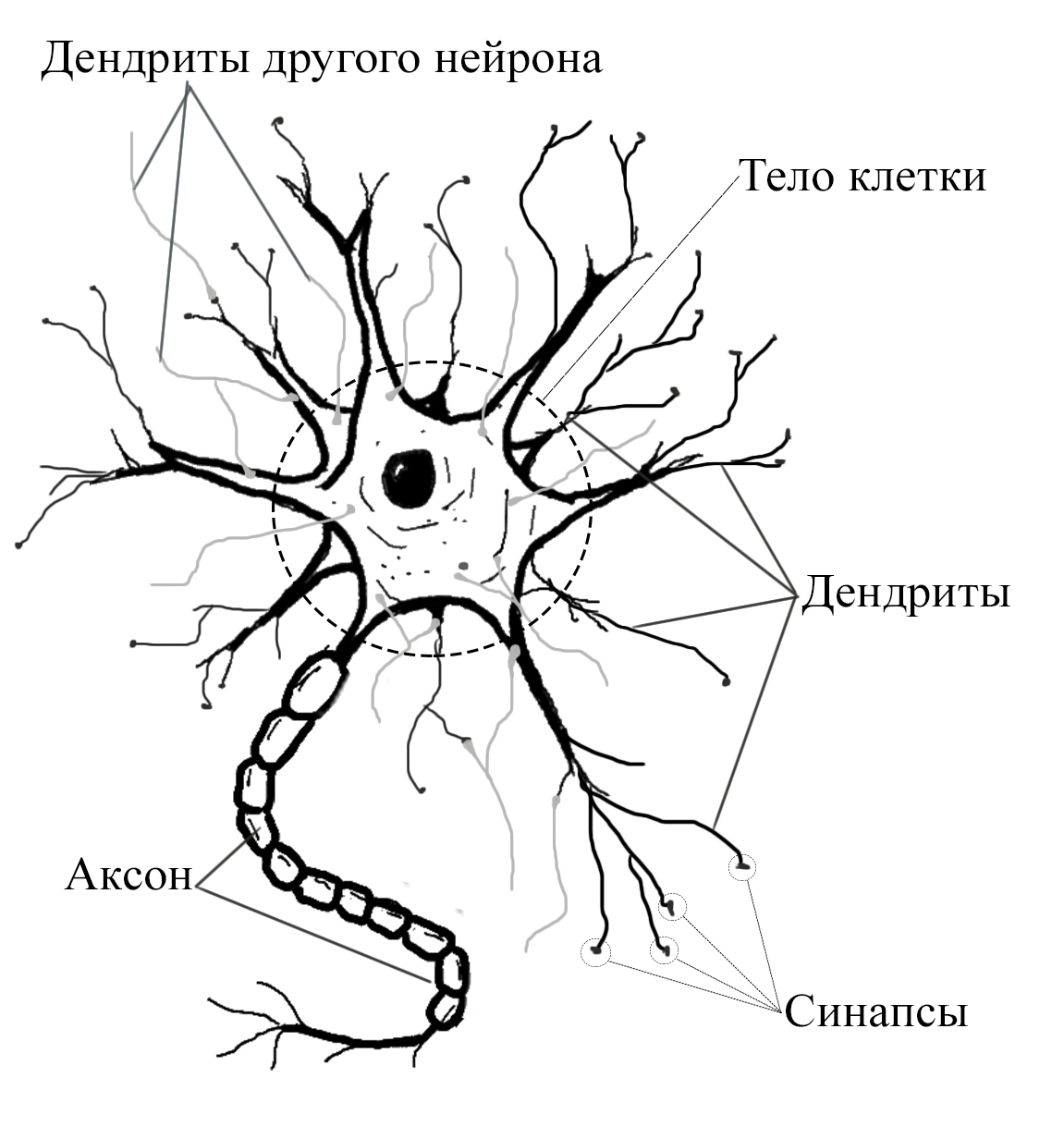
Нейрон является основным функциональным элементом нервной системы, он состоит из тела нервной клетки и её отростков. Существуют два вида отростков: аксоны и дендриты. Аксон – длинный покрытый миелиновой оболочкой отросток, предназначенный для передачи нервного импульса на далекие расстояния. Дендрит – короткий, ветвящийся отросток, благодаря которым происходит взаимосвязь с множеством соседних клеток.
Нейроны могут сильно отличаться по форме, размерам и конфигурации, не смотря на это, отмечается принципиальное сходство нервной ткани в различных участках нервной системе, отсутствуют и серьезные эволюционные различия. Нервная клетка моллюска Аплизии может выделять такие же нейромедиаторы и белки, что и клетка человека.
В зависимости от конфигурации выделяют три типа нейронов: 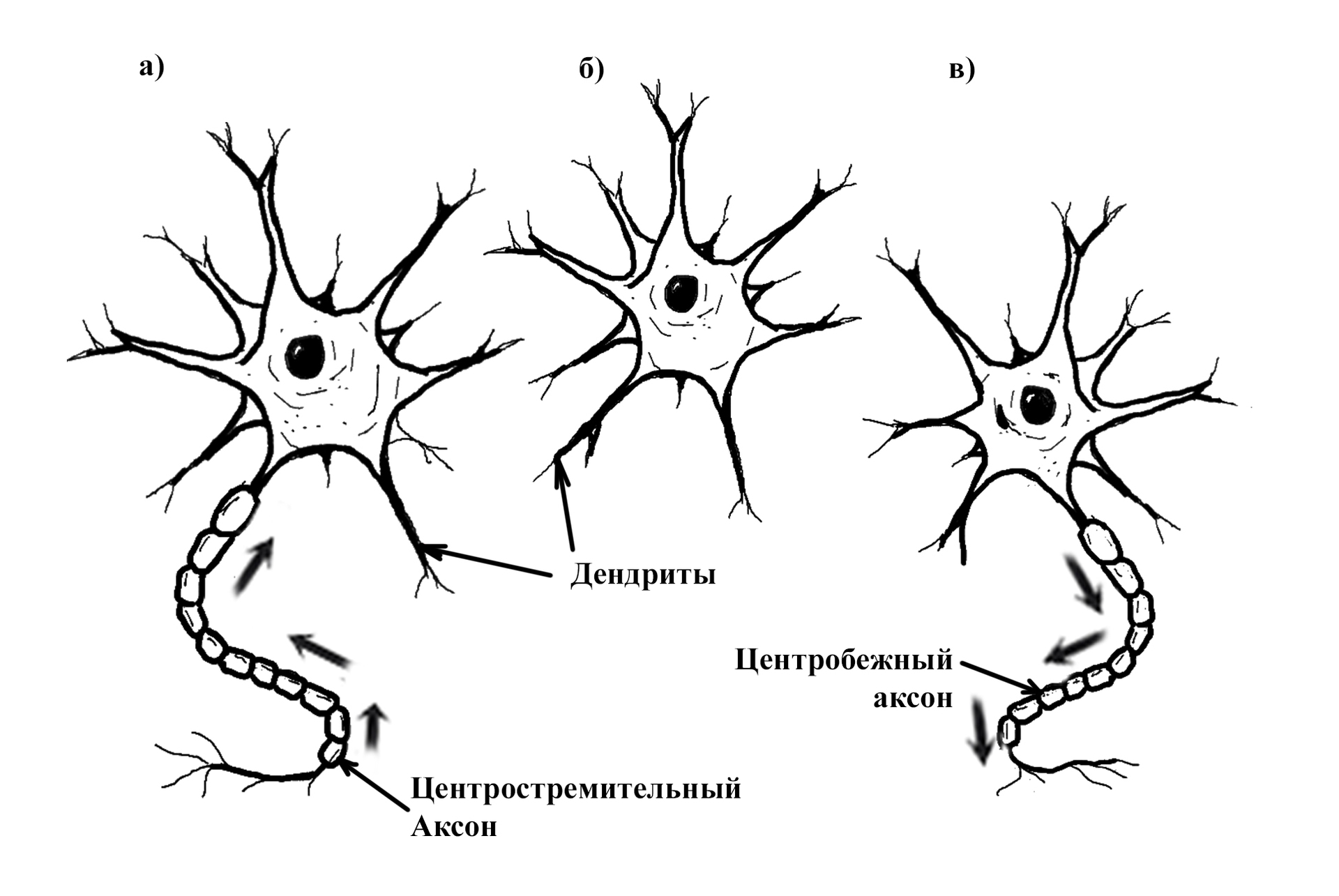
а) рецепторные, центростремительные, или афферентные нейроны, данные нейроны имеют центростремительный аксон, на конце которого имеются рецепторы, рецепторные или афферентные окончания. Эти нейроны можно определить, как элементы, передающие внешние сигналы в систему.
б) интернейроны (вставочные, контактные, или промежуточные) нейроны, не имеющие длинных отростков, но имеющие только дендриты. Таких нейронов в человеческом мозгу больше чем остальных. Данный вид нейронов является основным элементом рефлекторной дуги.
в) моторные, центробежные, или эфферентные, они имеют центростремительный аксон, который имеет эфферентные окончания передающий возбуждение мышечным или железистым клеткам. Эфферентные нейроны служат для передачи сигналов из нервной среды во внешнюю среду.
Передача возбуждения происходит от нейрона к нейрону, через специальные утолщения на концах дендритов, называемых синапсами. По типу передачи синапсы разделяют на два вида: химические и электрические. Электрические синапсы передают нервный импульс непосредственно через место контакта. Таких синапсов в нервных системах очень мало, в моделях не будут учитываться. Химические синапсы передают нервный импульс посредством специального вещества медиатора (нейромедиатора, нейротрансмиттера), данный вид синапса широко распространен и подразумевает вариативность в работе.
Важно отметить, что в биологическом нейроне постоянно происходят изменения, отращиваются новые дендриты и синапсы, возможны миграции нейронов. В местах контактов с другими нейронами образуются новообразования, для передающего нейрона — это синапс, для принимающего — это постсинаптическая мембрана, снабжаемая специальными рецепторами, реагирующими на медиатор, то есть можно говорить, что мембрана нейрона — это приемник, а синапсы на дендритах — это передатчики сигнала.
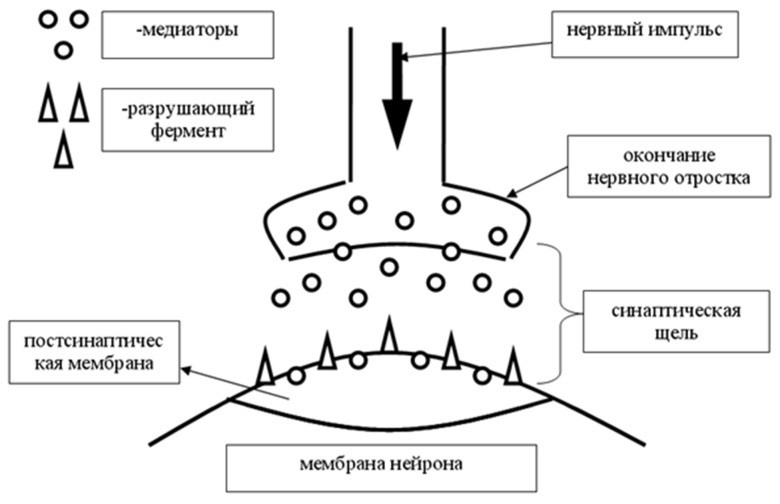
При активации синапса он выбрасывает порции медиатора, эти порции могут варьироваться, чем больше выделится медиатора, тем вероятнее, что принимаемая сигнал нервная клетка будет активирована. Медиатор, преодолевая синоптическую щель, попадает на постсинаптическую мембрану, на которой расположены рецепторы, реагирующие на медиатор. Далее медиатор может быть разрушен специальным разрушающим ферментом, либо поглощен обратно синапсом, это происходит для сокращения времени действия медиатора на рецепторы.
Так же помимо побудительного воздействия существуют синапсы, оказывающие тормозящее воздействие на нейрон. Обычно такие синапсы принадлежат определенным нейронам, которые обозначаются, как тормозящие нейроны.
Синапсов связывающих нейрон с одной и той же целевой клеткой, может быть множество. Для упрощения примем, всю совокупность, оказываемого воздействия одним нейроном, на другой целевой нейрон за синапс с определённой силой воздействия. Главной характеристикой синапса будет, является его сила.
В состоянии покоя мембрана нейрона поляризована. Это означает, что по обе стороны мембраны располагаются частицы, несущие противоположные заряды. В состоянии покоя наружная поверхность мембраны заряжена положительно, внутренняя – отрицательно. Основными переносчиками зарядов в организме являются ионы натрия (Na+), калия (K+) и хлора (Cl-).
Разница между зарядами поверхности мембраны и внутри тела клетки составляет мембранный потенциал. Медиатор вызывает нарушения поляризации – деполяризацию. Положительные ионы снаружи мембраны устремляются через открытые каналы в тело клетки, меняя соотношение зарядов между поверхностью мембраны и телом клетки.
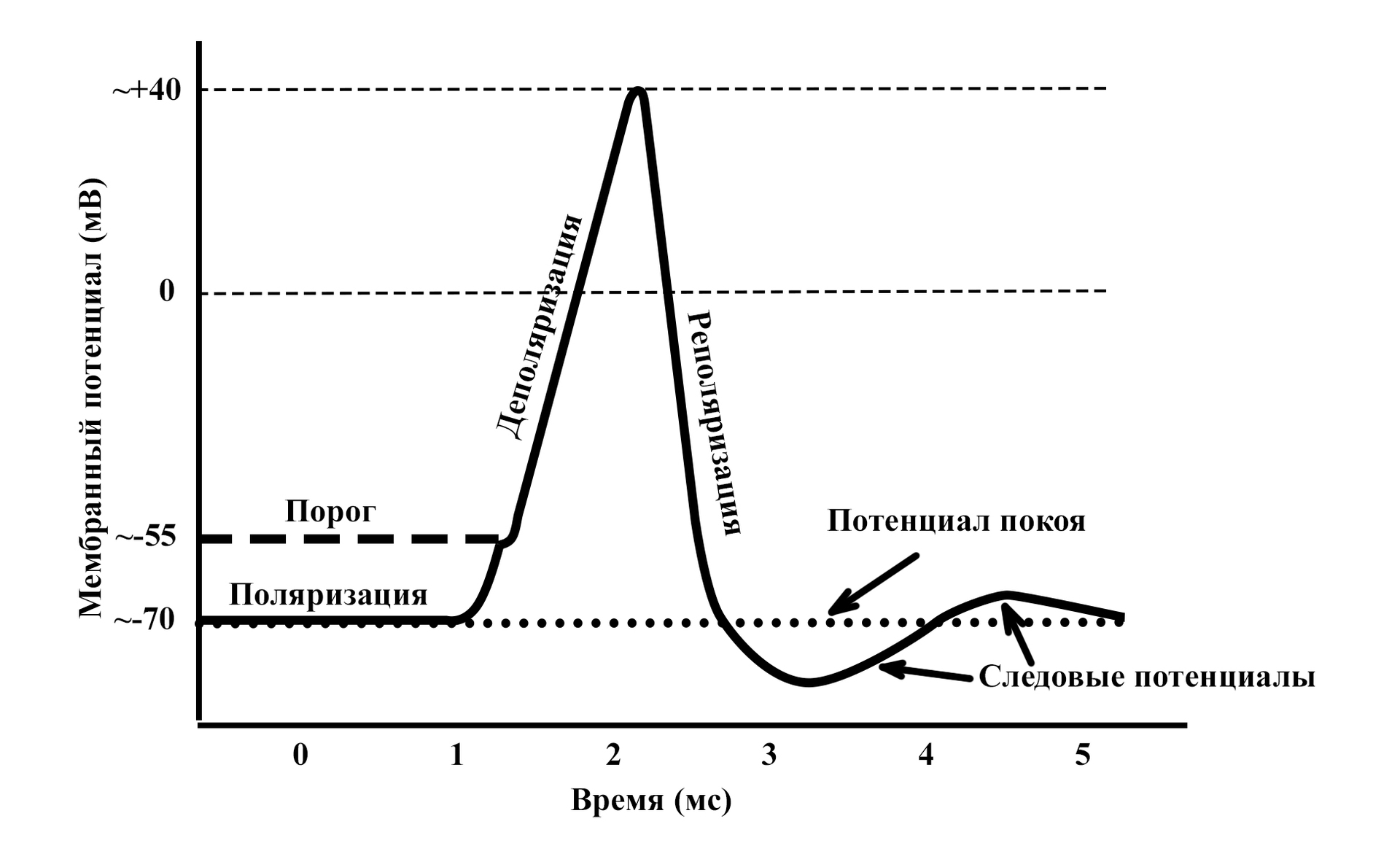
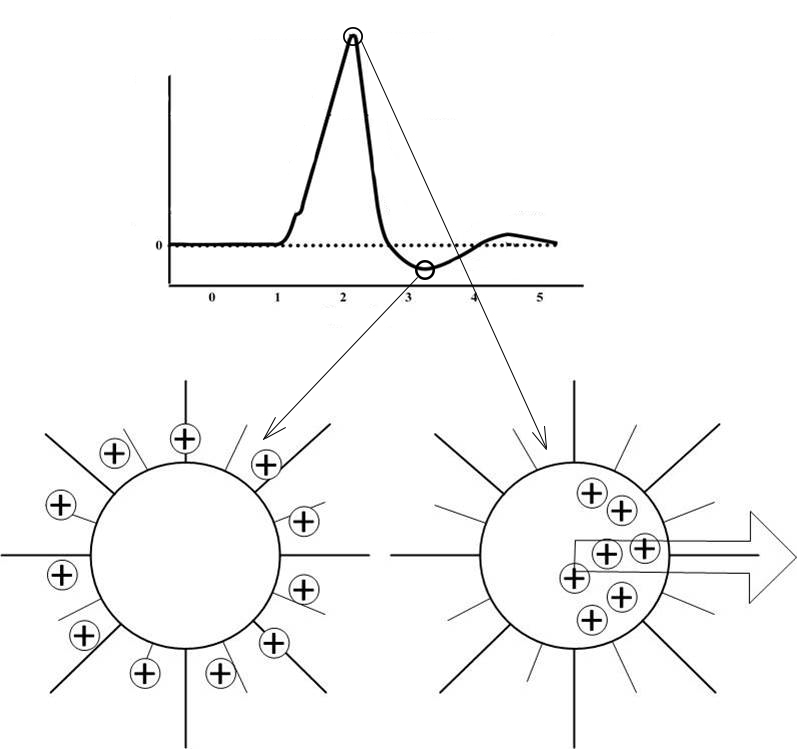
Изменение мембранного потенциала при возбуждении нейрона
Характер изменений мембранного потенциала при активации нервной ткани неизменен. Независимо от того кокой силы воздействия оказывается на нейрон, если сила превышает некоторое пороговое значение, ответ будет одинаков.
Забегая вперед, хочу отметить, что в работе нервной системы имеет значение даже следовые потенциалы (см. график выше). Они не появляются, вследствие каких-то гармонических колебаний уравновешивающих заряды, являются строгим проявлением определённой фазы состояния нервной ткани при возбуждении.
Итак, далее приведу теоретические предположения, которые позволят нам создавать математические модели. Главная идея заключается во взаимодействии между зарядами формирующихся внутри тела клетки, во время её активности, и зарядами с поверхностей мембран других активных клеток. Данные заряды являются разноименными, в связи этим можно предположить, как будут располагаться заряды в теле клетки под воздействием зарядов других активных клеток.
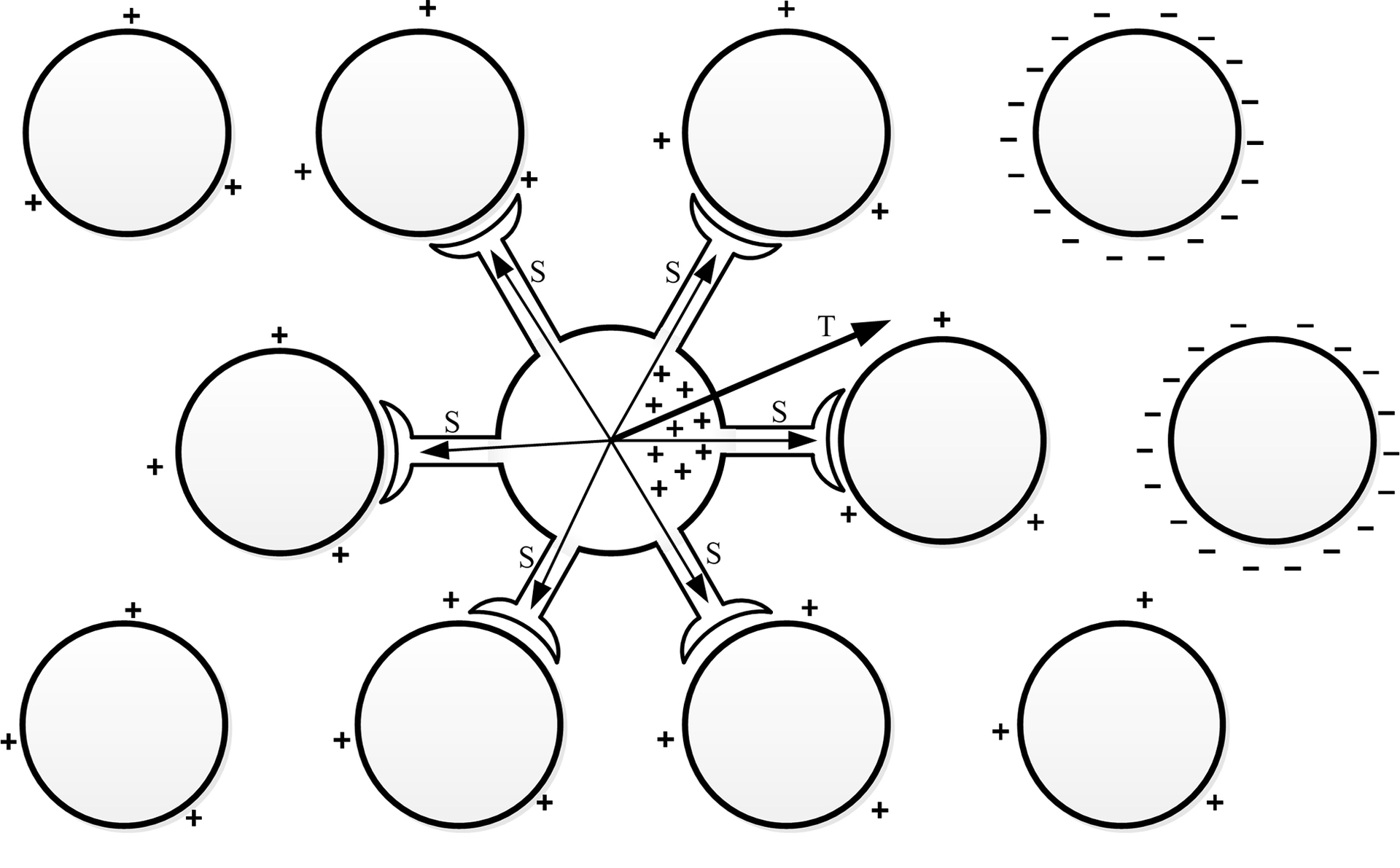
Можно сказать, что нейрон чувствует активность других нейронов на расстоянии, стремится направить распространения возбуждения в направлении других активных участков.
В момент активности нейрона можно рассчитать определённую точку в пространстве, которая определялась бы, как сумма масс зарядов, расположенных на поверхностях других нейронов. Указанную точку назовем точкой паттерна, её месторождение зависит от комбинации фаз активности всех нейронов нервной системы. Паттерном в физиологии нервной системы называется уникальная комбинация активных клеток, то есть можно говорить о влиянии возбуждённых участков мозга на работу отдельного нейрона.
Нужно представлять работу нейрона не просто как вычислителя, а своего рода ретранслятор возбуждения, который выбирает направления распространения возбуждения, таким образом, формируются сложные электрические схемы. Первоначально предполагалось, что нейрон просто избирательно отключает/включает для передачи свои синапсы, в зависимости от предпочитаемого направления возбуждения. Но более детальное изучение природы нейрона, привело к выводам, что нейрон может изменять степень воздействия на целевую клетку через силу своих синапсов, что делает нейрон более гибким и вариативным вычислительным элементом нервной системы.
Какое же направление для передачи возбуждения является предпочтительным? В различных экспериментах связанных с образованием безусловных рефлексов, можно определить, что в нервной системе образуются пути или рефлекторные дуги, которые связывают активируемые участки мозга при формировании безусловных рефлексов, создаются ассоциативные связи. Значит, нейрон должен передавать возбуждения к другим активным участкам мозга, запоминать направление и использовать его в дальнейшем.
Представим вектор начало, которого находится в центре активной клети, а конец направлен в точку паттерна определённую для данного нейрона. Обозначим, как вектор предпочитаемого направления распространения возбуждения (T, trend). В биологическом нейроне вектор Т может проявляться в структуре самой нейроплазмы, возможно, это каналы для движения ионов в теле клетки, или другие изменения в структуре нейрона.
Нейрон обладает свойством памяти, он может запоминать вектор Т, направление этого вектора, может меняться и перезаписываться в зависимости от внешних факторов. Степень с которой вектор Т может подвергается изменениям, называется нейропластичность.
Этот вектор в свою очередь оказывает влияние на работу синапсов нейрона. Для каждого синапса определим вектор S начало, которого находится в центре клетки, а конец направлен в центр целевого нейрона, с которым связан синапс. Теперь степень влияния для каждого синапса можно определить следующим образом: чем меньше угол между вектором T и S, тем больше синапс будет, усиливается; чем меньше угол, тем сильнее синапс будет ослабевать и возможно может прекратить передачу возбуждения. Каждый синапс имеет независимое свойство памяти, он помнит значение своей силы. Указанные значения изменяются при каждой активизации нейрона, под влиянием вектора Т, они либо увеличиваются, либо уменьшаются на определённое значение.
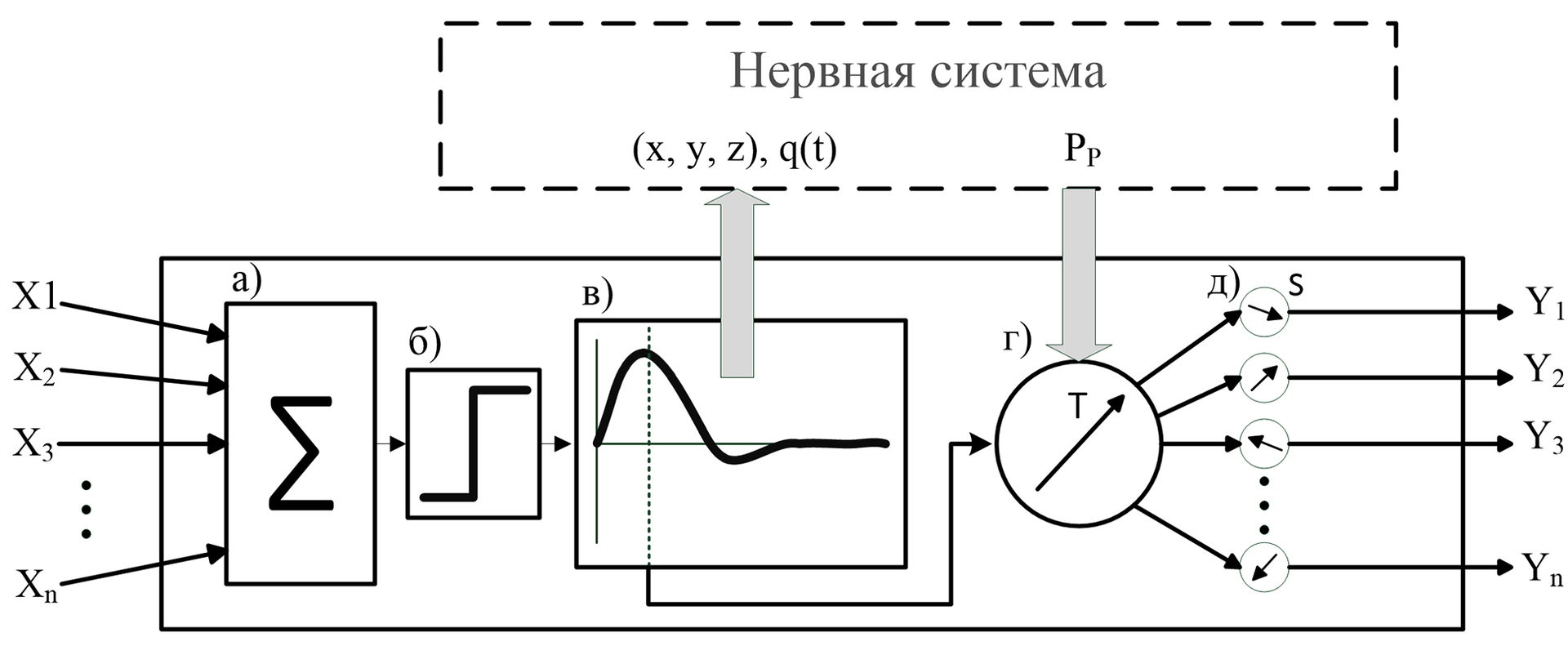
Входные сигналы (x1, x2,…xn) нейрона представляют собой вещественные числа, которые характеризуют силу синапсов нейронов, оказывающих воздействие на нейрон.
Положительное значение входа означает побудительное воздействие, оказываемое на нейрон, а отрицательное значение – тормозящее воздействие.
Для биологического нейрона не имеет значение, откуда поступил возбуждающий его сигнал, результат его активности будет идентичен. Нейрон будет активизирован, когда сумма воздействий на него будет превышать определённое пороговое значение. Поэтому, все сигналы проходят через сумматор (а), а поскольку нейроны и нервная система работают в реальном времени, следовательно, воздействие входов должно оцениваться в короткий промежуток времени, то есть воздействие синапса имеет временный характер.
Результат сумматора проходит пороговую функцию (б), если сумма превосходит пороговое значение, то это приводит к активности нейрона.
При активации нейрон сигнализирует о своей активности системе, передовая информацию о своём положении в пространстве нервной системы и заряде, изменяемом во времени (в).
Через определённое время, после активации нейрон передает возбуждение по всем имеющимся синапсам, предварительно производя пересчет их силы. Весь период активации нейрон перестает реагировать на внешние раздражители, то есть все воздействия синапсов других нейронов игнорируются. В период активации входит так же период восстановления нейрона.
Происходит корректировка вектора Т (г) с учётом значения точки паттерна Pp и уровнем нейропластичности. Далее происходит переоценка значений всех сил синапсов в нейроне(д).
Обратите внимание, что блоки (г) и (д) выполняются параллельно с блоком (в).
Если внимательно проанализировать предложенную модель, то можно увидеть, что источник возбуждения должен оказывать большее влияние на нейрон, чем другой удалённый, активный участок мозга. Следовательно возникает вопрос: почему же все равно происходит передача в направлении другого активного участка?
Данную проблему я смог определить, только создав компьютерную модель. Решение подсказал график изменения мембранного потенциала при активности нейрона.
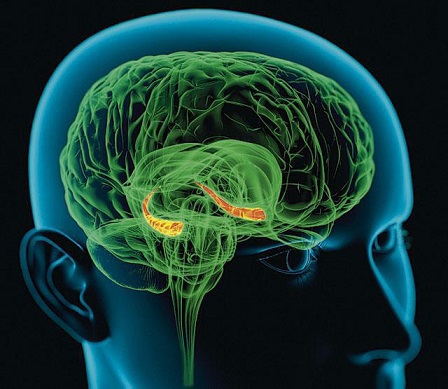
Используя теорию электромагнитного взаимодействия, можно объяснить многие явления и сложные процессы, протекающие в нервной системе. К примеру, одним из последних открытий, которое широко обсуждается в науках о мозге, является открытие когнитивных карт в гиппокампе.
Гиппокамп – это отдел мозга, которому отвечает за кратковременную память. Эксперименты на крысах выявили, что определённому месту в лабиринте соответствует своя локализованная группа клеток в гиппокампе, причем, не имеет значение, как животное попадает в это место, все равно будет активирован соответствующий этому месту участок нервной ткани. Естественно, животное должно помнить данный лабиринт, не стоит рассчитывать на топологическое соответствие пространства лабиринта и когнитивной карты.
Каждое место в лабиринте представляется в мозге, как совокупность раздражителей различного характера: запахи, цвет стен, возможные примечательные объекты, характерные звуки и т. д. Указанные раздражители отражаются на коре, различных представительствах органов чувств, в виде всплесков активности в определённых комбинациях. Мозг одновременно обрабатывает информацию в нескольких отделах, зачастую информационные каналы разделяются, одна и та же информация поступает в различные участки мозга.

Активация нейронов места в зависимости от положения в лабиринте (активность разных нейронов показана разным цветом). источник
Гиппокамп расположен в центре мозга, вся кара и её области удалены от него, на одинаковые расстояния. Если определить для каждой уникальной комбинации раздражителей точку масс зарядов поверхностей нейронов, то можно увидеть, что указанные точки будут различны, и будут находиться примерно в центре мозга. К этим точкам будет стремиться и распространятся возбуждение в гиппокампе, формируя устойчивые участки возбуждения. Более того, поочередная смена комбинаций раздражителей, будет приводить к смещению точки паттерна. Участки когнитивной карты будут ассоциативно связываться друг с другом последовательно, что приведет к тому, что животное, помещенное в начало знакомого ей лабиринта, может вспомнить весь последующий путь.
У многих возникнет вопрос, где в данной работе предпосылки к элементу разумности или проявления высшей интеллектуальной деятельности?
Важно отметить, что феномен человеческого поведения, есть следствие функционирования биологической структуры. Следовательно, чтобы имитировать разумное поведение, необходимо хорошо понимать принципы и особенности функционирования биологических структур. К сожалению, в науке биологии пока не представлен четкий алгоритм: как работает нейрон, как понимает, куда необходимо отращивать свои дендриты, как настроить свои синапсы, что бы в нервной системе смог сформироваться простой условный рефлекс, на подобие тех, которые демонстрировал и описывал в своих работах академик И.П. Павлов.
С другой стороны в науке об искусственном интеллекте, в восходящем (биологическом) подходе, сложилось парадоксальная ситуация, а именно: когда используемые в исследованиях модели основаны на устаревших представлениях о биологическом нейроне, консерватизм, в основе которого берётся персептрон без переосмысления его основных принципов, без обращения к биологическому первоисточнику, придумывается все более хитроумные алгоритмы и структуры, не имеющих биологических корней.
Конечно, никто не уменьшает достоинств классических нейронных сетей, которые дали множество полезных программных продуктов, но игра с ними не является путем к созданию интеллектуально действующей системы.
Более того, не редки заявления, о том, что нейрон подобен мощной вычислительной машине, приписывают свойство квантовых компьютеров. Из-за этой сверхсложности, нервной системе приписывается невозможность её повторения, ведь это соизмеримо с желанием смоделировать человеческую душу. Однако, в реальности природа идет по пути простоты и элегантности своих решений, перемещение зарядов на мембране клетки может служить, как для передачи нервного возбуждения, так и для трансляции информации о том, где происходит данная передача.
Несмотря на то, что указанная работа демонстрирует, как образуются элементарные условные рефлексы в нервной системе, она приближает к пониманию того, что такое интеллект и разумная деятельность.
Существуют еще множество аспектов работы нервной системы: механизмы торможения, принципы построения эмоций, организация безусловных рефлексов и обучение, без которых невозможно построить качественную модель нервной системы. Есть понимание, на интуитивном уровне, как работает нервная система, принципы которой возможно воплотить в моделях.
Создание первой модели помогли отработать и откорректировать представление об электромагнитном взаимодействии нейронов. Понять, как происходит формирование рефлекторных дуг, как каждый отдельный нейрон понимает, каким образом ему настроить свои синапсы для получения ассоциативных связей.
На данный момент я начал разрабатывать новую версию программы, которая позволит смоделировать многие другие аспекты работы нейрона и нервной системы.
Прошу принять активное участие в обсуждении выдвинутых здесь гипотез и предположений, так как я могу относиться к своим идеям предвзято. Ваше мнение очень важно для меня.
Читайте также:
