
Никотиновые рецепторы в цнс
Ацетилхолиновые рецепторы.
Нейротрансмиттер ацетилхолин высвобождается из везикул в пресинаптических нервных терминалях и связывается как с никотиновыми, так и мускариновыми рецепторами на поверхности клетки. Эти два типа ацетихолиновых рецепторов значительно отличается как по структуре, так и по функциям. Ацетилхолиновый никотиновый рецептор является одновременно и ионным каналом, т.е. относится к рецепторам-каналоформерам, тогда как ацетилхолиновый мускариновый рецептор относится к классу серпентиновых рецепторов, осуществляющих передачу сигнала через гетеротримерные G- белки.
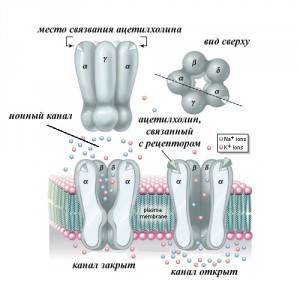
Наиболее хорошо изученным рецептором-ионным каналом является ацетилхолиновый никотиновый рецептор (рис.2) Свое название никотиновый ацетилхолиновый рецептор получил из-за его сродства к никотину. Никотин связывается непосредственно с α-субьединицей рецептора и стимулирует открывание неспецифического катионного канала, сформированного различными комбинациями α2, β, γ, δ и ε субьединиц.
Ацетилхолиновый рецептор – лиганд-зависимый ионный канал в постсинаптической мембране мышечного волокна. Это молекула массой 250 000 дальтон представляет собой пентамерный гликопротеин (рис. 3), состоящий из трансмембранных полипептидов четырех разных типов, каждый их которых кодируется отдельным геном, хотя они все во многом сходны по последовательности аминокислот, что говорит о происхождении их генов от общего гена-предшественника. Два из пяти полипептидов пентамера идентичны и образуют места связывания ацетилхолина. Две молекулы ацетилхолина присоединяются к пентамерному комплексу и вызывают конформационное изменение, приводящее к открытию канала.
ü две α-субъединицы (по 461 аминокислоте),
ü одну β-субъединицу (493 аминокислоты),
ü одну γ-субъединицу (506 аминокислот),
ü одну ε-субъединицу (522 аминокислоты).
Каждая из субъединиц образует 4 трансмембранных сегмента (4, а), а все вместе 5 субъединиц компактно собраны вокруг центральной ионной поры (рис. 4, б). Когда лиганд, в данном случае ацетилхолин (АХ), присоединяется к сайтам связывания двух альфа-субъединиц, канал открывается и одновалентные катионы двигаются по электрохимическому градиенту.
Ацетилхолин-зависимый канал, подобно потенциал-зависимому натриевому каналу, имеет несколько дискретных альтернативных конформаций и в присутствии лиганда переходит из одного состояния в другое, внезапно открываясь или закрываясь. Связав ацетилхолин и перейдя в открытое состояние, канал остается некоторое время открытым, это время варьирует случайным образом и составляет в среднем 1 мс. При длительном воздействии ацетилхолина (что в нормальных условиях случается редко) канал переходит в состояние десенсибилизации, аналогичное инактивированоому состоянию натриевых каналов. В открытой конформации канал имеет просвет, сужающийся от наружного конца диаметром 2,5 нм к внутреннему концу диаметром 0,65 нм. Заряды распределены по стенке канала таким образом, что отрицательные ионы не проходят через него, а положительные могут проникать в клетку. Ток создают в основном ионы натрия и калия, а также некоторое количество ионов кальция. Так как в отношении катионов заметной избирательности нет, вклад каждого иона в канальный ток зависит от его концентрации и электрохимической движущей силы. Открытие каналов в ацетилхолиновых рецепторах приводит к значительному притоку катионов в клетку и в результате к деполяризации мембраны.
Никотиновые холинергические рецепторы подразделяют на два типа:
Ø мышечные никотиновые рецепторы: ионный канал данного рецептора существует в дискретных открытом и закрытом состояниях. В открытом состоянии рецептор может пропускать ионы Na + та К + и, в меньшей мере, двухвалентные катионы. Проводимость ионного канала при этом является постоянной величиной. Тем не менее, время существования канала в открытом состоянии является характеристикой, которая зависит от напряжения приложенного к рецептору потенциала, при этом рецептор стабилизируется в открытом состоянии при переходе от малых значений напряжения (деполяризация мембраны) к большим (гиперполяризация). Длительно действующая аппликация ацетилхолина и других агонистов рецептора приводит к снижению его чувствительности к рецепторной молекуле и увеличению времени пребывания ионного канала в закрытом состоянии– то есть у никотинового рецептора наблюдается явление десенсетизации.
Ø нейрональные никотиновые рецепторы: данный рецептор обладает необыкновенные вольт-зависимыми свойства: общеклеточный ток, записанный в физиологическом состоянии, при наложении деполяризационных величин электрического потенциала указывает на достоверное уменьшение прохождение ионов через ионные каналы; при этом это явление регулируется концентрацией в растворе ионов Mg 2+ . Для сравнения, никотиновые рецепторы на мышечных клетках не претерпевают никаких изменений ионного тока при изменении значений мембранного электрического потенциала, а N-метил-D-аспартатний рецептор, которому также присущая высокая относительная проницаемость для ионов Са 2+ , обладает обратной картиной изменения ионных токов в ответ на смену электрического потенциала и наличие ионов магния: при повышении электрического потенциала до гиперполяризущих величин и повышении концентрации ионов Mg 2+ ионный ток через данный рецептор блокируется.
Другое важное свойство нейрональных никотиновых рецепторов – это их реакция на стимуляцию:
ü экспозиция высоких концентраций ацетилхолина приводит к очень быстрой десенсетизации ответа отдельного канала и быстрого падения электрического ответа всей клетки;
ü повторная экспозиция коротких импульсов ацетилхолина также приводит к уменьшению максимальной амплитуды рецепторного ответа. При этом энергетический подкорм клетки высокоэнергетическисодержащими молекулами (АТФ, фосфокреатин, креатин-фосфокиназа) или промежуточными продуктами их метаболизма способно предотвратить такое уменьшение.
Оба типа рецептора стимулируют токи Na + , K + и Ca 2+ за счет того, что формируют ионные каналы с электропроводимостью 50 pS (мышечный) и 15-40 pS (нейрональный), однако эти каналы существенно различаются по структуре.
Полипептидная цепь каждой из субъединиц 4 раза пронизывает мембрану. Альфа субъединица, связывающая ацетилхолин, имеет 7, а бета-субъединица – 4 изоформы.
В специализированных тканях никотиновый рецептор представлен в огромных количествах, возможно, поэтому он является наиболее изученным. Так, например, при нейромышечной передаче события развиваются следующим образом. Ацетилхолин, секретируемый из пресинаптической мембраны в синаптическую щель, взаимодействует с рецепторами постсинаптической мембраны. Это позволяет Na + входить через никотиновые каналы, вызывая, таким образом, локальную деполяризацию, которая приводит, в конечном счете, к сокращению мышцы.
В нейромышечной системе ацетилхолин действует через никотиновые холинергические рецепторы и вызывает сокращение скелетной мускулатуры. Он также передает сигнал внутри нервной системы. Эти рецепторы являются неспецифическими ионными каналами, которые проводят Na + и K + . Антагонистом для никотиновых рецепторов является тубокурарин.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Никотиновые ацетилхолиновые рецепторы относятся к одной из разновидностей ацетилхолиновых рецепторов, обеспечивающих передачу нервных импульсов в центральной и периферической нервной системе, мышечных тканях и частично, в клетках эпителия.
Для активизации этого рецептора необходим ацетилхолин, но из-за сродства этого вещества с никотином, при курении никотин замещает ацетилхолин в рецепторе и таким образом встраивается в систему передачи нервных импульсов от головного мозга на периферию.
Эти виды рецепторов впервые были изучены в начале 20 века, но только спустя несколько десятилетий, ученые доказали роль ацетилхолина в передаче нервных импульсов и мышечных сокращений, а также сродство ацетилхолина с никотином.
Что такое ацетилхолиновые рецепторы
Никотиновые ацетилхолиновые рецепторы – это один из подвидов общихацетилхолиновых рецепторов (кроме никотиновых также есть мускариновые рецепторы, получившие свое название из-за способности взаимодействия с мускарином – токсическим веществом, содержащимся в некоторых грибах, в частности – в мухоморах). Кроме физиологического сродства к ацетилхолину – веществу, вырабатывающемуся в организме человека в норме, эти рецепторы способны взаимодействовать с никотином, входящим в состав табака, из-за чего у курильщиков развивается физическая и психическая зависимость.
Ацетилхолиновые рецепторы расположены в головном мозге, периферических нервных ганглиях, гладкой и поперечнополосатой мускулатуре и в ряде других тканей и органов. Их основное предназначение – передача сигналов от нерва к мышцам или от одного нейрона к другому.
Эти рецепторы относятся к неспецифическим ионным каналам, которые проводят натрий и калий.
Строение и механизм передачи ацетилхолиновых рецепторов
Ацетилхолиновые рецепторы – это гетероолигомерный комплекс белков, состоящий из нескольких субъединиц, обозначаемых буквами греческого алфавита: α, β, γ, δ, ε (в соответствии с их молекулярной массой). В зависимости от месторасположения и выполняемых функций, структура и количество белков в разных рецепторах может меняться, неизменным остается только присутствие α белка, отвечающего за взаимодействие с агонистом – ацетилхолином или никотином.
Ацетилхолиновые рецепторы относятся к семейству мембранных белков, куда также входятрецепторы серотонина, глицина и ГАМК. Они встраиваются в мембраны клеток, образуя ионный канал, который способен пропускать положительно заряженные частицы.
Механизм действия никотиновых ацетилхолиновых рецепторов связан с их способностью открывать ионные каналы, вызывая деполяризацию мембраны клетки. При взаимодействиирецептора с ацетилхолином или никотином открывается ионный канал, по которому положительно заряженные ионы – натрия, калия или кальция, проходят внутрь клетки, меняя ее энергетический потенциал.
За счет этого передается нервный импульс или возникает сокращение мышечного волокна.
Значение ацетилхолиновых никотиновых рецепторов для жизнедеятельности человека
Ацетилхолиновые рецепторы считаются одними из важнейших в человеческом организме. Их основная задача – активизация процессов передачи импульса от нейронов к миоцитам – мышечным клеткам. Существует несколько видов таких рецепторов, расположенных в разных частях тела. Ацетилхолиновые рецепторы, расположенные в головном мозге, генерируют импульс, передающийся к периферическим нервным окончаниям, а от них – к мышечным волокнам, которые благодаря изменению потенциала мембраны, сокращаются, обеспечивая движения человеческого тела.
Но ацетилхолин задействован далеко не только в нейромышечной передаче.
Он является медиатором для:
- рецепторов, расположенных в различных отделах головного мозга;
- пресинаптических окончаний мотонейронов на клетках Реншоу (расположены в передних рогах спинного мозга и обеспечивающих стабилизацию работы мышечных клеток при постоянном мышечном напряжении, например для поддержания постоянства позы при сидении);
- симпатического отдела вегетативной нервной системы;
- синапсов мозгового вещества надпочечников;
- постганглионарных синапсов потовых желез;
Нервный импульс из центральной нервной системы передается в нервно - мышечное соединение на периферии и этот процесс активизирует высвобождение около 125 везикул ацетилхолина. Они устремляются к многочисленным ацетилхолиновым рецепторам, расположенным в мембранах мышечного волокна и представляющие собой управляемые кальциевые рецепторы. Такой рецептор представляет собой комплекс из 5 белковых субъединиц, которые образуют канал в толще мембраны клетки мышечного волокна, пронизывая его насквозь. При взаимодействии с ацетилхолиновыми молекулами канал расширяется и его диаметр достигает 0,6 нм, расстояние, достаточное для продвижения по нему положительно заряженных молекул натрия, калия и кальция. Благодаря большему отрицательному заряду, образующемуся в устье канала, ацетилхолиновые рецепторыпритягивают к себе больше количество положительно заряженных ионов, по сравнению с другими каналами, что обеспечивает более быструю деполяризацию мембраны клетки. За счет этого с внутренней стороны мембраны образуется местный положительный потенциал, который называют потенциалом концевой пластинки. Он необходим для инициации потенциала действия, который распространяется вдоль мышечного волокна и обеспечивает его синхронное сокращение.
Никотиновые ацетилхолиновые рецепторы в большой степени расположены в поперечнополосатой мускулатуре, тогда как мускариновые рецепторы обеспечивают сокращение гладкой мускулатуры внутренних органов.
Таким образом, никотиновые ацетилхолиновые рецепторы – это необходимый элемент передачи нервного импульса от коры головного мозга к скелетным мышцам и во многом именно благодаря ацетилхолину, человек способен совершать целенаправленные движения и руководить собственным телом.
Причины развития никотиновой зависимости
Никотин – растительный алкалоид, в большом количестве содержащийся в табачных листьях и при попадании в организм человека, способный вызвать физическую и психическую зависимость от курения.
Ученые надеются, что знание о природе развития никотиновой зависимости поможет курильщикам справиться с ее последствиями и легче отказаться от своей пагубной привычки.
Никотиновый ацетилхолиновых рецепторов (nACh-receptor) — подвид ацетилхолиновых рецепторов, который обеспечивает передачу нервного импульса через синапсы и активируется никотином (кроме ацетилхолина). Этот рецептор входит в группу рецепторов-ионных каналов вместе с ГАМК А -, глицинового и серотониновым 5-HT 3 рецепторами.
Этот рецептор найден в химических синапсах как в центральной, так и в периферической нервной системе, в нервно-мышечных синапсах, а также в эпителиальных клетках многих видов животных.
Структура
Высокая концентрация никотиновых ацетилхолиновых рецепторов в электрических органах некоторых скатов, в частности Torpedo californica и Torpedo marmorata, вместе с выделением α-бунгаротоксина (αBGT — полипептида с яда змеи Bungarus multicinctus, который необратимо лигандом и специфическим антагонистом ацетилхолина для никотиновых рецепторов с нервно-" мышечных синапсов), позволила биохимически очистить и детально исследовать никотиновые ацетилхолиновые рецепторы, а также идентифицировать сайт связывания ацетилхолина. Было показано, что данный рецептор является гетероолигомерним комплексом, который состоит из четырех различных белковых субъединиц, которые были названы в соответствии с их молекулярной массы (в килодальтон): α (40), β (50), γ (60), δ (65) . При естественной экспрессии в клетке сначала возникают димерные комплексы α-γ и α-δ, затем формируется триммер α-β-δ, и в конце концов, после объединения димера и тримера, в клеточную мембрану встраивается функциональный пентамер с стехиометрии α 2 βγδ.
Трансмембранно часть рецептора образует ионный канал, стенки которого сформированы сегментами М2 всех пяти субъединиц. Было доказано, что относительно небольшие пертрубации, а именно поворот на 4 ° двух агонист-связывающих субъединиц, приводят к значительному смещению сегментов М2 и открытие поры ионного канала, является условием возникновения катионного тока через рецептор.
Физиология и фармакология
Электрофизиологическая характеристика никотиновых рецепторов мускульной ткани впервые была дана благодаря внутриклеточном отводу электрических потенциалов; кроме того, никотиновый рецептор был одним из первых, на которых удалось записать электрические токи, проходящие через единичный рецепторный канал. Используя последний подход удалось показать, что ионный канал данного рецепору существует в дискретных открытом и закрытом состояниях. В открытом состоянии рецептор может пропускать ионы Na + и К + и, в меньшей степени, двухвалентного катионы; проводимость ионного канала при этом является постоянной величиной. Тем не менее, время существования канала в открытом состоянии является характеристикой, зависит от напряжения приложенного к рецептору потенциала, при этом рецептор стабилизируется в открытом состоянии при переходе от малых значений напряжения (деполяризация мембраны) до больших (гиперполяризация). Аренда аппликация ацетилхолина и других агонистов рецептора ведет к снижению его чувствительности к рецепторной молекулы и увеличение времени пребывания ионного канала в закрытом состоянии — то есть в никотинового рецептора наблюдается явление десенсетизации.
Классической характеристикой никотиновых рецепторов в нервных ганглиях и в головном мозге есть холинергические ответы на электрическое раздражение, которые блокируются дигидро-β-еритроидином; кроме того, для этих рецепторов характерно высокоафинными связывания с тритий-меченым никотином. αBGT-чувствительные рецепторы в нейронах гиппокампа характеризуются низкой чувствительностью к ацетилхолина, в отличие от αBGT-нечувствительных рецепторов. Селективным и обратимым конкурентным антагонистом αBGT чувствительных рецепторов является метилликаконитин, а некоторые производные анабезиину наносят селективный активационный влияние на эту группу рецепторов. Проводимость ионного канала αBGT чувствительных рецепторов довольно высокой (73pS) также им присуща относительно высокая проводимость ионов кальция по сравнению с ионами цезия (PCa / PCs 6.1 на фоне гиперполяризующий содержащего потенциала). Этот рецептор обнаруживает необычные вольт-зависимые свойства: обще-клеточный ток, записанный в физиологическом состоянии, при наложении деполяризации величин электрического потенциала вказуюе на достоверное уменьшение прохождения ионов через ионные каналы; при этом это явление регулируется концентрацией в растворе ионов Mg 2+. Для сравнения, никотиновые рецепторы на мышечных клетках не проявляют никакого изменения ионного тока при изменении значений мембранного электрического потенциала, а N-метил-D-аспартатных рецептор, которому также присуща высокая относительная проницаемость для ионов Са 2+ (PCa / PCs 10.1) , обнаруживает обратную картину изменения ионных токов в ответ на изменение электрического потенциала и наличие ионов магния: при повышении электрического потенциала в гиперполяризующий величин и повышении концентрации ионов Mg 2+ ионный ток через данный рецептор блокируется.
Другое важное свойство αBGT чувствительных нейрональных никотиновых рецепторов — это их реакция на стимуляцию. Экспозиция высоких концентраций ацетилхолина приводит к очень быстрой десенсетизации ответы отдельного канала и быстрого падения электрической ответы всей клетки. Повторяющихся экспозиция коротких импульсов ацетилхолина также приводит к уменьшению максимальной амплитуды рецепторного ответа. При этом энергетическая подпитка клетки високоенергомисткимы молекулами (АТФ, фосфокреатин, креатин-фосфокиназы) или промежуточными продуктами их метаболизма способно предотвратить такое уменьшению. Судя по всьго, уменьшение общей амплитуды клеточного ответа отражает прогрессирующий процесс перехода все новых порций рецепторов к некоторому стабильного десенсетизованого состояния, который может быть изменен на нормально функционирующий только с затратами энергии. Почти все аспекты функционирования αBGT чувствительных никотиновых рецепторов, включая эффективность агонистов, кооперативные эффекты, а также фракционирование по активности и десенсетизация, регулируются внеклеточного концентрацией Са 2+. Такая регуляция может быть особенно важным в случаях, когда рецепторы расположены на дендритах.
αBGT-нечувствительны нейрональные никотиновые рецепторы обладают высоким сродством к ацетилхолина, епибатидину и никотина — черта, которая определяет их селективную активацию с помощью экзогенных агонистов, что, в свою очередь, является причиной многих физиологических и поведенческих эффектов, обусловленных этим типом рецепторов. Учитывая то, что большинство (
90%) αBGT-нечувствительных рецепторов в мозгу имеют строение α4β2, этот тип рецепторов достаточно ярко проявляет много эффектов, которые вызываются низкими концентрациями агониста. С другой стороны, радиоактивно меченый агонист связывается со многими высокоафинными сайтами в нервной ткани, доводя существования других αBGT-нечувствительных рецепторов. Большое количество данных свидетельствует о том, что αBGT-нечувствительны рецепторы расположены дискретно на нескольких отдельных типах пресинаптических терминалов в специфических участках мозга, где они облегчают выделение части нейромедиаторов. По сравнению с αBGT-чувствительным сенсором, αBGT-нечувствителен рецептор имеет низкую проницаемость для Са 2+, а его активация потенцируются при высоких концентрациях Са 2+. Более того, αBGT-нечувствительны никотиновые тока делятся на чувствительные и нечувствительные к наличию в среде ионов Mg 2+.
В дополнение к селективной активации рецепторов ацетилхолин-образными агонистами, все подтипы никотиновых рецепторов активируются производными физостигмина; тем не менее, такая активация присущая только низкочастотным токам единичных рецепторов, которые не могут быть подавлены антагонистами ацетилхолина, а на общеклеточными уровне не обнаруживается. Последние исследования показывают, что активность никотиновых рецепторов, полученная с помощью ацетилхолина или других агонистов, может быть положительно модулированной производными физостигмина, которые связываются со своим специфическим сайтом на рецепторной молекуле. Достаточно возможной считается наличие эндогенного лиганда данного сайта, и наиболее вероятным кандидатом на эту роль считают 5-гирокситриптамин.
Нейроанатомия и патология
Роль периферийных никотиновых рецепторов освещается при аутоиммунных заболеваниях, при которых поражаются отдельные формы рецепторов. У пациентов, страдающих миостению, антитела к мышечным никотиновых рецепторов предотвращают нормальном моторном функционированию мышц, приводя к общему ослаблению (что первично симптомом данного заболевания).
Пузырчатка является дерматологическим поражением, которое характеризуется возникновением заполненных жидкостью волдырей, возникающих благодаря нарушению клеточной адгезии. Никотиновые рецепторы, обладающие композиция α3β2 играют важную роль в процессе клеточной адгезии в кератиноцитах, и антитела к этим рецепторам провоцируют возникновение пузырчатки.
В ЦНС дисфункция никотиновых ацетилхолиновых рецепторов в гиппокампе и коре больших полушарий приводит к возникновению болезни Альцгеймера.
Токсикология
Токсины, которые подавляют функционирование никотиновых рецепторов, найдены во многих представителей растительного и животного мира. Такой мощный лиганд, как анатоксин, который синтезируется пресноводными сине-зелеными водорослями, приводит к быстрой гибели водных организмов при цветении воды (то есть вспышки размножения этих водорослей). Некоторые растительные токсины, такие как кураре и физостигмин, поражающих никотиновые рецепторы, используются человеком уже несколько тысячелетий. Метил-ликасонитин товары шпорник (или дельфиниум — Delphinium elatum), является обратным конкурентным антагонистом с высоким сродством к αBGT чувствительных нейрональных никотиновых рецепторов. Известно также, что никотин сам по себе является веществом, которое отвечает за возникновение табачной зависимости. Данные о влиянии на функции αBGT чувствительных рецепторов ионов Pb 2+ могут, как минимум частично, объяснять негативное влияние этого металла на память и высшей нервной деятельности мозга.
В открытом состоянии никотиновые рецепторы мышечного типа блокируются большим количеством специфических лигандов — блокаторов ионного канала. К ним относятся местные анестетики (бупивакаин, кокаин, пиперокаин, лидокаин), антимускаринови соединения (атропин, скополамин), антагонисты с наркотическим эффектом (налтрексон), специфические токсины (гистрионикотоксин), антивирусные вещества и антибиотики (амантадин, квинакрин), стимуляторы (никотин ), психотропные вещества (кокаин, ССР), и фосфор-содержащие органические соединения (диизопропилфлюорофосфат, зарин, VX). Несмотря на различную химическую природу, эти агенты-блокаторы представляют собой в большинстве случаев молекулы, положительно заряженные на большей части поверхности. Учитывая то, что сайт, связывание с которым обеспечивает блокирование ионного канала, является чувствительным к заряду клеточной мембраны, действие блокаторов этого канала также електрозалежною; при гиперполяризацийних значениях заряда мембраны блокировки происходит активно.
Разновидности никотиновых рецепторов
Разновидности никотиновых рецепторов и их фармакологические свойства
Ацетилхолин (АХ) — очень важный медиатор. Деятельность холинергических нейронов центральной нервной системы (ЦНС), направляющихся от базальных структур переднего мозга к гиппокампу, обеспечивает возможность обучения и запоминания. Повреждение этих нейронов приводит к возникновению болезни Альцгеймера.
В периферической нервной системе холинергическими являются все двигательные нейроны скелетной мускулатуры, преганглионарные нейроны, иннервирующие симпатические и парасимпатические ганглии, а также постганглионарные нервные волокна, осуществляющие парасимпатическую иннервацию сердечной мышцы, гладкой мускулатуры кишечника и мочевого пузыря, а также гладкой мускулатуры глаза, ответственной за процессы аккомодации и зрения на близком расстоянии.
Ацетилхолин (АХ) синтезируется при переносе ацетильной группы с ацетилокофермента А (ацетил-КоА) на холин под действием фермента холинацетилтрансферазы. Холинацетилтрансфераза присутствует исключительно в холинергических нейронах. Холин поступает в нейрон из межклеточного пространства путем активного транспорта. Ацетил-КоА синтезируется в митохондриях, которые синтезируют холинацетилтрансферазу и в большом количестве расположены в нервных окончаниях.
После высвобождения ацетилхолина (АХ) в синаптическую щель происходит его разрушение под действием ацетилхолинестеразы (АХЭ) с образованием холина и уксусной кислоты, которые подвергаются обратному захвату и повторному использованию для синтеза новых молекул медиатора.
Этапы синтеза, распада и обратного захвата ацетилхолина (АХ) представлены на рисунке ниже.
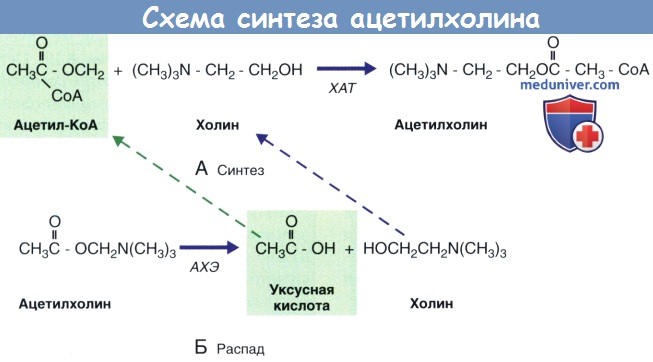
(А) Схема синтеза ацетилхолина (АХ) из ацетилкофермента А (Ацетил-КоА) и холина под действием холинацетилтрансферазы (ХАТ).
(Б) Распад молекулы ацетилхолина под действием ацетилхолинэстеразы (АХЭ).
Пунктирными стрелками обозначено повторное использование уксусной кислоты и холина.
Выделяют медиаторозависимые рецепторы ацетилхолина (АХ) и рецепторы, связанные с G-белками. Ионотропные рецепторы ацетилхолина (АХ) называют никотиновыми, поскольку первым веществом, которое вызвало их активацию, был никотин, выделенный из растения табака. Метаботропные рецепторы АХ называют мускариновыми, так как их активатором служит мускарин — вещество, выделенное из ядовитых грибов мухоморов.
2. Мускариновые рецепторы. G-белок-зависимые мускариновые рецепторы сконцентрированы (а) в височной доле головного мозга, где они участвуют в процессе формирования памяти; (б) в вегетативных ганглиях; (в) в волокнах сердечной мышцы, включая проводящие волокна; (г) в гладкой мускулатуре кишечника и мочевого пузыря; (д) в секреторных клетках потовых желез.
Выделяют пять подтипов мускариновых рецепторов — М1-М5 M1, M3- и М5-рецепторы — возбуждающие: посредством ферментных каскадов происходят активация фосфолипазы С и повышение внутриклеточного уровня Са 2+ . М2- и М4-рецепторы представляют собой тормозные ауторецепторы, уменьшающие внутриклеточный уровень цАМФ и/или увеличивающие выход К + из клетки в ходе гиперполяризации.
3. Обратный захват ацетилхолина. Продукты гидролиза ацетилхолина в синаптической щели — холин и ацетил-группа — захватываются молекулами специфических переносчиков обратно в клетку.
4. Отравление стрихнином. Стрихнин блокирует рецепторы глицина. Мучительные судороги при отравлении стрихнином обусловлены растормаживанием α-мотонейронов, вызванным нарушением тормозных влияний клеток Реншоу. Клинические проявления напоминают таковые при отравлении столбнячным токсином, который, как известно, препятствует высвобождению глицина из клеток Реншоу.
В ходе патологоанатомических исследований неизмененного мозга с использованием меченых молекул стрихнина было показано, что рецепторы глицина в большом количестве представлены на ассоциативных нейронах ядра тройничного нерва, иннервирующего жевательные мышцы, а также ядра лицевого нерва, иннервирующего мимическую мускулатуру. Именно эти две группы мышц в большей степени подвержены судорогам при отравлении.
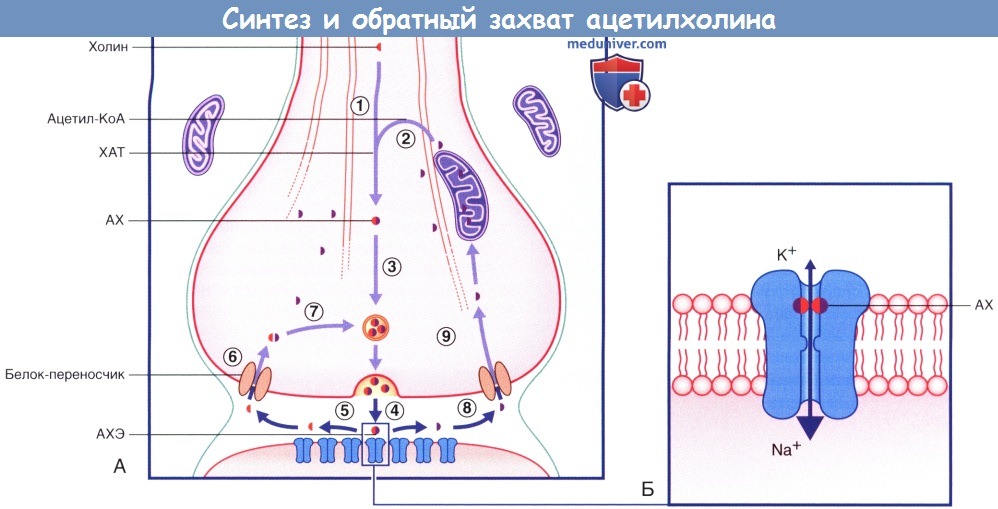
(А) Синтез и обратный захват молекул ацетилхолина (АХ) в ЦНС. На постсинаптической мембране расположены никотиновые рецепторы (н-АХ-рецепторы).
(1) Молекулы холина захватываются из межклеточной жидкости и передаются в нервное окончание.
(2) Под действием митохондриального фермента холинацетилтрансферазы (ХАТ) происходит ацетилирование холина ацетилкоферментом А (ацетил-КоА) с образованием ацетилхолина (АХ).
(3) Молекулы АХ помещаются в синаптические пузырьки.
(4) АХ высвобождается и связывается с соответствующими рецепторами.
(5) Под действием ацетилхолинэстеразы (АХЭ) происходит гидролиз молекул медиатора.
(6) Холиновые фрагменты молекул транспортируются обратно в цитозоль.
(7) Под действием трансфераз происходит синтез новых молекул ацетилхолина, которые вновь помещаются в синаптические пузырьки.
(8) Ацетатный фрагмент молекулы перемещается в цитозоль.
(9) В митохондриях происходит синтез новых молекул ацетил-КоА из уксусной кислоты.
(Б) Медиаторозависимый никотиновый рецептор. Присоединение АХ вызывает поступление большого количества ионов Na + в клетку и выход небольшого количества ионов К + из клетки.
Редактор: Искандер Милевски. Дата публикации: 12.11.2018
Читайте также:
