
Отдел нервной системы вызывают сокращение скелетных мышечных волокон
Сокращение мышц — это сложный процесс, состоящий из целого ряда этапов. Главными составляющими здесь являются миозин, актин, тропонин, тропомиозин и актомиозин, а также ионы кальция и соединения, которые обеспечивают мышцы энергией. Рассмотрим виды и механизмы мышечного сокращения. Изучим, из каких этапов они состоят и что необходимо для цикличного процесса.

Мышцы
Мышцы объединяются в группы, у которых одинаковый механизм мышечных сокращений. По этому же признаку они и разделяются на 3 вида:
- поперечно-полосатые мышцы тела;
- поперечно-полосатые мышцы предсердий и сердечных желудочков;
- гладкие мышцы органов, сосудов и кожи.
Поперечно-полосатые мышцы входят в опорно-двигательный аппарат, являясь его частью, так как помимо них сюда входят сухожилия, связки, кости. Когда реализуется механизм мышечных сокращений, выполняются следующие задачи и функции:
- тело передвигается;
- части тела перемещаются друг относительно друга;
- тело поддерживается в пространстве;
- вырабатывается тепло;
- кора активируется посредством афферентации с рецептивных мышечных полей.
Из гладких мышц состоит:
- двигательный аппарат внутренних органов, в который входят бронхиальное дерево, легкие и пищеварительная трубка;
- лимфатическая и кровеносная системы;
- система мочеполовых органов.
Физиологические свойства

Как и у всех позвоночных животных, в человеческом организме выделяют три самых важных свойства волокон скелетных мышц:
- сократимость — сокращение и изменение напряжения при возбуждении;
- проводимость — движение потенциала по всему волокну;
- возбудимость — реагирование на раздражитель посредством изменения мембранного потенциала и ионной проницаемости.
Мышцы возбуждаются и начинают сокращаться от нервных импульсов, идущих от центров. Но в искусственных условиях используют электростимуляцию. Мышца тогда может раздражаться напрямую (прямое раздражение) или через нерв, иннервирующий мышцу (непрямое раздражение).
Виды сокращений
Механизм мышечных сокращений подразумевает преобразование химической энергии в механическую работу. Этот процесс можно измерить при эксперименте с лягушкой: ее икроножную мышцу нагружают небольшим весом, а затем раздражают легкими электроимпульсами. Сокращение, при котором мышца становится короче, называется изотоническим. При изометрическом сокращении укорачивания не происходит. Сухожилия не позволяют при развитии мышцей силы укорачиваться. Еще один ауксотонический механизм мышечных сокращений предполагает условия интенсивных нагрузок, когда мышца укорачивается минимальным образом, а сила развивается максимальная.
Структура и иннервация скелетных мышц
В поперечно-полосатые скелетные мышцы входит множество волокон, находящихся в соединительной ткани и крепящихся к сухожилиям. В одних мышцах волокна расположены параллельно длинной оси, а в других они имеют косой вид, прикрепляясь к центральному тяжу сухожильному и к перистому типу.

Главная особенность волокна заключается в саркоплазме массы тонких нитей — миофибрилл. В них входят светлые и темные участки, чередующиеся друг с другом, а у соседних поперечно-полосатые волокна находятся на одном уровне — на поперечном сечении. Благодаря этому получается поперечная полосатость по всему волокну мышц.
Саркомером является комплекс из темного и двух светлых дисков, и он отграничивается Z-образными линиями. Саркомеры — это сократительный аппарат мышцы. Получается, что сократительное мышечное волокно состоит из:
- сократительного аппарата (системы миофибрилл);
- трофического аппарата с митохондриями, комплексом Гольджи и слабой эндоплазматической сетью;
- мембранного аппарата;
- опорного аппарата;
- нервного аппарата.
Мышечное волокно разделяется на 5 частей со своими структурами и функциями и является целостной частью ткани мышц.
Иннервация
Иннервация может быть одиночной и множественной. В первом случае она реализуется компактными моторными окончаниями. Обычно это характерно для крупных мотонейронов. Мышечные волокна (называющиеся в этом случае физическими, или быстрыми) генерируют ПД (потенциалы действий), которые распространяются на них.
Множественная иннервация встречается, к примеру, во внешних глазных мышцах. Здесь не генерируется потенциал действия, так как в мембране нет электровозбудимых натриевых каналов. В них распространяется деполяризация по всему волокну из синаптических окончаний. Это необходимо для того, чтобы привести в действие механизм мышечного сокращения. Процесс здесь происходит не так быстро, как в первом случае. Поэтому его называют медленным.
Структура миофибрилл
Исследования мышечного волокна сегодня проводятся на основе рентгеноструктурного анализа, электронной микроскопии, а также гистохимическими методами.
Рассчитано, что в каждую миофибриллу, диаметр которой составляет 1 мкм, входит примерно 2500 протофибрилл, то есть удлиненных полимеризованных молекул белков (актина и миозина). Актиновые протофибриллы в два раза тоньше миозиновых. В покое эти мышцы находятся так, что актиновые нити кончиками проникают в промежутки между миозиновыми протофибриллами.
Узкая светлая полоса в диске А свободна от актиновых нитей. А мембрана Z скрепляет их.
На миозиновых нитях есть поперечные выступы длиной до 20 нм, в головках которых находится порядка 150 молекул миозина. Они отходят биополярно, и каждая головка соединяет миозиновую с актиновой нитью. Когда происходит усилие актиновых центров на нитях миозина, актиновая нить приближается к центру саркомера. В конце миозиновые нити доходят до линии Z. Тогда они занимают собой весь саркомер, а актиновые находятся между ними. При этом длина диска I сокращается, а в конце он исчезает полностью, вместе с чем линия Z становится толще.
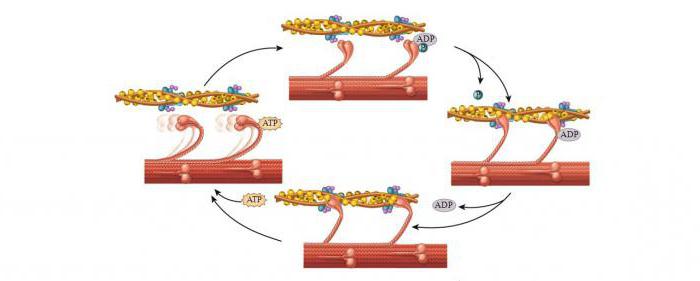
Механизм мышечного сокращения волокна
Главным в теории является то, что не нити (миозиновые и актиновые) укорачиваются. Длина их остается неизменной и при растяжении мышц. Но пучки тонких нитей, проскальзывая, выходят между толстыми нитями, уменьшается степень их перекрытия, таким образом происходит сокращение.
Молекулярный механизм мышечного сокращения посредством скольжения актиновых нитей заключается в следующем. Миозиновые головки соединяют протофибриллу с актиновой. При их наклонах происходит скольжение, двигающее актиновую нить к центру саркомера. За счет биполярной организации миозиновых молекул на обеих сторонах нитей создаются условия для скольжения актиновых нитей в разные стороны.
При расслаблении мышц миозиновая головка отходит от актиновых нитей. Благодаря легкому скольжению расслабленные мышцы растяжению сопротивляются гораздо меньше. Поэтому они пассивно удлиняются.
Этапы сокращения
Механизм мышечного сокращения кратко можно подразделить на следующие этапы:
- Мышечное волокно стимулируется, когда потенциал действия поступает от мотонейронов из синапсов.
- Потенциал действия создается на мембране мышечного волокна, а затем распространяется к миофибриллам.
- Совершается электромеханическое сопряжение, представляющее собой преобразование электрического ПД в механическое скольжение. В этом обязательно участвуют ионы кальция.
Ионы кальция

Для лучшего понимания процесса активации волокна ионами кальция удобно рассмотреть структуру актиновой нити. Длина ее составляет порядка 1 мкм, толщина — от 5 до 7 нм. Это пара закрученных ниток, которые напоминают мономер актина. Примерно через каждые 40 нм здесь находятся сферические тропониновые молекулы, а между цепями — тропомиозиновые.
Когда ионы кальция отсутствуют, то есть миофибриллы расслабляются, длинные тропомиозиновые молекулы блокируют крепление актиновых цепей и мостиков миозина. Но при активизации ионов кальция тропомиозиновые молекулы опускаются глубже, и участки открываются.
Тогда миозиновые мостики прикрепляются к актиновым нитям, а АТФ расщепляется, и сила мышц развивается. Это становится возможным за счет воздействия кальция на тропонин. При этом молекула последнего деформируется, проталкивая тем самым тропомиозин.
Когда мышца расслаблена, в ней на 1 грамм сырого веса содержится больше 1 мкмоль кальция. Соли кальция изолированы и находятся в особых хранилищах. В противном случае мышцы бы все время сокращались.
Хранение кальция происходит следующим образом. На разных участках мембраны клетки мышцы внутри волокна имеются трубки, через которые происходит соединение со средой вне клеток. Это система поперечных трубочек. А перпендикулярно ей находится система продольных, на концах которых — пузырьки (терминальные цистерны), расположенные в непосредственной близости к мембранам поперечной системы. Вместе получается триада. Именно в пузырьках хранится кальций.
Так ПД распространяется внутрь клетки, и происходит электромеханическое сопряжение. Возбуждение проникает в волокно, переходит в продольную систему, высвобождает кальций. Таким образом осуществляется механизм сокращения мышечного волокна.

3 процесса с АТФ
При взаимодействии обеих нитей при наличии ионов кальция немалая роль отводится АТФ. Когда реализуется механизм мышечного сокращения скелетной мышцы, энергия АТФ применяется для:
- работы насоса натрия и калия, который поддерживает постоянную концентрацию ионов;
- этих веществ по разные стороны мембраны;
- скольжения нитей, укорачивающих миофибриллы;
- работы насоса кальция, действующего для расслабления.
АТФ находится в клеточной мембране, нитях миозина и мембранах ретикулума саркоплазматического. Фермент расщепляется и утилизируется миозином.
Потребление АТФ
Известно, что миозиновые головки взаимодействуют с актином и содержат элементы для расщепления АТФ. Последняя активизируется актином и миозином при наличии ионов магния. Поэтому расщепление фермента происходит при прикреплении миозиновой головки к актину. При этом чем больше поперечных мостиков, тем скорость расщепления будет выше.
Механизм АТФ
После завершения движения молекула АФТ обеспечивает энергией для разделения участвующих в реакции миозина и актина. Миозиновые головки разделяются, АТФ расщепляется до фосфата и АДФ. В конце подсоединяется новая АТФ-молекула, и цикл возобновляется. Таковым является механизм мышечного сокращения и расслабления на молекулярном уровне.
Активность поперечных мостиков будет продолжаться лишь до тех пор, пока происходит гидролиз АТФ. При блокировке фермента мостики не станут снова прикрепляться.
С наступлением смерти организма уровень АТФ в клетках падает, и мостики остаются устойчиво прикрепленными к актиновой нити. Так происходит стадия трупного окоченения.

Ресинтез АТФ
Ресинтез возможно реализовать двумя путями.
Посредством ферментативного переноса от креатинфосфата фосфатной группы на АДФ. Так как запасов в клетке креатинфосфата намного больше АТФ, ресинтез реализуется очень быстро. В то же время посредством окисления пировиноградной и молочной кислот ресинтез будет осуществляться медленно.
АТФ и КФ могут исчезнуть полностью, если ресинтез будет нарушен ядами. Тогда и кальциевый насос прекратит работу, вследствие чего мышца необратимо сократится (то есть настанет контрактура). Таким образом, нарушится механизм мышечного сокращения.
Физиология процесса
Подытоживая вышесказанное, отметим, что сокращение волокна мышцы состоит в укорочении миофибрилл в каждом из саркомеров. Нити миозина (толстые) и актина (тонкие) связаны концами в расслабленном состоянии. Но они начинают скользящие движения друг навстречу к другу, когда реализуется механизм мышечного сокращения. Физиология (кратко) объясняет процесс, когда под влиянием миозина выделяется необходимая энергия для преобразования АТФ в АДФ. При этом активность миозина будет реализована лишь при достаточном содержании ионов кальция, накапливающихся в саркоплазматической сети.
Теория по нормальной физиологии на тему: Синапс. Физиология мышечных волокон. В данной статье рассматривается механизм синаптической передачи, НМС…
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Синапс — это специфическое место контакта двух возбудимых систем (клеток) для передачи возбуждения.
По способу передачи сигналов:
- механические,
- химические,
- электрические.
По виду медиатора: холинэргические и др.
Нервно-мышечный синапс (НМС) — химический, передача с помощью медиатора ацетилхолина.
Синонимы к слову НМС:
- Нервно-мышечное соединение;
- Моторная концевая пластинка.

Аксоны нервных клеток на своих окончаниях теряют миелиновую оболочку , ветвятся, и концевые веточки аксона утолщаются. Это пресинаптическая терминаль или бляшка или пуговка , которая погружается в углубление на поверхности мышечного волокна.
Покрывающая концевую веточку аксона поверхностная мембрана называется пресинаптической мембраной , т.е. это мембрана, покрывающая поверхность синаптической бляшки (терминали аксона).
Мембрана, покрывающая мышечное волокно в области синапса, называется постсинаптической мембраной , или концевой пластинкой. Она имеет извитую структуру, образуя многочисленные складки, уходящие вглубь мышечного волокна, за счет чего увеличивается площадь контакта.

На постсинаптической мембране находятся белковые структуры — рецепторы , способные связывать медиатор . В одном синапсе количество рецепторов достигает 10-20 млн.
Между пре- и постсинаптическими мембранами находится синаптическая щель , размеры ее в среднем 50 нм, она открывается в межклеточное пространство и заполнена межклеточной жидкостью .
В синаптической щели находится мукополисахаридное плотное вещество в виде полосок, мостиков и содержится фермент ацетилхолинэстераза .
В пресинаптической терминали находится большое количество пузырьков или везикул , заполненных медиатором — химическим веществом посредником, осуществляющим передачу возбуждения.
В нервно-мышечном синапсе медиатор — ацетилхолин (АХ).

АХ синтезируется из холина и уксусной кислоты (ацетил-коэнзима А) с помощью фермента холинэстеразы. Эти вещества перемещаются из тела нейрона по аксону к пресинаптической мембране. Здесь в пузырьках происходит окончательное образование АХ.
3 фракции медиатора:
- Первая фракция — доступная — располагается рядом с пресинаптической мембраной.
- Вторая фракция — депонированная — располагается над первой фракцией.
- Третья фракция — диффузно рассеянная — наиболее удаленная от пресинаптической мембраны.
Механизм синаптической передачи



Ионы Ca вызывают образование специального белкового комплекса , который включает в себя везикулу и структуры, расположенные непосредственно около пресинаптической мембраны.
Они связаны между собой так называемыми белками экзоцитоза.
Часть белков расположена на везикулах (синапсин, синаптотагмин, синаптобревин), а часть — на пресинаптической мембране (синтаксин, синапсо-ассоциированный белок). Данный комплекс получил название секретосома .



Излитию содержимого пузырька в щель способствует белок синаптопорин , формирующий канал, по которому идет выброс медиатора.
Квант медиатора — количество молекул, содержащихся в одной везикуле.
На 1 ПД выбрасывается 100 квантов АХ.





На постсинаптической мембране возникает потенциал концевой пластинки (ПКП). Он является аналогом локального ответа (ЛО).
Потенциал действия на постсинаптической мембране не возникает ! Он формируется на соседней мембране мышечного волокна.
- связывание с рецептором,
- разрушение ферментов (ацетилхолинэстеразой),
- обратное поглощение в пресинаптическую мембрану,
- вымывание из щели и фагоцитоз.
События в синапсе :
- ПД приходит к терминали аксона;
- Он деполяризует пресинаптическую мембрану;
- Ca2+ входит в терминаль, что приводит к выделению АХ;
- В синаптическую щель выделяется медиатор АХ;
- Он диффундирует в щель и связывается с рецепторами постсинаптической мембраны;
- Меняется проницаемость постсинаптической мембраны для ионов Na+;
- Ионы Na+ проникают в постсинаптическую мембрану и уменьшают ее заряд — возникает потенциал концевой пластинки (ПКП) .
На самой постсинаптической мембране ПД возникнуть не может, так как здесь отсутствуют потенциалзависимые каналы, они являются хемозависимыми!
- ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и его распространению по мышечному волокну (около 5 м/с).
Достигнув пороговой величины, то есть КУД, ПКП возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна за счет местных круговых токов.

Особенности проведения возбуждения в нервно-мышечном синапсе
Одностороннее проведение возбуждения — только в направлении от пресинаптического окончания к постсинаптической мембране.
Суммация возбуждения соседних постсинаптических мембран.
Синаптическая задержка — замедление в проведении импульса от нейрона к мышце составляет 0,5-1 мс. Это время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, взаимодействие с рецептором, формирование ПКП, их суммацию.
Низкая лабильность — она составляет 100-150 имп/с для сигнала, что в 5-6 раз ниже лабильности нервного волокна.
Чувствительность к действию лекарственных веществ, ядов, БАВ, выполняющих роль медиатора.
Утомляемость химических синапсов — выражается в ухудшении проводимости вплоть до блокады в синапсе при длительном функционировании синапса. Главная причина утомляемости — исчерпание запасов медиатора в пресинаптическом окончании.
Законы проведения возбуждения по нервам:
- Закон функциональной целостности нерва.
- Закон изолированного проведения возбуждения.
- Закон двустороннего проведения возбуждения.
В зависимости от скорости проведения возбуждения нервные волокна подразделяются на 3 группы: A, B, C. В группе A выделяют 4 подгруппы: альфа, бетта, гамма и сигма.
Физиология мышечных волокон
- скелетная (40-50% массы тела),
- сердечная (менее 1%),
- гладкая (8-9%).
Физиологические свойства скелетных мышц:
- Возбудимость — способность отвечать на действие раздражителя возбуждением.
- Проводимость — способность проводить возбуждение из места его возникновения к другим участкам мышцы.
- Лабильность — способность мышцы сокращаться в соответствии с частотой действия раздражителя (200-300 Гц для скелетной мышцы).
- Сократимость — для мышцы является специфическим свойством — это способность мышцы изменять длину или напряжение в ответ на действие раздражителя.
Физические свойства скелетных мышц:
- Растяжимость — способность мышцы изменять длину под действием растягивающей силы.
- Эластичность — способность мышцы восстанавливать первоначальную длину или форму после прекращения действия растягивающей силы.
- Силы мышц — способность мышцы поднять максимальный груз.
- Способность мышцы совершать работу.
Режимы сокращения:
- Изотонический,
- Изометрический,
- Ауксотонический.
Изотонический режим — сокращение мышцы происходит с изменением ее длины без изменения напряжения (тонуса) (напр.: сокращение мышц языка).
Изометрический режим — длина постоянная, увеличивается степень мышечного напряжения (тонуса) (напр.: при поднятии непосильного груза).
Ауксотонический режим — одновременно изменяется длина и напряжение мышцы (характерен для обычных двигательных актов).
Механизм сокращения поперечно-полосатых мышц
Любая скелетная мышца состоит из мышечных волокон, которые, в свою очередь, состоят из множества тонких нитей — миофибрилл , расположенных продольно. Каждая миофибрилла состоит из протофибрилл — нитей сократительных белков: миозина (миозиновая протофибрилла), актина (актиновая протофибрилла).
Кроме сократительных белков в миофибрилле имеются два регуляторных белка: тропомиозин и тропонин .
Миозиновые волокна соединены в толстый пучок, от которого в торону актиновых нитей отходят поперечные мостики. У каждого мостика выделяют шейку и головку.
Нить актина располагается в виде 2 скрученных ниток бус. На ней имеются актиновые центры.

Тропомиозин в виде спиралей оплетает поверхность актина, закрывая в покое ее центры. Одна молекула тропомиозина контактирует с 7 молекулами актина.
Тропонин образует утолщение на конце каждой нити тропомиозина.
Под влиянием возникшего в мышечном волокне ПД из саркоплазматического ретикулума (СПР — депо Ca2+) высвобождаются ионы Ca. Кальций связывается с тропонином, который смещает тропомиозиновый стержень, что приводит к открытию актиновых центров.

В результате, к актиновым центрам присоединяются головки поперечных миозиновых мостиков.
Процесс расслабления происходит в обратной последовательности с использованием энергии АТФ за счет функционирования кальциевого насоса.
При отсутствии повторного импульса ионы Ca не поступают из СПР. В результате отсутствия Ca-тропонинового комплекса, тропомиозин возвращается на свое прежнее место, блокируя актиновые центры актина. Актиновые протофибриллы легко скользят в обратном направлении благодаря эластичности мышцы, и мышца удлиняется (расслабляется).
Гладкие мышцы
Гладкие мышцы — это мышцы, формирующие слой стенок полых внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток без поперечной исчерченности за счет хаотичного расположения миофибрилл.
Особенности гладких мышц:
- Иннервируются волокнами вегетативной нервной системы (ВНС);
- Обладают низкой возбудимостью:
- Обладают низкой величиной МП (мембранного потенциала) — -50 — -60 мВ из-за более высокой проницаемости для ионов Na+
- ПД (потенциал действия) отличается меньшей амплитудой и большей длительностью. Он формируется в основном за счет ионов Ca2+
- Медленная проводимость:
Клетки в гладких мышцах функционально связаны между собой посредством щелевидных контактов — нексусов, которые имеют низкое электрическое сопротивление. За счет этих контактов ПД распространяется с одного мышечного волокна на другое, охватывая большие мышечные пласты, и в реакцию вовлекается вся мышца.
Гладкие мышцы способны осуществлять относительно медленные ритмические и длительные тонические сокращения.
Медленные ритмические сокращения обеспечивают перемещение содержимого органа из одной области в другую.
Длительные тонические сокращения, особенно сфинктеров полых органов, препятствуют выходу из них содержимого.
Это способность сохранять приданную им при растяжении или деформации форму. Благодаря пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии.
Особенность гладких мышц, отличающая их от скелетных. Благодаря автоматии гладкие мышцы могут сокращаться в условиях отсутствия иннервации . Важную роль в этом играет растяжение.
Растяжение является адекватным раздражителем для гладкой мускулатуры. Сильное и резкое растяжение гладких мышц вызывает их сокращение.
Сравнительная характеристика скелетных и гладких мышц:
Типы мышечных тканей
Скелетные мышцы являются единственной мышечной тканью в организме человека, которая управляется сознательно. Каждое физическое действие, которое человек сознательно выполняет (например: разговор, ходьба или письмо) требует движения скелетных мышц. Скелетные могут сжиматься, чтобы перемещать части тела ближе к кости, к которой мышца прикрепляется. Большинство скелетных мышц прикреплены к двум костям через суставы, так что они служат для перемещения частей этих костей ближе друг к другу.
Каркасные (скелетные) мышечные клетки образуются, когда множество мелких клеток — предшественников скомковываются вместе, чтобы сформировать длинные, прямые, многоядерные волокна. Исчерчены каркасные мышцы так же, как и сердечная, поэтому они очень сильны. Скелетная мышца получает свое название от того, что она всегда подключаются к скелету, по крайней мере, в одном месте.
Анатомия скелетных мышц
Большинство скелетных прикреплены к двум костям через сухожилия. Сухожилия — жесткие полосы плотной регулярной соединительной ткани; сильные коллагеновые волокна прочно прикрепляют мышцы к костям. Сухожилия находятся в крайнем напряжении, когда они тянутся, так что они очень сильно вплетены в покрытия мышц и костей.
Мышцы двигаются за счет сокращения их длины, натягивания сухожилий и перемещения костей ближе друг к другу. Одна из костей втягивается по направлению к другой кости, которая остается неподвижной. Место на движущейся кости, которая соединяется с мышцей через сухожилия называется вставкой. Мышцы живота находятся между сухожилиями, что позволяет делать фактическое сокращение.
Названия скелетных мышц
Их названия происходят на основе множества различных факторов, в том числе местонахождения, происхождения и вставки, количества, формы, размера, направления и функции.
Много мышц получают имена от анатомической области. Брюшная и прямая, поперечная брюшная, например, находятся в брюшной полости. Другие, как и передняя большеберцовая, названы из-за части кости (передняя часть голени), к которой они присоединены. Другие мышцы используют симбиоз двух видов названий, как плечелучевая, которая названа в честь области нахождения.
Некоторые мышцы названы на основе их подключения к стационарной и движущейся кости. Эти мышцы становится очень легко определить, когда вы знаете имена костей, к которым они присоединены.
Некоторые подключаются к более чем 1 кости или более чем в одном месте и имеют более чем один источник. Мышца сразу с двумя происхождения называется бицепсом, а с тремя происхождения — трицепсной. И, наконец, мышца с четырьмя происхождениями называется четырехглавой.
Также важно классифицировать мышцы по форме. Например, дельтовидные имеют дельта — или треугольную форму. Зубчатые имеют зубчатую или пилообразный форму. Ромбовидные — обладают формой ромба.
Размер может быть использован, чтобы различать два типа мышц, найденных в одном и том же регионе. Область ягодичной части содержит три мышцы, дифференцированные по размеру: ягодичная большая, ягодичная средняя и малая. И, наконец, направления мышечных волокон могут быть использованы для их идентификации. В брюшине существует несколько широких и плоских. Мышцы с волокнами, расположенными вверх и вниз — являются прямыми, работающие в поперечном направлении (слева направо) — поперечные, а работающие под углом, являются косыми.
Функции мышечной ткани человека
Мышцы иногда классифицируют по типу функции, которую они выполняют. Большинство мышц предплечья именуются в зависимости от их функций, потому что они расположены в том же регионе и имеют одинаковые формы и размеры. Например, сгибатели предплечья сгибают запястья и пальцы.
Супинатор — это мышца, которая поднимает запястье ладонью вверх. В ноге есть такие, которые называются аддукторами, чья роль заключается в стягивании ног.
Чаще всего они работают в группах, чтобы произвести точные движения. Мышца, которая производит какое — либо конкретное движение тела известна как агонист или тягач. Агонисты всегда парны с антагонистами, которые производят противоположный эффект на одних и тех же костях. Например, двуглавая мышцы плеча сгибает руку в локте. В качестве антагониста для этого движения — трехглавая плеча — расширяет руку в локте. Когда трицепсы расширяют руку, бицепс будет считаться антагонистом.
В дополнение к агонист / антагонист классификации, другие мышцы работают, чтобы поддержать движение агониста.
Синергистами являются мышцы, которые помогают стабилизировать движение и уменьшить лишние движения. Они обычно находятся в областях вблизи агониста и часто подключаются к той же кости. Если вы поднимаете что-то тяжелое, они помогают держать тело в вертикальном положении неподвижно, так что вы поддерживаете свой баланс во время подъема.
Гистология скелетной мускулатуры
Скелетные мышечные волокна значительно отличаются от других тканей организма из — за их узкоспециализированных функций. Многие из органелл, которые составляют мышечные волокна являются уникальными для данного типа клетки.
Сарколемма является клеточной мембраной мышечных волокон. Сарколемма выступает в качестве проводника для электрохимических сигналов, которые стимулируют мышечные клетки. Подключенные к сарколемме поперечные трубочки (Т-трубочки) помогают переносить электрохимические сигналы в середину мышечного волокна. Саркоплазматический ретикулум служит в качестве хранилища для ионов кальция (Са2 +), которые имеют жизненно важное значение для сокращения мышц.
Митохондрии, движущая сила клетки, в изобилии находятся в мышечных клетках, чтобы обеспечивать энергией в виде АТФ активные мышцы. Большая часть структуры мышечного волокна выполнена из миофибрилл, которые являются сократительными структурами клетки. Миофибриллы составлены из многих белковых волокон, расположенных в повторяющихся субъединицах, называемых саркомерами. Саркомера является функциональной единицей мышечных волокон.
Саркомеры изготавливаются из двух типов белковых волокон: толстых нитей и тонких нитей.
Толстые нити состоят из множества соединенных звеньев белка миозина. Миозин является белком, который вызывает мышцы сокращаться.
Тонкие нити состоят из трех белков:
Актин.
Актин образует спиральную структуру, которая составляет большую часть массы тонкой нити.
Тропомиозин.
Тропомиозин — длинный волокнистый белок, который оборачивается вокруг актина и охватывает миозин, связывая с актином.
Тропонин.
Белок, связывающийся очень плотно с тропомиозином во время мышечного сокращения.
Функции мышечной ткани
Основной функцией мышечной системы является движение. Мышцы являются единственной тканью в организме, что имеет возможность перемещать другие части тела.
Связанная с функцией движения является вторая функция мускульной системы: поддержание позы и положения тела. Мышцы зачастую держат тело неподвижно или в определенном положении, а не вызывают движение. Мышцы, отвечающие за положение тела имеют наивысшую выносливость — они выполняют свои функции в течение всего дня, не становясь усталыми.
Еще одна функция, связанная с движением является движение веществ внутри тела. Сердечные и висцеральные мышцы, в первую очередь, ответственны за транспортировку веществ, таких как кровь или питательные вещества из одной части тела в другую.
Последняя функция мышечной ткани является генерация тепла . В результате высокой скорости метаболизма сокращающейся мышцы, наша мышечная система производит большое количество отработанного тепла. Многие небольшие сокращения мышц в организме производят наше естественное тепло тела. Когда мы прилагаем усилия больше, чем обычно, дополнительные сокращения мышц приводят к повышению температуры тела и в конечном итоге к потливости.
Скелетная мускулатура в роли рычага
Мышцы скелетной системы работают вместе с костями и суставами образуя рычажные системы. Они действуют как передатчики усилия, а кость выступает в качестве опоры; при движении мышцы и кости, объект перемещается.
Есть три класса рычагов, но подавляющее большинство рычагов в теле — рычаги третьего класса. Рычаг третьего класса представляет собой систему, в которой точка опоры находится на конце рычага. В организме, рычаги третьего класса, служат для увеличения расстояния для сокращения мышцы.
Двигательные единицы мышц
Нервные клетки, называемые моторными нейронами, управляют скелетными мышцами. Каждый двигательный нейрон контролирует несколько мышечных клеток в группе. Когда двигательный нейрон получает сигнал от мозга, он стимулирует все клетки мышц в то же время.
Размер двигательных единиц изменяется по всему телу, в зависимости от функции. Мышцы, которые выполняют тонкие движения — как мышцы глаз или пальцев, имеют очень много нейронов для повышения точности контроля мозга над этими структурами. Мышцы, которые требуют много сил, чтобы выполнять свои функции, как ноги или руки — имеют много мышечных клеток и меньше нейронов в каждом блоке.
Когда положительные ионы достигают саркоплазматического ретикулума, ионы Са2 + высвобождаются и протекают в миофибриллы. Ионы Са2 + связываются с тропонином, что вызывает молекулу тропонина изменять форму и переместить близлежащие молекулы тропомиозина. Тропомиозин отодвигается от миозина и связывается с молекулой актина, что позволяет актину и миозину связываться друг с другом.
Типы мышечных сокращений
Силой сжатия мышц можно управлять двумя факторами: количеством двигательных единиц (нейронов), участвующих в сокращении и количеством импульсов от нервной системы. Один нервный импульс моторного нейрона вызовет краткое напряжение группы мышц, а затем заставит расслабиться. Если двигательный нейрон обеспечивает несколько сигналов в течение короткого периода времени, то сила и продолжительность сжатия увеличивается. Если двигательный нейрон обеспечивает много нервных импульсов в быстрой последовательности, мышца может войти в состояние полного и прочного сокращения. Мышца останется в сжатом положении, пока скорость сигнала нерва не замедлится или до тех пор, пока мышца станет слишком усталой, чтобы поддерживать напряжение.
Не все сокращения мышц производят движение. Изометрическое сокращение — легкие схватки, которые увеличивают напряжение в мышцах, не оказывая достаточной силы, чтобы переместить часть тела. Когда тело напряжено из-за стресса, мышцы выполняют изометрическое сокращение. Поддержание позы является также результатом изометрических сокращений. Сужения мышц, что действительно производит движение является изотоническими сокращениями. Изотонические сокращения необходимы для наращивания мышечной массы за счет подъема веса.
Мышечный тонус является естественным состоянием, в котором скелетные мышцы остаются во всё время. Мышечный тонус обеспечивает легкое натяжение мышц, чтобы предотвратить повреждение мышц и суставов от резких движений, а также помогает поддерживать осанку тела. Все не повреждённые мышцы поддерживают некоторое количество мышечного тонуса во всё время.
Функциональные типы скелетных мышечных волокон
Cкелетные мышечные волокона, можно разделить на два типа в зависимости от того, как они производят и используют энергию:
I тип — волокна с очень медленным и осторожным сокращением. Они очень устойчивы к усталости, потому что используют аэробное дыхание для производства энергии из сахара. Находятся I типа волокона в мышцах по всему телу для выносливости и осанки, рядом с позвоночником и в регионах шеи.
Волокна типа II разбиты на две подгруппы: II типа А и типа II B.
Тип II волокна А быстрее и сильнее, чем I типа волокона, но не имеют столько же выносливости. Типа II A волокна находятся по всему телу, но особенно в ногах,где они работают, чтобы поддерживать ваше тело на протяжении долгого времени для ходьбы и стояния.
Тип II B — волокна еще быстрее и сильнее, чем II типа А, но еще меньше выносливые. Тип II B волокна немного светлее, чем тип I и тип II А из-за их отсутствия миоглобина — кислородного пигмента. Находятся волокна типа II B по всему телу, но особенно в верхней части, где они дают скорость и силу рукам и груди за счет выносливости.
Мышечный метаболизм и усталость
Мышцы получают энергию из различных источников, в зависимости от ситуации, в которой мышца работает. Мышцы способны использовать аэробное дыхание, когда необходимо произвести от низкого до умеренного уровня силы упражнения. Аэробное дыхание требует кислорода, чтобы произвести около 36-38 молекул АТФ из молекулы глюкозы. Аэробные дыхания является очень эффективным и может продолжаться до тех пор, пока мышца получает достаточное количество кислорода и глюкозы. Когда мы используем мышцы, чтобы произвести высокий уровень силы, они становятся настолько плотными, что находящийся кислород в крови не может войти в мышцу. Это условие приводит к тому, что мышцы используют для выработки энергии брожение молочной кислоты (форма анаэробного дыхания). Анаэробное дыхание менее эффективно аэробного дыхания — только 2 АТФ производится из каждой молекулы глюкозы.
Для того, чтобы мышцы работали в течение более длительного периода времени, мышечные волокна содержат несколько важных энергетических молекул. Миоглобин, красный пигмент содержащийся в мышцах, содержит железо и сохраняет кислород в манере, подобной гемоглобину крови. Кислород из миоглобина позволяет мышцам продолжать аэробное дыхание в отсутствии кислорода. Другой химикат, который помогает мышцам работать — креатинфосфат. Мышцы используют энергию в виде АТФ, происходит превращение АТФ в АДФ, чтобы выпустить свою энергию. Креатинфосфат жертвует свою фосфатную группу АДФ, чтобы включить её в АТФ, с тем, чтобы обеспечить дополнительную энергию для мышц. Наконец, мышечные волокна содержат энергию аккумулирующих гликогенов, больших макромолекул, изготовленных из множества связанной между собой глюкозы. Активные мышцы отщепляют глюкозу от молекул гликогена, чтобы обеспечить внутренний запас топлива.
Мышечная усталость
Когда мышцы исчерпали энергию во время аэробного или анаэробного дыхания, то быстро утомляются и теряют способность сокращаться. Это состояние известно как мышечная усталость. Утомление мышц не говорит о содержании очень малого количества или отсутствия кислорода, глюкозы или АТФ, но вместо этого имеет много продуктов — отходов дыхания, таких как молочная кислота и АДФ. Тело должно принимать дополнительное количество кислорода после физической нагрузки, чтобы заменить кислород, который находился в миоглобине мышечных волокон, а также для питания аэробного дыхания, которое обеспечивает поставки энергии внутри клетки. Восстановление потребления кислорода (кислородное голодание) — это восприятие дополнительного кислорода, который организм должен принять, чтобы восстановить мышечные клетки, их привести в состояние покоя. Это объясняет, почему появляется одышка в течение нескольких минут после напряженной деятельности — ваше тело пытается восстановить себя в нормальное состояние.
Читайте также:
