
Полярный закон возбуждения нерва
(изменения мембранного потенциала при действии на возбудимые ткани постоянного электрического тока).
Постоянный ток проявляет свое раздражающее действие только в момент замыкания и размыкания цепи.
При замыкании цепи постоянного тока возбуждение возникает под катодом; при размыкании по анодом.
Изменение возбудимости под катодом.
При замыкании цепи постоянного тока под катодом (действуют допороговым, но продолжительным раздражителем) на мембране возникает стойкая длительная деполяризация, которая не связана с изменением ионной проницаемости мембраны, а обусловлена перераспределением ионов снаружи (привносятся на электроде) и внутри – катион перемещается к катоду.
Вместе со смещением мембранного потенциала смещается и уровень критической деполяризации – к нулю. При размыкании цепи постоянного тока под катодом мембранный потенциал быстро возвращается к исходному уровню, а УКД медленно, следовательно, порог увеличивается, возбудимость снижается – катодическая депрессия Вериго. Таким образом, ввозникает только при замыкании цепи постоянного тока под катодом.
Изменение возбудимости под анодом.
При замыкании цепи постоянного тока под анодом (допороговый, продолжительный раздражитель) на мембране развивается гиперполяризация за счет перераспределения ионов по обе стороны мембраны (без изменения ионной проницаемости мембраны) и возникающее за ней смещение уровня критической деполяризации в сторону мембранного потенциала. Следовательно, порог уменьшается, возбудимость повышается – анодическая экзальтация.
При размыкании цепи мембранный потенциал быстро восстанавливается к исходному уровню и достигает сниженного уровня критической деполяризации, генерируется потенциал действия. Таким образом, возбуждение возникает только при размыкании цепи постоянного тока под анодом.
Сдвиги мембранного потенциала вблизи полюсов постоянного тока получили название электротонических.
Сдвиги мембранного потенциала не связанные с изменением ионной проницаемости мембраны клетки называют пассивными.
Изменение возбудимости клеток или ткани под действием постоянного электрического тока называется физиологическим электротоном.Соответственно различают катэлектрон и анэлектрон (изменение возбудимости под катодом и анодом).
12) Закон раздражения Дюбуа-Реймона (аккомодации):
•раздражающее действие постоянного тока зависит не только от абсолютной величины силы тока или его плотности, но и от скорости нарастания тока во времени.
•При действии медленно нарастающегораздражителя возбуждение не возникает, так как происходит приспосабливание возбудимой ткани к действию этого раздражителя, что получило название аккомодации. Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране возбудимой ткани происходит повышение критического уровня деполяризации
При снижении скорости нарастания силы раздражителядо некоторого минимального значения потенциал действия вообще не возникает. Причина заключается в том, что деполяризация мембраны является пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению натриевой проницаемости, и тем самым обусловливающего возникновение потенциала действия, и медленного, приводящего к инактивации натриевой проницаемости и как следствие этого -окончанию потенциала действия.
•При медленном нарастании токана первый план выступают процессы инактивации, приводящие к повышению порога или ликвидации возможности генерировать ПД вообще. Способность к аккомодации различных структур неодинакова. Наиболее .высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка.
•При быстром нарастании стимулаповышение натриевой проницаемости успевает достичь значительной величины прежде, чем наступит инактивация натриевой проницаемости.
Аккомодация возбудимых тканей
•Раздражители характеризуются не только силой и длительностью действия, но и скоростью роста во времени силы воздействия на объект, т. е. градиентом.
•Уменьшение крутизны нарастания силы раздражителя ведет к повышению порога возбуждения, вследствие чего, ответ биосистемы при некоторой минимальной крутизне вообще исчезает. Это явление названо аккомодацией.
•Зависимость между крутизной нарастания силы раздражения и величиной возбуждения определена в законе градиента:реакция живой системы зависит от градиента раздражения: чем выше крутизна нарастания раздражителя во времени, тем больше до известных пределов величина функционального ответа.
Закон силы раздражения
Качественные и количественные изменения процессов, протекающих в организме, соответственно отображают качественные и количественные особенности действующих на него раздражителей и способа их действия на организм, т. е. раздражения.
Наименьшая сила раздражителя, которая вызывает минимальное возбуждение, называется порогом раздражения. Так как порог раздражения характеризует возбудимость, то он является вместе с тем и порогом возбудимости. Чем больше возбудимость, тем все более уменьшается порог раздражения, и, наоборот, чем меньше возбудимость, тем больше сила раздражения, которая вызывает наименьшее возбуждение. Порог возбудимости определяется на нервно-мышечном препарате по силе постоянного электрического тока, необходимого для получения едва заметного сокращения мышцы.
Чем больше сила раздражения, тем до определенного предела больше возбуждение и, следовательно, ответная реакция возбуждаемой ткани.
Сила раздражения меньше пороговой называется подпороговой, а больше пороговой — надпороговой. Наименьшая сила раздражения, вызывающая наибольшую ответную реакцию ткани, называется максимальной. Разные нарастающие величины силы разложения, расположенные между пороговой и максимально: называются субмаксимальными, а больше максимальной — сверхмаксимальными.
Порог возбудимости зависит от свойств возбудимой ткани от ее физиологического состояния в момент нанесения раздражения, от способа и продолжительности раздражения и от круп: нарастания силы раздражения.
Закон градиента раздражения (аккомодация)
В 1848 г. Дюбуа-Реймон обнаружил, что если через нерв или любую другую ткань проходит постоянный электрический ток пороговой силы и сила этого тока на протяжении значительного отрезка времени не меняется, то такой ток при своем прохождении не вызывает возбуждения ткани. Возбуждение возникает только в том случае, если сила электрического раздражителя быстро нарастает или убывает. При очень медленном нарастании силы тока раздражения нет. Закон Дюбуа-Реймона относится не только к действию электрического тока, но и к действию любого другого раздражителя. Это — закон градиента. Градиентом раздражения обозначается быстрота возрастания силы раздражения. Чем больше ее увеличение в каждую последующую единицу времени, тем до определенного предела больше реакция живой ткани на это раздражение. Быстрота нарастания возбуждения зависит от градиента раздражения. Возбуждение возрастает тем медленнее, чем меньше градиент раздражения.
Порог возбудимости значительно повышается при медленном нарастании раздражения. Можно предполагать, что живая ткань противодействует внешнему раздражению. Например, если быстро ударить по нерву, очень быстро его охладить или нагреть при силе раздражения выше пороговой, то возникает возбуждение. Если же. медленно надавливать на нерв, медленно его охлаждать или нагревать, то возбуждение не вызывается. Синусоидальный переменный электрический ток низкой частоты не вызывает возбуждение, так как скорость его изменения слишком мала. Следовательно, при медленном нарастании раздражения возникает приспособление, адаптация раздражаемой ткани к раздражителю Ш. С. Бетитов, X. С. Воронцов. Это приспособление называется аккомодацией.
Чем быстрее нарастает сила раздражения, тем до определенного предела сильнее возбуждение, и наоборот. Показатель скорости аккомодации — наименьшая крутизна нарастания силы раздражения, при которой оно еще вызывает возбуждение. Это пороговый градиент аккомодации.
У двигательных нервов аккомодация значительно больше, чем у чувствительных. Самая малая аккомодация у тканей, обладающих автоматизмом (сердечной мышцы, гладкой мускулатуры пищеварительного канала и других органов).
Закон гиперболы
Для получения возбуждения необходимо некоторое минимальное время раздражения постоянным электрическим током. Существует определенная зависимость между силой раздражающего постоянного электрического тока и временем раздражения, необходимым для возникновения возбуждения, или латентным периодом. Эта зависимость выражается кривой силы — времени, имеющей вид равносторонней гиперболы (Гоорвег, 1892, Вейс, 1901).
Закон гиперболы: каждому минимальному промежутку времени раздражения соответствует минимальная сила постоянного тока, при которой получается возбуждение, и наоборот. В современной физиологии существуют электронные приборы, позволяющие раздражать ткань в течение тысячных и меньше долей секунды или в микроинтервалах времени (0,001 с сокращенно обозначается буквой σ — сигма).
Чем сильнее ток, тем меньше продолжительность его действия, необходимая для получения возбуждения, и наоборот.
Полярный закон Пфлюгера
Пфлюгер (1859) установил, что при раздражении постоянным электрическим током возбуждение возникает в момент его замыкания или при возрастании его силы в области приложения к раздражаемой ткани отрицательного полюса — катода, откуда оно распространяется вдоль по нерву или мышце. В момент размыкания тока или при его ослаблении возбуждение возникает в области приложения положительного полюса — анода. При одной и той же силе тока возбуждение больше при замыкании в области катода, чем при размыкании в области анода. При раздражении нервно-мышечного препарата постоянным электрическим током получаются различные результаты в зависимости от его силы и направления. Различают входящие направление тока, при котором ближе к мышце расположен анод, и нисходящее — если ближе к мышце расположен катод.
При замыкании и прохождении постоянного тока через нерв или мышцу изменяются физиологические и физико-химические свойства на полюсах.
При прохождении постоянного тока в области приложения катода возбудимость временно повышается, а в области приложения анода возбудимость временно понижается. Даже слабые и кратковременные токи вслед за повышением возбудимости вызывают понижение возбудимости в области действия катода. Особенно отчетливо выступает это последующее понижение возбудимости в этой области при действии относительно сильных и продолжительных токов — катодическая депрессия (Б. Ф. Вериго, 1888). Катодическая депрессия может превратись проведение нервных импульсов (Д. С. Воронцов, 1937). Она исчезает через 7-8 мс после выключения постоянного тока.
В области действия катода при замыкании увеличивается скорость проведения возбуждения, а в области действия анода она уменьшается. В области действия катода уменьшается высота волны возбуждения и увеличивается её продолжительность, а в области действия анода, наоборот, высота увеличивается, а продолжительность её уменьшается. Продолжительность полной невозбудимости в области действия катода увеличивается, а анода – уменьшается. Поэтому лабильность в области действия катода уменьшается, а в области действия анода увеличивается.
Эти изменения физиологических свойств нерва в области действия катода обозначаются как катэлектротон, а в области действия анода — как анэлектротон. Изменения физиологических свойств нерва происходят не только в месте приложения полюсов постоянного тока, но и на некотором расстоянии от них. На расстоянии около 2 см за пределами катода возбудимость нерва понижается, а за пределами анода она повышается. Этот факт открыл Н. Я. Перна (1914) и обозначил его как периэлектротон.
Следовательно, в периферических нервах не только распространяются волны возбуждения, но при появлении очага возбуждения на некотором расстоянии от него возникают и устанавливаются вдоль всего нерва участки повышенной и пониженной возбудимости в виде стационарной волны. Таким образом, в периферических нервах имеется двойная нервная сигнализация: импульсная и тоническая. Некоторые авторы отрицают существование периэлектротона (Д. С. Воронцов, 1961).
В пунктах приложения полюсов постоянного тока увеличивается количество ацетилхолина в области действия катода и уменьшается в области действия анода, относительно увеличивается содержание ионов калия в области действия катода и ионов кальция в области действия анода, увеличивается проницаемость белковых мембран в области действия катода и уменьшается их проницаемость в области действия анода.
Изменения возбудимости нервов при действии постоянного тока наблюдаются и у человека. Электрод с малой поверхностью, или дифферентный, прикладывают к раздражаемой области нерва, а электрод с большой поверхностью, или индифферентный, накладывают на отдаленную часть тела. При таком униполярном способе раздражения действие тока проявляется только вблизи дифферентного электрода. В зависимости от силы тока получают различные результаты.
При слабой силе постоянного тока раздражение в области анода является подпороговым. Поэтому независимо от направления тока сокращение получается только в области катода, так как возбуждение на этом полюсе больше, чем на аноде. При средней силе тока раздражение в области анода достигает порога. Поэтому независимо от направления тока сокращения получаются и в области катода и в области анода.
При сильном восходящем токе возбуждение возникает в области катода при замыкании, но оно не может достигнуть мышцы, так как по пути возникает анэлектротон (резкое понижение возбудимости и проводимости), поэтому сокращение получается только при размыкании. При сильном нисходящем токе замыкание вызывает сокращение мышцы, а при размыкании сокращения нет. Это отсутствие сокращения зависит от того, что в момент размыкания в области катода возбудимость и проводимость резко понижаются и возбуждение, возникающее на аноде, не проводится к мышце.
Полярный закон раздражения (закон Пфлюгера)

При замыкании цепи постоянного электрического тока возбуждение возникает только под катодом, а при размыкании - только под анодом
Доказать правильность закона можно при помощи опыта, в котором меняется расположение катода и анода на поврежденном и неповрежденном участках нерва, иннервирующего мышцу
Прохождение постоянного электрического тока через мембрану вызывает изменение мембранного потенциала покоя.
Так, при замыкании цепи возле катода скапливается его "-" заряд, который уменьшает "+" заряд наружной поверхности мембраны.
Разность потенциалов (между наружной и внутренней поверхностями мембраны) уменьшается, и мембранный потенциал изменяется в сторону уровня критической деполяризации, т.е. формируется деполяризация(возбудимость повышается).
Достижение критического уровня приводит к возникновению пикового потенциала (потенциала действия)
При замыкании цепи возле анода скапливается его "+" заряд.
Он увеличивает "+" заряд мембраны и величину мембранного потенциала
Мембранный потенциал удаляется от критического уровня, превышает значение потенциала покоя и формируетгиперполяризацию (возбудимость уменьшается)
При размыкании цепи, прекращение поступления дополнительного "+" заряда от анода приводит к уменьшению(восстановлению) заряда наружной поверхности мембраны.
Мембранный потенциал, уменьшаясь, приближается к критическому уровню
Формируется деполяризация (возбудимость увеличивается).
После достижения критического значения развивается пиковый потенциал
(Поскольку размыкание происходит после замыкания, а, следовательно, на фоне гиперполяризации и пониженной возбудимости, то для возникновения ПД необходим раздражитель, превышающий по силе пороговый - это анодно-размыкательный эффект)
При размыкании возле катода прекращается накопление его "-" заряда
Заряд наружной поверхности мембраны увеличивается (восстанавливается), мембранный потенциал, увеличиваясь, удаляется от критического уровня и возбуждение не возникает
Закон физиологического электротона
Действие постоянного электрического тока на ткань сопровождается изменением ее возбудимости
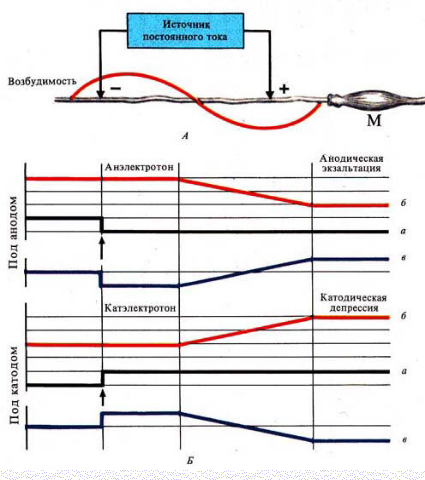

Различают 3 вида физиологического электротона или изменения возбудимости:
Катэлектротон - изменение возбудимости под катодом.
В момент замыкания под катодом формируется деполяризация и возбудимость повышается.
По мере удаления от катода количество его "-" зарядов, а следовательно и выраженность деполяризацииуменьшается.
В результате возбудимость уменьшается, но она остается выше, чем в состоянии покоя
Анэлектротон - изменение возбудимости под анодом.
В момент замыкания под анодом формируется гиперполяризация и возбудимость снижается.
По мере удаления от анода количество его "+" зарядов, а, следовательно, и гиперполяризация уменьшается.
В результате возбудимость увеличивается, но она остается ниже, чем в состоянии покоя
Периэлектротон - обратное изменение возбудимости вне электротонических областей.
В зоне прекращения действия катода возбудимость снижается.
В зоне прекращения влияний анода она, наоборот, увеличивается
При длительном прохождении постоянного тока через ткань происходит извращение измененной возбудимости
При длительном замыкании под катодом происходит:
·инактивация Na проницаемости
·повышение уровня критической деполяризации
·увеличение порога деполяризации
·снижение вначале повышенной возбудимости
Это явление получило название катодическая депрессия
При длительном замыкании под анодом происходит:
·снижение К проницаемости
·уровень критической деполяризации снижается
·порог деполяризации уменьшается
·повышается вначале сниженная возбудимость
Это явление получило название анодическая экзальтация
Этот закон необходимо учитывать в медицинской практике
С одной стороны, он может быть использован, если требуется заблокировать проведение возбуждения по нервной или мышечной ткани (катодическая депрессия) или повысить возбудимость (анодическая экзальтация)
С другой стороны, необходимо помнить о возможности привыкания к длительному действию раздражителей, в частности, фармацевтических препаратов, влияющих на возбудимость мембраны.
ФИЗИОЛОГИЯ МЫШЦ И НЕРВОВ
Структура миофибрилл и ее изменения при сокращении.Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечнополосатых мышечных волокнахмиофибриллы-разделены на правильно чередующиеся участки (диски), обладающие разными оптическими свойствами. Одни участки анизотропны,т. е. обладают двойным лучепреломлением. В обыкновенном свете они ^выглядят темными, а. в поляризованномг— прозрачными в продольном направлении й непрозрачными в поперечном. Другие участки в Обыкновенной свете выглядят светлыми — они изотропны,т. е. не ; обладают двойным' лучепреломлением (рис. 34, а) . Анизотропные участки обозначают буквой А,изотропные — буквой I.В середине диска Аразличается светлая полоска Н, посередине диска / — темная "полоска -представляющая собой тонкую мембрану, сквозь поры которой проходят миофибриллы. Благодаря наличию этой опорной структуры параллельно расположенные однозначные диски отдельных фибрилл внутри одного волокна во время сокращения не смещаются по,отношению друг к другу.
Современные представления о структуре миофибриллярного аппарата основываются на исследованиях структуры мышечного волокна-при помощи электронной микроскопии, рентгеноструктурного анализа, фазово-контрастной и интерференционной микроскопии в сочетании с гистохимическими методами.
Установлено, что каждая миофибрилла мышечного волокна диаметром около 1 мкм состоит в среднем из 2500 протофибрилл,представляющих собой удлиненные полимери-, зованные молекулы белков миозина и актина; Миозиновые протофибриллы, или, как, их принято обозначать, нити, вдвое толще актиновых. Их диаметр,примерно 10 нм. В состоянии покоя мышечного волокна нити расположены в миофибрилле таким образом, ^что тонкие длинные актиновые нити входят своими концами в промежутки между толстыми и более короткими миозиновыми нитями (рис. 34, б). Благодаря этому диски / состоят только из актиновых нитей, а диски А — из нитей миозина, а, возможно, еще и другого белка.


Рис. 34. Структура миофибриллы (схема), Показаны диски А и 1-полоски Zи Н(а). Взаимное расположение толстых (миозиновых) и тонких ; (актиновых) нитей в расслабленной (б) й сокращённой (в) миофибрилле.
Молекула актина
Это глобулярный белок, состоящий из одного полипептида, который полимеризуется с другими молекулами актина и образует две цепи, обвивающие друг друга (рис. 4-4 А). Такая двойная спираль представляет собой остов тонкого филамента. На каждой молекуле актина есть участок связывания миозина. В покоящемся мышечном волокне взаимодействие между актином и миозином предотвращают два белка - тропонин и тропомиозин (рис. 4-4 Б).

Тропомиозин - стержневидная молекула из двух полипептидов, обвивающихся друг около друга; молекула соответствует в длину примерно семи мономерам актина. Цепи из молекул тропомиозина, уложенные конец в конец, располагаются вдоль всего тонкого филамента. Молекулы тропомиозина частично прикрывают участки связывания каждой молекулы актина, мешая контакту миозина с актином. В таком блокирующем положении молекула тропомиозина удерживается тропонином.
Тропонин - гетеротримерный белок. Он состоит из тропонина Т (отвечает за связывание с одиночной молекулой тропомиозина), тропонина С (связывает ион Са 2+ ) и тропонина I (связывает актин и ингибирует сокращение). Каждая молекула тропомиозина связана с одной гетеротримерной молекулой тропонина, которая регулирует доступ к участкам связывания миозина на семи мономерах актина, прилегающих к молекуле тропомиозина.
1) закон полярного раздражения
3) закон порога (силы раздражения)
4) закон крутизны
5) закон длительности действия
1. Закон полярности (Пфлюгер) – в момент замыкания раздражения тока или в момент увеличения его силы, заряд возникает в области катода (отрицат полюс), при ослаблении тока (в момент размыкания) - в области анода. При одной и то же силе раздражающее действие замыкания выражается сильнее, чем действие размыкании. В случае электродов снаружи:
1) при включении тока раздражение возникает в области катода.
2) при выключении – в области а-да.
Закон электротона. Изменение полярности мембран изменяет величины МП, создаваемое пропусканием через данный участок мембраны эл тока от внешнего источника приводит к изменению возбудимости. Существует катэлектротон при выходящем токе, и анэлектротон при входящем токе.
При действии катода – выход тока – происходит разрядка мембранной емкости, при этом возбудимость и проводимость оказываются повышенными. Под анодом идет процесс дозарядки мемб-уменьшается возбудимость и проводимость.
Законы раздражения возбудимых тканей. Соотношение между силой и временем раздражения. Хронаксиметрия.
1) закон полярного раздражения
3) закон порога (силы раздражения)
4) закон крутизны
5) закон длительности действия
Закон крутизны. При раздражении деполяризаций ток должен нарастать круто. Если промежуток подачи тока велик, то происходит смещения уровня КУД в + сторону и изменяющийся в этом же направлении МП его не догоняет. Позитивное смещение КУД при длительной деполяризации - аккомодация – приспособление ткани к току. Если ток нарастает очень медленно, то он никогда не догонит КУД и никогда не возникнет ПД.
Закон длительности действия. Каждому напряжению тока соответствует мин. длительность его воздействия на ткань, чтобы ток смог вызвать возбуждение. Если при данном напряжении удлинять время прохождения тока через ткань сверх мин длительности, то никакого изменения в наступлении эффекта возбуждения не происходит.
Даже очень большое напряжение, если действ очень короткий промежуток времени не может вызвать возбуждение. Очень слабые раздражители как бы долго они не действовали, не вызывают возбуждения. Ответная реакция зависит от времени действия тока. Существует минимальное напряжение, которого достаточно при неограниченно долгом действии тока, чтобы вызвать возб. - реобаза. Мин время, которое необходимо, чтобы вызвать возб током в одну реобазу – полезное время. Хронаксия – время тока в 2 реобазы, прив к возб. Величина хронаксии находится во взаимосвязи со скоростью реакции: чем быстрее реагирует ткань, тем короче ее хронаксия. Хронаксия – миним время и миним напряжение, которое дает положительный эффект.
Законы раздражения возбудимых тканей. Адекватные и неадекватные раздражители. Порог раздражения.
1) закон полярного раздражения
3) закон порога (силы раздражения)
4) закон крутизны
5) закон длительности действия
Закон порога. Для возбудимых элементов существует минимальная сила раздражителя, необходимая для миним по величине возбуждения, получившая название порог возбуждения. Величина порога является мерой возбудимости ткани. Т.е. порог – минимальная сила раздражителя, при которой возникает минимальная величина возбуждения.
По своему физиологическому значению все раздражители делят на
адекватные и неадекватные.
Адекватными называются те раздражители, которые действуют на данную
биологическую структуру в естественных условиях, к восприятию которых она
специально приспособлена и чувствительность к которым у нее чрезвычайно
велика. Для палочек и колбочек сетчатки глаза адекватным раздражителем
являются лучи видимой части солнечного спектра, для тактильных рецепторов
кожи - давление, для вкусовых сосочков языка - разнообразные химические
Неадекватными называются те раздражители, для восприятия которых данная
клетка или орган специально не приспособлены. Так, мышца сокращается при
воздействии кислоты или щелочи, электрического тока, внезапного растяжения,
механического удара, быстрого согревания и т. д.
Клетки значительно более чувствительны по отношению к своим адекватным
раздражителям, чем к неадекватным. Это является выражением функционального
приспособления, выработавшегося в процессе эволюции.
Кроме всеобщих законов раздражения, которые применимы к любым раздражителям, специфические законы характеризуют закономерности действия постоянного электрического тока, прохождение которого через нервное или мышечное волокно вызывает изменение мембранного потенциала покоя и возбудимости у места приложения электродов имеющих разный заряд. Отметим, что речь идет именно о постоянном, а не о переменном токе, действие которого носит совершенно специфический характер
Закон полярного действия постоянного тока.
Закон не имеет однозначной формулировки и характеризует изменение мембранного потенциала и вероятность возникновения возбуждения мембраны у места приложения электродов. Поскольку при этом всегда возникает электрический ток, направленный от области положительного заряда к области отрицательного заряда, то в наиболее общем виде закон звучит так: возникновение возбуждения происходят при действии на клетку выходящего тока. При действии входящего тока происходит противоположные изменения – гиперполяризация и снижение возбудимости, возбуждение не возникает.
При внеклеточном раздражении возбуждение возникает в области катода (–). При внутриклеточном раздражении для возникновения возбуждения необходимо, чтобы внутриклеточный электрод имел положительный знак (рис. 6).
Рис. 6. Изменения, наступающие в нервном волокне при внутриклеточном раздражении (А, Г) и при внеклеточном раздражении в области анода (Б) и катода (В). Стрелкой показано направление электрического тока.
Следует отметить, что механизм инициации возбуждения определяется не столько направлением тока, сколько зарядом электрода. Кроме того, имеет значение, замыкается или размыкается электрическая цепь. Поэтому в более полном варианте закон полярного действия постоянного тока звучит так: при замыкании тока возбуждение возникает под катодом (-), а при размыкании - под анодом (+).
Действительно, при замыкании цепи, в области приложения катода (-), положительный потенциал на наружной стороне мембраны уменьшается, заряд мембраны снижается, это активирует механизм переноса Na+ внутрь клетки, при этом мембрана деполяризация. Как только деполяризация достигнет критического уровня (КУД) ), ткань возбуждается — генерируется ПД.
В области же приложения анода (+), положительный потенциал на наружной стороне мембраны возрастает, происходит гиперполяризация мембраны и возбуждение не возникает.
При этом возбудимость ткани сначала снижается из-за увеличения порогового потенциала, а затем начинает повышаться в результате его уменьшения, так как анод уменьшает количество инактивированных потенциалзависимых Na-каналов. КУД смещается в сторону увеличения и при определенной силе гиперполяризующего тока постепенно выходит на уровень исходной величины мембранного потенциала.
При размыкании постоянного тока мембранный потенциал под анодом возвращается к норме, одновременно выходя на КУД; при этом ткань возбуждается — запускается механизм генерации ПД.
Закон физиологического электротона.
Этот закон иногда объединяют с предыдущим, но в отличие от нег он характеризует изменения не мембранного потенциала, а возбудимости ткани, при прохождении через неё постоянного тока. Кроме того, он применим только в случае внеклеточного раздражения.
Изменения возбудимости достаточно сложные и зависят как от заряда приложенного к поверхности электрода, так и от времени действия тока, поэтому в общем виде закон можно сформулировать так: действие постоянного тока на ткань сопровождается изменением ее возбудимости (рис 7).
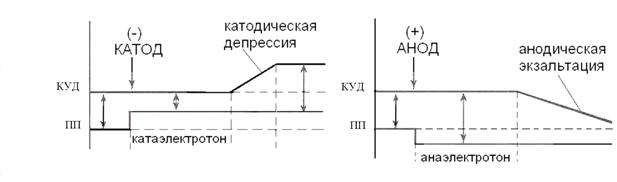
Рис. 7. Изменения возбудимости при действии на ткань постоянного тока под катодом (-) и анодом(+).
При прохождении постоянного тока через нерв или мышцу порог раздражения под катодом (-) и соседних с ним участках понижается вследствие деполяризации мембраны - возбудимость повышается. В области приложения анода происходит повышение порога раздражения, т. е. снижение возбудимости вследствие гиперполяризации мембраны. Эти изменения возбудимости под катодом и анодом получили название электротона (электротоническое изменение возбудимости). Повышение возбудимости под катодом называется катэлектротоном, а снижение возбудимости под анодом - анэлектротоном.
При дальнейшем действии постоянного тока первоначальное повышение возбудимости под катодом сменяется ее понижением, развивается так называемая катодическая депрессия. Первоначальное же снижение возбудимости под анодом сменяется ее повышением - анодная экзальтация. При этом в области приложения катода происходит инактивация натриевых каналов, а в области действия анода происходит снижение калиевой проницаемости и ослабление исходной инактивации натриевой проницаемости.
ПРАКТИЧЕСКИЕ ЗАДАНИЯ

1. Анализ компонентов биологического потенциала.
Одиночный цикл возбуждения характеризуется электрографическими, функциональными и электрохимическими показателями.
Первый – регистрируется в виде кривой потенциала действия (ПД), отражающей изменение мембранного потенциала в процессе одиночного цикла возбуждения
Второй – связан с изменением возбудимости мембраны и графически отражается кривой изменения возбудимости
Третий – характеризует электрическое состояния плазматической мембраны возбудимой клетки обеспечиваемое её транспортными системами в каждую фазу развития потенциала действия.
Анализ процессов, которые обеспечивают эти состояния, в реальном времени позволяет понять физиологическую сущность и механизм процесса возбуждения, а значит, объяснить и предсказать реакцию клетки на её раздражение. Это может иметь важное значение в изучении механизмов, лежащих в основе деятельности нервной системы, в регуляции как физиологических, так и психических процессов.
О с н а щ е н и е : схемы записи потенциала действия (ПД).
С о д е р ж а н и е р а б о т ы. Проанализируйте по имеющимся схемам фазы развития потенциала действия ПД на мембране возбудимой клетки (рис. 8).
1. Зарисуйте ПД; обозначьте его фазы.
2. Отметьте направление ионных токов характеризующие каждую из фаз потенциала действия.
3. Сопоставьте фазы ПД и колебания возбудимости клетки, объясните причины не возбудимости клетки в некоторые фазы развития ПД.
4. Охарактеризуйте состояние мембраны в каждую фазу развития ПД, объясните, почему даже при самой высокой частоте раздражения возникновение ПД клетки имеет дискретный характер.
Читайте также:
