
Потенциал действия миелинизированного нервного волокна
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.

Передача потенциала действия по нервным волокнам
При передаче информации между различными участками нервной системы, отстоящими друг от друга на значительные расстояния, необходимо распространение нервных импульсов по аксонам нейронов. Скорость проведения зависит от толщины и наличия миелиновой оболочки. Имеется два типа нервных волокон: миелиновые и безмиелиновые. Оболочку безмиелиновых волокон образуют шванновские клетки, мембрана которых не имеет миелина. Оболочку миелиновых волокон в периферической нервной системе формируют шванновские клетки с миелином, а в ЦНС – сами олигодендроциты. Миелиновая оболочка через равные промежутки (0,5-2,0 мм) прерывается, образуя, свободные от миелина участки – узловые перехваты Ранвье. Протяженность этих перехватов в периферической нервной системе составляет 0,25 -1,0 мкм, а в волокнах ЦНС их длина достигает 14 мкм. Миелиновая оболочка нервных волокон выполняет изолирующую функцию, обеспечивает более экономное и быстрое проведение возбуждения.
В зависимости от толщины нервных волокон, наличия или отсутствия у них миелиновой оболочки все нервные волокна делят на три основных типа: А, В, и С.
1. Волокна типа А – это наиболее толстые, хорошо миелинизированные афферентые и эфферентые волокна соматичекой нервной системы. Скорость проведения этих волокон варьирует от 120 м/с до 15 м/с.
2. Волокна типа В слабомиелинизированные преганглионарные (парасимпатичесике) волокна вегетативной нервной системы. Скорость проведения составляет 5 – 14 м/с.
3. Волокна типа С – это немиелинизированные в основном постганглионарные (симпатические) волокна вегетативной нервной системы. Скорость проведения от 0,5 до 2,3 м/с.
Механизм проведения потенциала действия. Проведение возможно только при наличии на всем протяжении или ограниченных, но повторяющихся участках волокна потенциалзависимых ионных каналов, ответственных за формирование новых ПД. В распространении ПД можно выделить два этапа: этап распространения электрического поля, которое снижает МП, и этап генерации новых ПД в новых участках нервного волокна.
Электрическое поле – разновидность материи, посредством которой осуществляется силовое воздействие на электрические заряды, находящиеся в этом поле. Электрическое поле, которое генерируется биологическими структурами, является источником информации о состоянии клеток и органов организма.
В зависимости от расположения и концентрации ионных каналов в мембране нервного волокна имеются два варианта проведения ПД: непрерывный и сальтаторный.
Непрерывное проведение ПД происходит в безмиелиновых нервных волокнах (тип С), имеющих равномерное распределение потенциалзависимых ионных каналов по всей длине волокна, которое участвует в генерации ПД. Проведение нервного импульса начинается с распространения электрического поля. Амплитуда ПД в нервном волокне составляет 100-120 мВ. Расстояние, на котором сохраняется 37% величины ПД в виде электрического поля (постоянная длины мембраны), в немиелинизированных волокнах составляет от 0,1 до 1,0 мм. Возникший ПД за счет электрического поля деполяризует мембрану соседнего участка до критического уровня на постоянную длины мембраны (0,1 до 1,0 мм). Это означает, что на этом участке одновременно генерируются новые ПД, обусловленные движением ионов Na+ в клетку и ионов К+ из клетки. Число одновременно возникающих ПД ограничивается длиной возбужденного участка (от 0,1 до 1,0 мм для немиелинизированных волокон). ПД возникают рядом друг с другом в непосредственной близости. Причем сами ПД не перемещаются. Они исчезают там, где возникают. Главную роль в возникновении новых ПД играет передний ПД. Вспомогательную роль в генерации новых ПД в невозбужденных участках нервного волокна играют соседние ПД (возникшие сзади переднего ПД), так как их электрическое поле суммируется с электрическим полем переднего ПД. Таким образом, непрерывное распространение нервного импульса идет через генерацию новых ПД по эстафете, когда каждый участок мембраны сначала выступает как раздражаемый электрическим полем, а затем как раздражающий (в результате формирования в нем новых ПД).

Рис.1. Непрерывное проведение ПД по немиелинизированному нервному волокну.
Сальтаторное проведение ПД по миелинизированным волокнам является эволюционно более поздним механизмом. Оно происходит в волокнах типа А и В (миелинизированных), для которых характерна концентрация потенциалзависимых ионных каналов только в небольших участках мембраны – в перехватах Ранвье. В области миелиновых муфт, обладающих хорошими изолирующими свойствами, потенциалзависимых каналов нет, поэтому ПД здесь не возникают. Участок нервного волокна, покрытый миелиновой муфтой, в механизме проведения потенциала действия играет роль изолятора. В этих условиях ПД, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до критического уровня, что приводит к возникновению в них новых ПД, то есть возбуждение проводится скачкообразно от одного перехвата к другим. Так как, Nа+- каналы начинают открываться при достижении деполяризации клеточной мембраны 50%-ов, а постоянная длины мембраны миелинизированного волокна составляет 5 мм, то электрическое поле ПД на данном расстоянии сохраняет 37% своей амплитуды (это около 30 мВ) и может деполяризовать мембрану до критического уровня (пороговый потенциал в перехватах равен 15 мВ) не только соседнего перехвата, но и вплоть до пятого. Поэтому возбуждение распространяется очень быстро по всей длине волокна. При этом ионы движутся только перпендикулярно относительно длины волокна. Электрическое поле потенциалов действия, возникших сзади переднего ПД, суммируется с его электрическим полем, так как и при непрерывном распространении возбуждения. Следовательно, сальтаторное проведение возбуждения обусловлено генерацией новых ПД по эстафете, когда каждый перехват Ранвье сначала выступает как раздражаемый электрическим полем, а затем как раздражающий соседние перехваты.

Рис.2. Сальтаторное проведение ПД в миелинизированном нервном волокне.
Возникающие ПД не могут инициировать возникновение других ПД в обратном направлении, так как нервное волокно находится еще в рефрактерном состоянии. В натуральных условиях первый потенциал действия, инициирующий распространение возбуждения по аксону, возникает в аксонном холмике, а возбуждение проводится только в одном направлении – по аксону к другой клетке.
Сравнение непрерывного и сальтаторного проведения возбуждения показывает, что различие в механизме проведения возбуждения по миелинизированным и немиелинизированным нервным волокнам заключается в том, что очередные ПД в безмякотном волокне возникают на более близком расстоянии друг от друга, так как ионные каналы расположены близко и непрерывно по всей длине. Поэтому такое проведение и назвали непрерывным. Число одновременно возникающих ПД в мякотном волокне строго ограничено числом возбужденных перехватов – максимально 5.
Сальтаторное проведение возбуждения имеет два важных преимущества по сравнению с непрерывным проведением возбуждения.
1. Сальтаторное проведение более экономично в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых имеет 1% мембраны, и, следовательно. Надо меньше энергии для восстановления трансмембранных градиентов Nа+ и К+, расходующихся в процессе возникновения ПД.
2. Возбуждение в миелинизированных волокнах проводится с большей скоростью, чем в безмиелиновых волокнах, так как в них электрическое поле ПД в области миелиновых муфт распространяется значительно дальше – на соседние перехваты Ранвье, поскольку электроизоляция уменьшает рассеивание электрического поля. Кроме того, миелинизированные волокна в большинстве своем тольще немиелинизированных, что также ускоряет проведение возбуждения, поскольку электрическое сопротивление более толстых волокон меньше.
Время распространения ПД в миелинизированных и немиелинизированных нервных волокнах определяется только временем возникновения ПД, то есть перпендикулярным относительно мембраны движением ионов в клетку и из клетки, поскольку электрическое поле распространяется мгновенно.
Характеристика проведения возбуждения по нервным волокнам.
1. Нервные волокна могут проводить возбуждения в двух направлениях. В эксперименте при раздражении любого участка нерва, ПД распространяется в обе стороны от места раздражения.
2. Возбуждение проводится изолировано в каждом нервном волокне.
3. Большая скорость проведения возбуждения (до 120 м/с). По сравнению – гуморальное проведение информации по организму совершается за 22 с.
4. Малая утомляемость нервного волокна. При нормальной доставке кислорода и питательных веществ к нервному волокну – оно практически не утомляемо, так как расход энергии в нем очень мал.
5. Возможность функционального блока проведения возбуждения при морфологической целостности волокон. Наличие явления парабиоза – функциональных изменений в нерве после действия на него длительных и сильных раздражений. Причиной блока проведения возбуждения является инактивация Nа+ - каналов.
Аксонный транспорт. Основная масса веществ, образующихся в теле нейрона, используется в различных его отделах. Различают аксонный транспорт и дендритный. Аксонный транспорт изучен лучше. Он делится на быстрый и медленный.
1. Быстрый аксонный транспорт идет в двух направлениях: от тела клетки до аксонных окончаний – антеградный транспорт, и в противоположном направлении – ретроградный транспорт. Посредством антеградного транспорта в окончания аксона доставляются ферменты, медиаторы, липиды, везикулы, содержащие гликопротеины мембран. При помощи ретроградного транспорта в тело нейрона переносится везикулы, содержащие остатки разрушенных структур, фрагменты мембран, ацетилхолинэстераза. В патологическиъ условиях по аксону к соме могут транспортироваться вирусы бешенства, герпеса, полиомиелита, столбнячный токсин. Быстрый аксонный транспорт осуществляется с помощью специальных структурных элементов нейрона: микротрубочек и микрофиламентов. Для транспорта необходима энергия АТФ.
2. Медленный аксонный транспорт осуществляется только в антеградном направлении и представляет собой передвижение всего столба аксоплазмы. С помощью медленного транспорта перемещаются белки микротрубочек и микрофиламентов, ферменты цитозоля, РНК, белки каналов, насосов.
Аксонный транспорт играет важную роль также и при регенерации поврежденных нервных волокон.
Впервые скорость распространения потенциала действия (возбуждения) по нервному волокну была измерена профессором физиологии Кенигсбергского университета Германом Гельмгольцем в 1850 году – спустя год после того как ученый И. Физо измерил скорость распространения света. Оказалось, что скорость распространения возбуждения составляет всего порядка 30 м/с. Значения скорости распространения ПД было примерно в 10 000 000 меньше скорости распространения электрического тока по металлическому проводнику и даже в 10 раз медленнее скорости распространения звука в воздухе.
Полученные результаты, с одной стороны, нанесли удар по сторонникам теории мгновенного распространения возбуждения, но одновременно и поставили исследователей перед необходимостью более детально изучить различия механизмов проведения электрического потенциала в проводниках и нервном волокне. Прежде всего, следовало дать ответ на вопросы: почему возбуждение способно распространяться по нервному волокну и от чего зависит скорость распространения нервного импульса.
Для того чтобы ответить на эти вопросы, рассмотрим электрические свойства нервного волокна. Оно представляет собой цилиндр, боковую поверхность которого образует мембрана, отделяющая внутренний раствор электролита от наружного. Это придает волокну свойства коаксиального кабеля, изоляцией которого служит клеточная мембрана. Но нервное волокно — очень плохой кабель. Сопротивление изоляции этого живого кабеля примерно в 105 раз меньше, чем у обычного кабеля, так как в первом случае ее толщина составляет 10
6 см, а во втором — около 10 см. Кроме того, внутренняя жила живого кабеля — это раствор электролита, удельное сопротивление которого в миллионы раз больше сопротивления металла. Поэтому невозбужденное нервное волокно плохо приспособлено для передачи электрических сигналов на большие расстояния. Было установлено, что уменьшение электрического потенциала на мембране нервного волокна по мере удаления от источника возбуждения убывает по экспоненциальному закону
где j0 – значение потенциала в точке возбуждения, jl –значение потенциала в точке, расположенной на расстоянии l от источника возбуждения, l - константа длины нервного волокна, равная расстоянию, на котором величина потенциала убывает в e (е=2,718281828…) раз.
Константа l зависит от удельного электрического сопротивления оболочки нервного волокна rm, удельного электрического сопротивления цитоплазмы ri, радиуса нервного волокна r:

Чем больше l, тем больше скорость распространения нервного возбуждения. Как следует из приведенной выше формулы, l тем больше, чем больше радиус нервного волокна и чем больше удельное электрическое сопротивление мембраны нервного волокна.
Для повышения скорости Природа использовала обе возможности. Для простейших организмов она создала нервные волокна большого диаметра – можно вспомнить гигантский аксон кальмара, диаметр которого достигает 0,5 мм.
Рассмотрим процесс распространения возбуждения по безмиелиновым нервным волокнам.
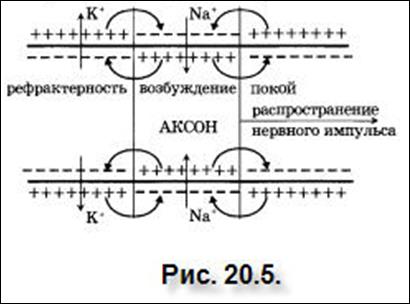
При возбуждении участка нервного волокна (см. рис. 20.5) происходит деполяризация данного участка. При деполяризации с наружной стороны мембран возникает отрицательный электрический потенциал, а с внутренней стороны – положительный потенциал. Между возбужденным и невозбужденным участками нервного волокна возникнут локальные электрические токи, так у возбужденного участка внутренняя поверхность имеет положительный заряд, а у невозбужденного участка – отрицательный электрический заряд и между ними возникает разность потенциалов. По поверхности нервного волокна локальный электрический ток течет от невозбужденного участка к возбужденному, внутри волокна электрический ток течет в обратном направлении – от возбужденного участка к невозбужденному.
Эти локальные токи являются раздражителями для невозбужденных участков, непосредственно примыкающих к возбужденному участку. В них также возникает возбуждение (потенциал действия). В то же время локальные токи приводят к тому, что возбужденные участки приходят в состояние покоя – в них ПД сменяется ПП.
Возбужденные участки, в свою очередь, предают сигнал дальше, и процесс распространяется вдоль аксона. (Существует внешняя аналогия между распространением нервного импульса и распространением огня по бикфордову шнуру).
В миелинизированном волокне распространение импульса возбуждение происходит за счет локальных токов, которые вынуждены циркулировать между соседними перехватами Ранвье (см. рис. 20.6). Передача возбуждения происходит быстрее, чем в немиелинизированном волокне.

ü Практически выполнить:
| Лабораторная работа: | Компьютерное моделирование электрогенеза в клетках |
| Цель работы: | На основании математической модели Ходжкина-Хаксли, реализованной в среде пакета MathCad, исследовать особенности генерации потенциала действия. |
| Аппаратное и программное обеспечение: | ПК на основе МП Intel Celeron или AMD. Операционная система MS Windows XP. Пакет программ MathСad |
I. Теоретическая часть
С электрической точки зрения, клеточную мембрану можно представить как параллельно включенные конденсатор емкости и два источника натриевого и калиевого токов, имеющих Э.Д.С, направленных друг против друга, и внутренние сопротивления RK и RNa. Эти сопротивления являются переменными и зависят от калиевой GK и натриевой GNa проводимости (см. рис. 20.7).

Ионы калия проходят в обе стороны через мембрану всегда. Когда потенциал на мембране V равен равновесному значению нернстовского потенциала VK, то на мембране сохраняется динамическое равновесие, суммарный поток ионов калия равен нулю, и калиевый ток равен нулю. При отклонении мембранного потенциала от равновесного значения возникает калиевый ток, величина которого определяется выражением:
Аналогичную формулу можно записать и для натриевого тока:
Суммарный ток, который протекает через конденсатор С (см. рис.1)
равен сумме натриевого, калиевого токов и тока I(t), подающегося на мембрану извне (например, тока возбуждения, если мы задались целью моделировать процесс генерации потенциала действия ПД):
Силу тока, проходящего через конденсатор, можно определить, зная скорость изменения заряда на конденсаторе:
Объединяя уравнения (1) – (4), получим:
На основании многочисленных экспериментов Ходжкин и Хаксли, с учетом уравнения (5), получили следующую систему уравнений, которая описывает все многообразие взаимосвязанных изменений во времени электрических характеристик мембраны:
Система уравнений (6) - (17) была создана Ходжкиным и Хаксли в 1952 г. и называется математической моделью Ходжкина–Хаксли (Х-Х). Для расчетов модели в те годы компьютеры недоступны (из-из их экзотичности – первая ЭВМ была создана в США в 1946 г. и использовалась почти исключительно для военных нужд) и Ходжкин с Хаксли вручную рассчитали, как меняется мембранный потенциал с течением времени, если на мембране создать возбуждение.
Сегодня при наличии персональных компьютеров (ПК) и современного программного обеспечения, реализация модели Х-Х (численные расчеты) не составит труда даже студенту-первокурснику.
Из программных сред, используемых для моделирования в области медицины и биологи на сегодняшний день, наиболее адекватным является пакет MathCad, в котором реализована идея «автоматизированного математического проектирования. Идея математического автоматизированного проектирования (МАП) позволяет специалисту-медику, который не имеет фундаментальных познаний в области численных методов и программирования, изучать и моделировать физиологические процессы, протекающие в живых организмах.
II. Практическая часть
q Исходное состояние – включен ПК, загружена операционная система Windows XP и имеется доступ к ресурсам локальной компьютерной сети (для получения доступа в сеть необходимо при загрузке Windows XP ввести имя пользователя Worker и парольWork).
1. С помощью созданного на Рабочем столе ярлыка загрузить пакет Mathcad.
2. Через команду “Файл” из меню команд загрузить файл c:\Мои документы\stud\ModelXX.mcd, содержащий модель ХХ, реализованную в среде пакета Mathcad.
3. В качестве исходных данных ввести следующие значения:
6. Закрыть Windows- приложение (пакет Mathcad).
8. Выключить ПК (если дана на то команда преподавателя) в соответствии с приведенной ниже инструкцией.
Все клетки возбудимых тканей при действии различных раздражителей достаточной силы способны переходить в состояние возбуждения. Обязательным признаком возбуждения является изменение электрического состояния клеточной мембраны. Общее изменение разности потенциалов между клеткой и средой, происходящее при пороговом и сверхпороговом возбуждении клеток, называется потенциалом действия.
Потенциалы действия обеспечивают проведение возбуждения по нервным волокнам и инициируют процессы сокращения мышечных и секреции железистых клеток.
На основе обобщения большого экспериментального материала было установлено, что потенциалы действия возникают в результате избыточного по сравнению с покоем диффузии ионов натрия из окружающей жидкости внутрь клетки.
Формирование потенциала действия обусловлено двумя ионными потоками через мембрану: поток ионов натрия внутрь клетки приводит к перезарядке мембраны, а противоположно направленный поток ионов калия обусловливает восстановление исходного потенциала покоя. Потоки приблизительно равны по величине, но сдвинуты во времени. Благодаря этому сдвигу во времени и возможно появление потенциала действия.
Потенциал действия, возникнув в одном участке нервной клетки, быстро распространяется по всей ее поверхности. Распространение потенциала действия обусловлено возникновением так называемых локальных токов, циркулирующих между возбужденным и невозбужденным участками клетки.
В состоянии покоя внешняя поверхность клеточной мембраны имеет положительный потенциал, а внутренняя - отрицательный. В момент возбуждения полярность мембраны изменяется на обратную: ее внешняя поверхность заряжена отрицательно по отношению к внутренней (рис.2).
Рис.2. Мембранные потенциалы.
Рис..2. Мембранные потенциалы.
Рис.2. Мембранные потенциалы.
В результате этого между возбужденным и невозбужденным участками мембраны имеется разность потенциалов. Наличие разности потенциалов приводит к появлению между этими участками электрических токов, называемых локальными токами или токами действия. На поверхности клетки локальный ток течет от невозбужденного участка к возбужденному; внутри клетки он течет в обратном направлении. Локальный ток, как и любой электрический ток, оказывает раздражающее действие на соседние невозбужденные участки и вызывает увеличение проницаемости их мембран. Это приводит к снижению в них потенциала покоя. Когда деполяризация достигает критического значения, в этих участках возникают потенциалы действия, а в том участке, который ранее был возбужденным, в это время уже происходят восстановительные процессы реполяризации. Вновь возбужденный участок, в свою очередь, становится электроотрицательным и возникающий локальный ток раздражает следующий за ним участок. Этот процесс многократно повторяется и обусловливает распространение импульсов возбуждения по всей длине клетки в обоих направлениях. В нервной системе прохождение импульсов только в определенном направлении обусловлено наличием синапсов, обладающих односторонней проводимостью.
Под влиянием локальных токов волна возбуждения распространяется вдоль волокна без затухания (бездекрементное проведение). Это обусловлено тем, что локальные токи только деполяризуют мембрану до критического уровня, а потенциалы действия в каждом участке мембраны поддерживаются независимыми ионными потоками, перпендикулярными к направлению распространения возбуждения.
Скорость уменьшения мембранного потенциала до критического уровня зависит от разности потенциалов между возбужденным и невозбужденным участками и от кабельных свойств волокна: электрической емкости и сопротивления мембраны, сопротивлений аксоплазмы и окружающей среды. Кабельные свойства волокна обеспечивают деполяризацию мембраны до критического уровня, а последующая диффузия натрия в клетку усиливает деполяризацию и обеспечивает незатухающее проведение импульса.
Количественно процесс может быть представлен следующим образом. Скорость распространения потенциала действия определяется в основном тем временем, которое необходимо для критической деполяризации мембраны. Обозначим деполяризацию до критического уровня "Dj ". Тогда "Dj ", емкость мембраны "с" и заряд "Dq", необходимый для деполяризации мембраны на "Dj " будут:
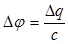
(4)
Заряд мембраны и ее потенциал изменяется вследствие протекания локального тока "I". Тогда время "t", в течение которого заряд мембраны изменяется на Dq, а потенциал на "Dj " будет равно:
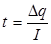
(5)
Из уравнений (4) и (5) получим:
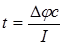
(6)
Величина локального тока "I" по закону Ома определяется разностью потенциалов "U" между возбужденным и невозбужденным участками мембраны и сопротивлением "R" всех участков, по которым протекает локальный ток: 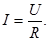
Сопротивление "R" суммируется из сопротивлений всех последовательных участков, по которым протекает локальный ток: мембраны, цитоплазмы, окружающей клетку жидкости.
В основном оно определяется сопротивлением мембраны. Подставляя значение " I " в уравнение (6), получим:
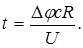
Учитывая, что "Dj " и "U" величины постоянные, можно написать:
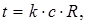
(7)
где k - коэффициент пропорциональности.
Уравнение (7) показывает, что время деполяризации мембраны и, следовательно, время проведения возбуждения пропорциональны произведению "RC", называемому постоянной времени мембраны. Тогда скорость проведения возбуждения будет обратно пропорциональна постоянной времени мембраны.
Скорость проведения импульса возрастает с увеличением диаметра волокна. Это объясняется тем, что с увеличением диаметра уменьшается сопротивление, приходящееся на единицу длины волокна.
В нервных волокнах характер распространения возбуждения зависит от наличия или отсутствия в них миелиновых оболочек. В безмякотных волокнах возбуждение распространяется непрерывно вдоль всей мембраны. Все участки мембраны при этом в свое время становятся возбужденными. В мякотных нервных волокнах возбуждение распространяется несколько по иному. Мякотные нервные волокна имеют толстые миелиновые оболочки, которые через 1 - 3 мм прерывается с образованием так называемых перехватов Ранвье.
 |
Рис.3. Нервные волокна с толстыми миелиновыми оболочками.
В электрическом отношении миелин является изолятором; его удельное сопротивление в 10 млн. раз превышает удельное сопротивление раствора Рингера. В результате этого локальные токи через миелиновые оболочки протекать не могут; они циркулируют между перехватами Ранвье. При возбуждении одного перехвата Ранвье между ним и следующим перехватом возникают локальные токи и импульс как бы перескакивает на второй перехват, со второго - на третий и т.д. Такой способ проведения нервного импульса называется сальтаторным. При блокировании одного перехвата Ранвье каким-либо анестезирующим веществом, например кокаином, импульс сразу передается на третий перехват. При блокировании сразу двух перехватов импульс дальше распространяться не может: сопротивление между первым и четвертым перехватами велико, и локальный ток между ними не достигает порогового значения.
В результате сальтаторного способа передачи скорость распространения первого импульса в мякотных волокнах примерно в 10 раз выше, чем в безмякотных, при одинаковом диаметре волокон. Помимо этого, сальтаторный способ проведения возбуждения является более экономичным, поскольку в этом случае ионные потоки проходят не через всю поверхность клетки, как при непрерывном распространении, а только через поверхность в области перехватов Ранвье.
1.3. Основные характеристики электрического поля. Электрический диполь. Поле диполя. Диполь в электрическом поле.
Электрическим диполем (диполем) называют систему, состоящую из двух равных, но противоположных по знаку точечных электрических зарядов, расположенных на некотором расстоянии друг от друга (плечо диполя).
Основной характеристикой диполя (рис.4) является его электрический, или дипольный момент – вектор, направленный от отрицательного заряда к положительному и равный произведению заряда на плечо диполя:
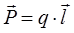
(8)
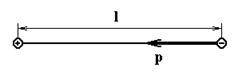
Рис.4. Электрический диполь.
Единицей электрического момента диполя является кулон-метр.
Поместим диполь в однородное электрическое поле напряжённостью 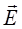
(рис.5). На каждый из зарядов диполя действуют силы 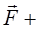
=q
и
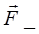
= -q
; эти силы противоположно направлены и создают момент пары сил. Как видно из рисунка, он равен
M=qEl sina=pEsina,(9)
Или в векторной форме
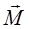
= 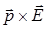
(10)
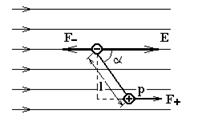
Рис.5. Диполь в однородном электрическом поле.
Таким образом, на диполь в однородном электрическом поле действует вращающий момент, зависящий от электрического момента, ориентации диполя в поле и напряжённости поля.
Рассмотрим теперь диполь в неоднородном электрическом поле. Простоты ради предположим, что диполь расположен вдоль силовой линии. На него действуют силы
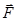
+= 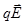
+ и
-=
- ,
где 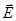
+ и
- -напряжённости поля соответственно в месте нахождения положительного и отрицательного зарядов (на рис.6
->
+). Значение равнодействующей этих сил
F= F_ - F+ = qE_ - qE+ = q (E_ - E+). (11)
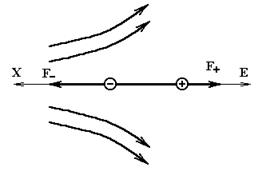
Рис.6. Силы, действующие на диполь.
Введём отношение (E_ - E+)/l, характеризующее среднее изменение напряжённости, приходящееся на единицу длины диполя. Так как обычно плечо невелико, то приближенно можно считать
(Е_ - Е+)/l = dE/dx, (12)
где dE/dx – производная от напряженности электрического поля по направлению оси ОХ, являющаяся мерой неоднородности электрического поля вдоль соответствующего направления. Из (12) следует
Тогда формулу (5) можно представить в виде
F = ql dE/dx = p dE/dx.
Итак, на диполь действует сила, зависящая от его электрического момента и степени неоднородности поля dE/dx.
Если диполь ориентирован в неоднородном электрическом поле не вдоль силовой линии, то на него дополнительно действует еще и вращающий момент. Так что свободный диполь практически всегда будет втягиваться в область больших значений напряженности поля.
Читайте также:
