
Преобразование квантов света в нервных импульсах
С помощью зрительной функции человек способен увидеть происходящие рядом с ним явления, может анализировать разные события, находить отличия одного предмета от другого, а также замечать надвигающуюся угрозу.
Органы зрения устроены таким образом, что различают не только сами объекты, но ещё и цветовое разнообразие живой и неживой природы. Ответственность за это лежит на особых микроскопических клетках — палочках и колбочках, присутствующих в сетчатке глаза. Именно они являются начальным звеном в цепочке по передаче информации об увиденном объекте в затылочную часть головного мозга.
Рецепторы глаз
На сетчатке человека находится приблизительно 115—120 миллионов рецепторов. Это рецепторы в глазу человека, которые помогают воспринимать окружающую реальность. Внешне напоминают продолговатый цилиндр. Они крайне чувствительны к свету, но не могут обеспечить цветовое зрение. Отличаются от колбочек сетчатки глаза, палочки. Они плохо различают цвета и медленно реагируют на передвижения предметов. Состояние этих рецепторов не сказывается на качестве зрения человека. Они находятся на периферии зрения и отвечают за видение в ночное время суток.
Другие зрительные рецепторы в глазах человека называются колбочки. Их приблизительно 7 миллионов, а форма соответствует названию. Как и палочки, колбочки помогают глазу воспринять изображения окружающей среды. Они вместе с палочками преобразовывают нейронные импульсы из лучей света и отправляют их по зрительному нерву в мозг. Колбочки в сетчатке отвечают за восприятие окружающей реальности днем. Именно к цветам чувствительны колбочки сетчатки. Это связано с пигментами, которые находятся в их составе. Расположены колбочки в глазу у человека в области макулы.
Разделяются на 3 типа:
- коротковолновые;
- средневолновые;
- длинноволновые.
Открываясь, чтобы впустить больше света
Ваши зрачки — это черные области перед глазами, которые пропускают свет. Они выглядят черными, потому что свет, который достигает их, поглощается внутри глазного яблока. Затем он преобразуется вашим мозгом в ваше восприятие мира.
Вы, наверное, заметили, что зрачки могут изменить размер в ответ на свет. Снаружи в яркий солнечный день ваши зрачки становятся очень маленькими. Это дает меньше света в глаза, так как есть много доступных.
Когда вы переходите в темное место, ваши зрачки открываются, чтобы стать как можно больше. Это расширение позволяет вашему глазу собирать больше света, чем там есть.
Но от самого маленького размера до самого широкого зрачка ваш зрачок может увеличить свою площадь всего лишь в 16 раз. Вы можете хорошо видеть поперечные изменения уровня освещенности гораздо больше, чем в миллион раз. Так что здесь должно быть что-то еще происходит.
Строение рецепторов
- наружное поле (диск);
- связующую зону;
- внутреннюю;
- базальная зона.
В длину одна палка 0,06 миллиметров, а диаметр — 0,002 мм. Эти фоторецепторы глаза крайне светочувствительны. Они воспринимают максимальное количество волн света, что предоставляет человеку возможность различать предметы в темное время суток. В рецепторах присутствует родопсин или зрительный пурпур, который содержится на мембранных дисках. В желтом пятне палочек практически нет. Под воздействием лучей он раздражается и помогает улавливать свет в ночное время.
Колбочки по строению схожи с палочками:
- наружная зона;
- связующая (перетяжка);
- внутренняя;
- базальная.
Длина рецепторов — 0,05 мм, а диаметр в широкой зоне составляет 0,004 мм. В дисках колбочек содержится йодопсин. Благодаря ему светочувствительные рецепторы обрабатывают поступающее изображение и изменяют его в нейронный импульс. Такая работа обеспечивает дневное видение и более точное изображение реальности. Колбочки улавливают красный и зеленый оттенков. Различают 3 вида йодопсина: эритролаб, хлоролаб цианолаб. Каждый из них отвечает за различие одного из 3-х основных оттенков: синего, красного и зеленого. Но если первые 2 вида были официально найдены учеными, то цианолаб еще не открыт, но уже имеет название.
Существует теория о двухкомпонентном восприятии цветов. Так как цианолаб еще не был найден, то приверженцы этой теории считают, что эритролаб и хлоролаб дают возможность глазу различать красный и зеленый спектры, а синий оттенок глаз улавливает с помощью выцветших родопсин (пигмента палочек). Эту гипотезу подтверждают исследования людей, что не различают синие цвета и плохо ориентируются в темноте.
Сколько времени это займет?
Когда вы находитесь в ярком свете, ваши стержни полностью перегружены и не работают. Если вы выключите свет, ваш зрачок сразу же откроется. Ваши фоторецепторы начинают улучшать свою чувствительность, чтобы впитывать любой свет, который они могут в новых тусклых условиях.
Колбочки делают это быстро – примерно через пять минут их чувствительность повышается. Примерно через 10 минут в темном месте, ваши стержни наконец-то нагоняют и вступают во владение. Вы начнете видеть намного лучше. Примерно через 20 минут ваши удилища сделают все возможное, и вы будете видеть как можно лучше “в темноте.”
Найдите очень темное место, может быть, вашу спальню ночью. Включите любой имеющийся у вас свет и соберите несколько красочных предметов. Потратьте некоторое время, замечая, как красочные, острые и полные контрастные вещи выглядят.
Затем выключите свет и посмотрите, как со временем меняется внешний вид вашей комнаты и предметов. Сначала это будет казаться очень темным; тогда вы быстро увидите лучше благодаря зрачкам и колбочкам, которые делают свою работу. Затем, если он достаточно темный, вы заметите еще одно довольно внезапное улучшение примерно через 10 минут, когда стержни начнут показывать свои вещи. Это называется темная адаптация.
Как насчет полной темноты? Если вы можете найти место, где нет абсолютно никакого света, например, шкаф, ванная или подвал, вы можете попробовать эксперимент снова. На этот раз даже через 20 минут вы не увидите никаких предметов в комнате. Но вы также не увидите полной черноты. Попробуйте и понаблюдайте, что происходит.
Функции рецепторов
Зрительные рецепторы отвечают за качество изображения и за цветное зрение. Светочувствительность у палочек рецепторов сетчатки намного выше, чем у колбочек. При сильном воздействии яркий лучей единственный пигмент родопсин выцветает и воспринимает только короткие волны синего света. Но в темноте он восстанавливается, что дает возможность человеку видеть.
Чувствительность глаз, к предметам, лежащим вне полей зрения, что еще называется конвергенция, выше у тех, у кого наблюдается объединение палочек в группы и соединением с интернейроном, собирающим сигналы c сетчатки.
Следовательно, к функциям палочек и колбочек относится:
- цветовосприятие;
- одновременное распознание нескольких объектов;
- расширение периферического зрения;
- видимость в темноте и сумерках.
География сетчатки
Строение и функции этой оболочки разные в зависимости от местонахождения. В центре расположена круглая зона диаметром около 2 мм, где находится оптический нерв. В этом месте нет светочувствительных рецепторов, это зона слепого пятна.
Левее слепого пятна на 4,5-5 мм находится фовеа или макула – центральная ямка сетчатки или желтое пятно. На самом деле это пятно диаметром до 5 мм, где нет кровеносных сосудов, но расположено максимальное количество световоспринимающих клеток. Центральная ямка – это всего 5 % оптической сетчатки, но именно она отвечает за наибольшую остроту зрения.
Нарушения рецепторов
Дополнительные слои
За слоем с фоторецепторами идут слои, без которых работа всего световоспринимающего аппарата невозможна:
- Наружная пограничная, или мембрана Везхова, разделяет слои друг от друга и необходима для обеспечения трансформации энергии химических связей в нервный импульс.
- Наружный ядерный слой содержит ядра колбочек и палочек.
- Наружный сетчатый слой (плексиформный) образован отростками фоторецепторов и биполярных нейронов.
- Внутренний слой сетчатки содержит ядра биполярных нейронов.
- Во внутреннем ретикулярном слое располагаются клетки, которые ограничивают светочувствительность сетчатки. Именно тут проходит граница между частями сетчатки, где есть сосуды и где их нет. И это последняя ступенька в обработке информации перед направлением ее в мозг.
- Ганглиозный многополярный слой. Наибольшая его толщина в пять рядов клеток в районе центральной ямки сетчатки.
- Волокнистый слой с волокнами зрительного нерва.
- Последний слой – внутренняя мембрана, которая образована нейроглиальными (соединительными) клетками Мюллера и непосредственно прилегает к стекловидному телу.
Механизм восприятия
Палочки работают в изумрудно-зеленой спектральной зоне с длиной волны до 498 нм. Остальные участки воспринимают колбочки, но они чувствительны не только к своим цветам. Длинноволновые и средневолновые рецепторы также реагируют и на другие, просто менее активно. Так как ночью фотонный поток минимален, то распознают его лишь палочки, поэтому человек видит в монохроме и цвета не различает.
При попадании на сетчатку лучей разрушается под действием йодопсина и родопсина. Зрительные пигменты раздражаются и преобразовывают свет в нейронный импульс. Палочки образуют слой нервных волокон. По ним передается импульс от рецепторов в зрительный нерв. Под влиянием света происходит распад пигментов в рецепторах. Их восстановление происходит благодаря белку, который в них содержится. Возобновление белка занимает около 30 минут. Этого времени хватает для полного отображение окружающей среды.
Главные в пироге
Итак, самый наружный слой сетчатки, который непосредственно соприкасается с сосудистой оболочкой глазного яблока и отделен от нее мембраной Бруха, – это пигментный эпителий. Как любой тип эпителия, тут имеются плотно упакованные клетки, шестигранные и организованные в линию. Их особенность – наличие зрительного пурпура. Именно этот пигмент играет важную роль в предохранении фоторецепторов от рассеивания и потерь, бликов и переотражения света. Пигментный слой сетчатки выполняет функции ввода питательных веществ и отвода продуктов метаболизма от всех остальных частей этого пирога, и обеспечивает гемато-ретинальный барьер.
К клеткам пигментного эпителия прилегают светочувствительные клетки – колбочки и палочки. На их строении остановимся чуть подробнее, а главная функция этого слоя сетчатки – преобразование фотонов света в нервные импульсы. Или преобразование энергии световых волн в электрические сигналы.
Когда стоит начать беспокоиться
Симптоматика патологий сетчатки не специфическая, и часто пациент долгое время не подозревает о существующей проблеме. Записаться к офтальмологу на обследование следует, если:
- Появилось ощущение снижения общей остроты зрения.
- Появляются вспышки, блики или молнии перед глазами.
- Если поле зрения сузилось.
- Появляются круги или темные пятна перед глазами.
Офтальмолог после осмотра назначит дополнительное обследование, которое включает офтальмоскопию, УЗИ глаз, флуоресцентную ангиографию, оптическую когерентную томографию. После чего может быть поставлен диагноз и начато лечение.

Давление света (или давление электромагнитного излучения) это механическое давление, оказываемое на любую поверхность в результате обмена импульсом между объектом и электромагнитным полем.
Первооткрывателем этого понятия является Иоганн Кеплер (1571-1630). В 1619 году, наблюдая за кометой, он отметил, что ее хвост всегда направлен в сторону от Солнца.
Спустя более двухсот лет в 1862 году Джеймс Максвелл (1831-1879) предположил, что свет как электромагнитное излучение обладает свойствами импульса и, следовательно, оказывает давление на любую поверхность, с которой контактирует. Экспериментально это было подтверждено лишь в 1900 году Петром Лебедевым.

Иоганн Кеплер, Петр Лебедев и Арнольд Зоммерфельд.
Если суммировать все воедино, то мы получим следующее: частицы света (фотоны) ударяются об атомы тела и передают ему часть своего импульса, а тело от этого становится быстрее.
Пока все логично. Однако не все так просто. Ранее проводились опыты, в которых фотоны определенной длины волны выбивали из атомов отдельные электроны. Импульс этих электронов был больше, чем у фотона, который с ним взаимодействовал. Это невозможно, скажете вы, ибо есть второй закон Ньютона, в котором говорится, что на любое действие имеется противоположное равное противодействие (утрированно говоря). Тем не менее, в 1930 году немецкий ученый Арнольд Зоммерфельд предположил, что дополнительный импульс выброшенного электрона происходит из атома, который он покинул. Получается, что движение атома должно быть направлено в сторону источника фотонов, т.е. к свету. Теория весьма смелая, но в те годы подтвердить ее на практике было нереально ввиду отсутствия необходимых технологий.
И вот 90 лет спустя наши современники смогли впервые в мире воочию понаблюдать этой таинственный процесс.
Авторы труда напоминают, что вектор электрического поля электромагнитной волны ориентирован перпендикулярно оси распространения света. Поскольку это поле управляет фотоионизацией*, стоит предположить, что его направление будет осью симметрии для угловых распределений фотоэлектронов и фотоионов.
Фотоионизация* — ионизация молекулы/атома непосредственно при абсорбции фотонов, энергия которых равна или больше энергии ионизации.
Фотоэффект — процесс взаимодействия электромагнитного излучения и вещества, когда энергия фотонов передается электронам вещества.
Фотоэлектрон — электроны, вытесняемые из вещества, когда на него воздействует электромагнитное излучение.
Фотоион — катион (положительно заряженный ион), полученный в результате фотоионизации.
Однако при высоких энергиях фотонов Eγ и соответствующих высоких фотонных импульсах kγ эта симметрия нарушается, а импульсные распределения фрагментов реакции асимметричны относительно направления распространения света.
Зоммерфельд в своих изысканиях понял, что средний прямой импульс электронов, превышающий импульс фотона (⟨k e x⟩ > kγ), влечет за собой то, что средний импульс фотоиона должен быть противоположным для учета сохранения импульса.
Стоит также отметить, что так называемые недипольные эффекты, возникающие из-за ненулевого импульса фотона, также оказывают существенное влияние на однофотонную ионизация. Кроме того, более высокие мультипольные компоненты взаимодействия света и вещества не только изменяют угловое распределение фотоэлектронов, но также открывают дополнительные пути ионизации, которые запрещены диполями.
В данном исследовании эксперименты по однофотонной ионизации были выполнены в двух вариантах:
- высокоэнергетический (300–1775 эВ) на PETRA III (DESY/Немецкий электронный синхротрон) с применением света с циркулярной поляризацией;
- низкоэнергетический (12–40 кэВ) на ID31 (European Synchrotron Radiation Facility) с применением света с линейной поляризацией.
Для измерений состояния заряда и трехмерного вектора импульса фотоионов был использован метод спектроскопии COLTRIMS (Cold Target Recoil Ion Momentum Spectroscopy ).
Пучок фотонов проходил под прямым углом со сверхзвуковой газовой струей He (низкоэнергетический эксперимент) или N2 (высокоэнергетический эксперимент).
Фотонный пучок был пересечен под прямым углом сверхзвуковой газовой струей He (низкоэнергетический эксперимент) или N2 (высокоэнергетический эксперимент). Ионы направлялись электрическим полем к чувствительному ко времени и положению детектору с отсчетом положения линии задержки*.
Линия задержки* — устройство задержки электрических и электромагнитных сигналов на заданный промежуток времен.
Начальные импульсы после фотоионизации были получены от времени полета ионов и положения точки контакта. В экспериментах с N2 рассматривалась исключительно ионизация K-оболочки (электронная оболочка атома первого уровня) с последующим распадом Оже*.
Эффект Оже* — выход электрона из атомной оболочки ввиду безызлучательного перехода в атоме при снятии возбуждения.
В таком случае возникает два однозарядных иона, которые совпадают с оже-электроном. Из этих трех векторов импульса был рассчитан импульс иона N2 + в момент после фотоэлектронной эмиссии.
Чтобы получить доступ к ионным импульсам в абсолютном масштабе, важно точно знать местоположение ионов с нулевым импульсом на нашем детекторе. Для данных высоких энергий эта нулевая точка получается из ионов, которые создаются комптоновским рассеянием*.
Комптоновское рассеяние* — некогерентное (фотоны до и после рассеяния не интерферируют) рассеяние фотонов на свободных электронах.
В этом случае импульс фотона передается электрону, и поэтому ион остается с распределением импульса, центрированным в начальной точке.
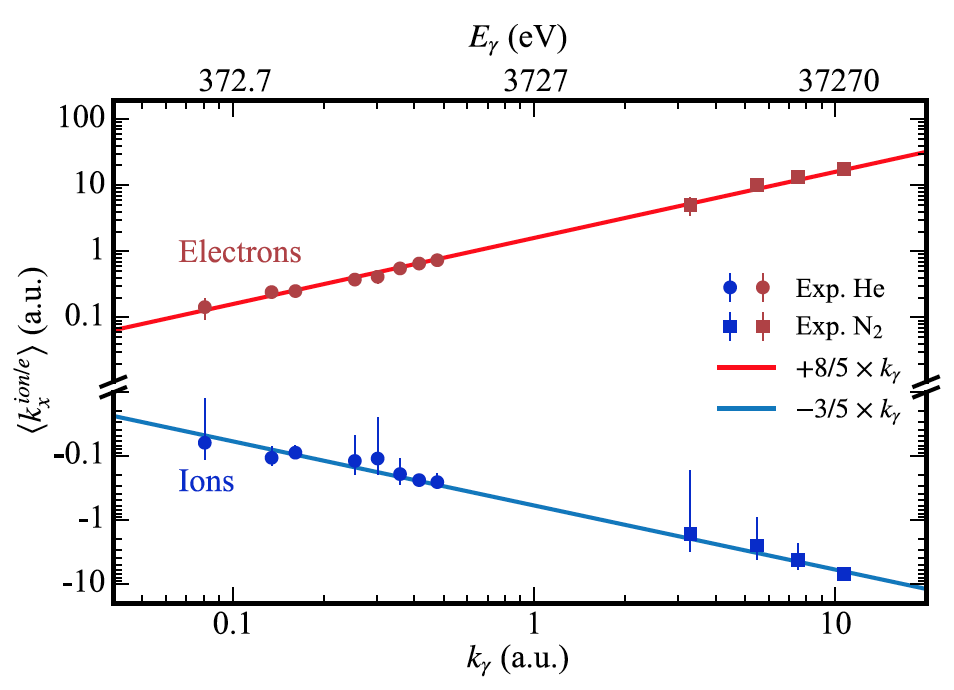
Изображение №1
На графике выше суммированы результаты исследования. Синим цветом показано измеренное среднее значение импульса иона в направлении распространения света ⟨k ion x⟩ как функция энергии фотона (верхняя шкала) или импульса фотона (нижняя шкала). Точки (низкие энергии фотонов) соответствуют однократной ионизации He, а квадраты (высокие энергии фотонов) — ионизации K-оболочки N2.
Отрицательные значения соответствуют обратному излучению, то есть в противоположную сторону от направления распространения фотона. Красным цветом обозначено среднее значение импульса фотоэлектрона ⟨k e x⟩, полученное за счет измеренного импульса иона с учетом сохранения импульса.
Красная и синяя линии демонстрируют прогнозируемые данные в соответствии со следующими формулами:
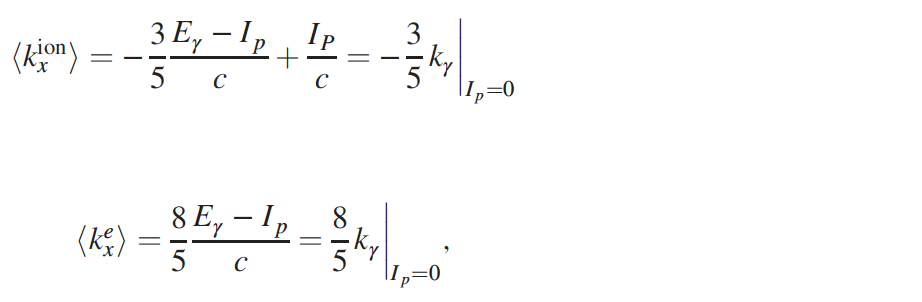
где Ip — потенциал ионизации; с — скорость света.
Из вышеописанных данных следует, что это является прямым практическим доказательством теории касательно обратно направленной эмиссии ионов при фотоионизации.
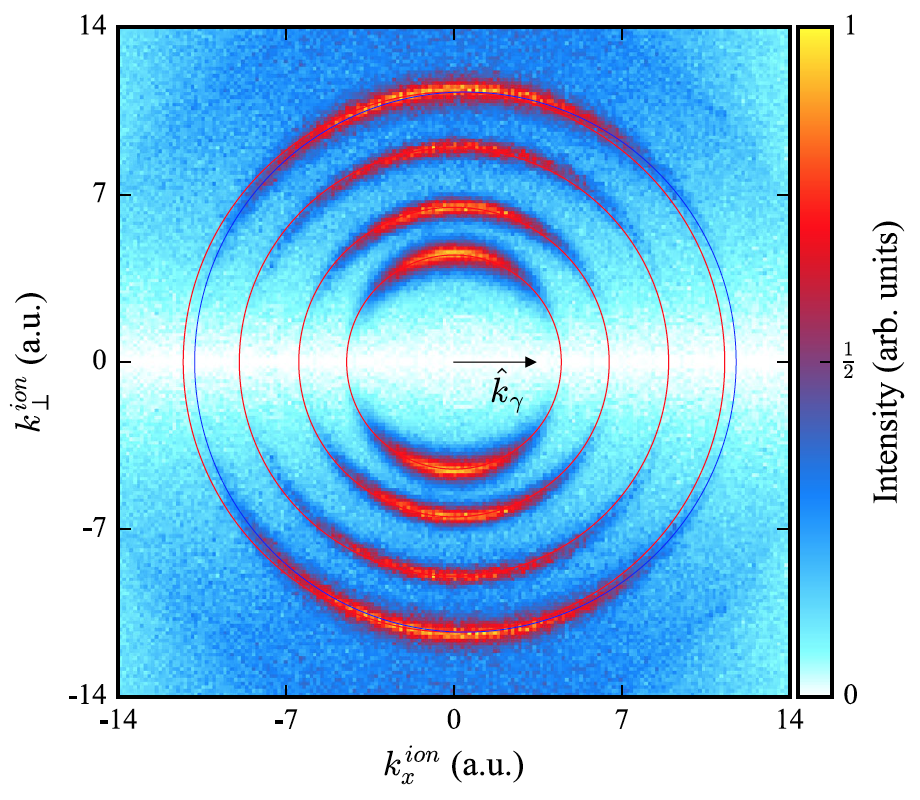
Изображение №2
Изображение выше демонстрирует нам распределение фотоионного импульса для фотоионизации He, где использовались фотоны с циркулярной поляризацией в 300, 600, 1125 и 1775 эВ. Горизонтальная ось — составляющая импульса, параллельная kγ, а вертикальная ось это импульс, перпендикулярный оси фотона. Красным отмечены концентрические кольца, центр которых расположен там же, где и начальная точка импульсного пространства. Радиус колец равен соответствующим фотоэлектронным импульсам ke = √ 2(Eγ — Ip).
События ионизации не накапливаются на этих кольцах, а смещаются вперед в направлении распространения фотонов. Это наиболее четко видно на внешнем кольце, соответствующем энергии фотона 1775 эВ. При этом синие кольца смещаются вперед фотонным импульсом 1775 эВ фотона.
Следовательно, измеренные распределения импульса иона непосредственно показывают, что импульс фотона в основном поглощается ионом, что является следствием сохранения импульса.
В каждом отдельном событии ионизации импульс фотона передается центру масс системы, который почти совпадает с ионом. Соответствующее импульсное распределение электрона показывает окружность того же радиуса, но не смещенную вперед.
Помимо смещения вперед кольца в импульсном пространстве ионов, распределение импульсов на этом кольце также изменяется в зависимости от энергии фотона. Это распределение больше отклоняется в обратное полушарие при увеличении Eγ.
Сохранение импульса требует, чтобы конечный импульс измеряемого иона равнялся импульсу фотона за вычетом импульса фотоэлектрона. Таким образом, распределение ионов на смещенной сфере в импульсном пространстве и угловое распределение фотоэлектронов в лабораторной системе отсчета являются прямыми зеркальными отражениями друг друга (изображение №3).
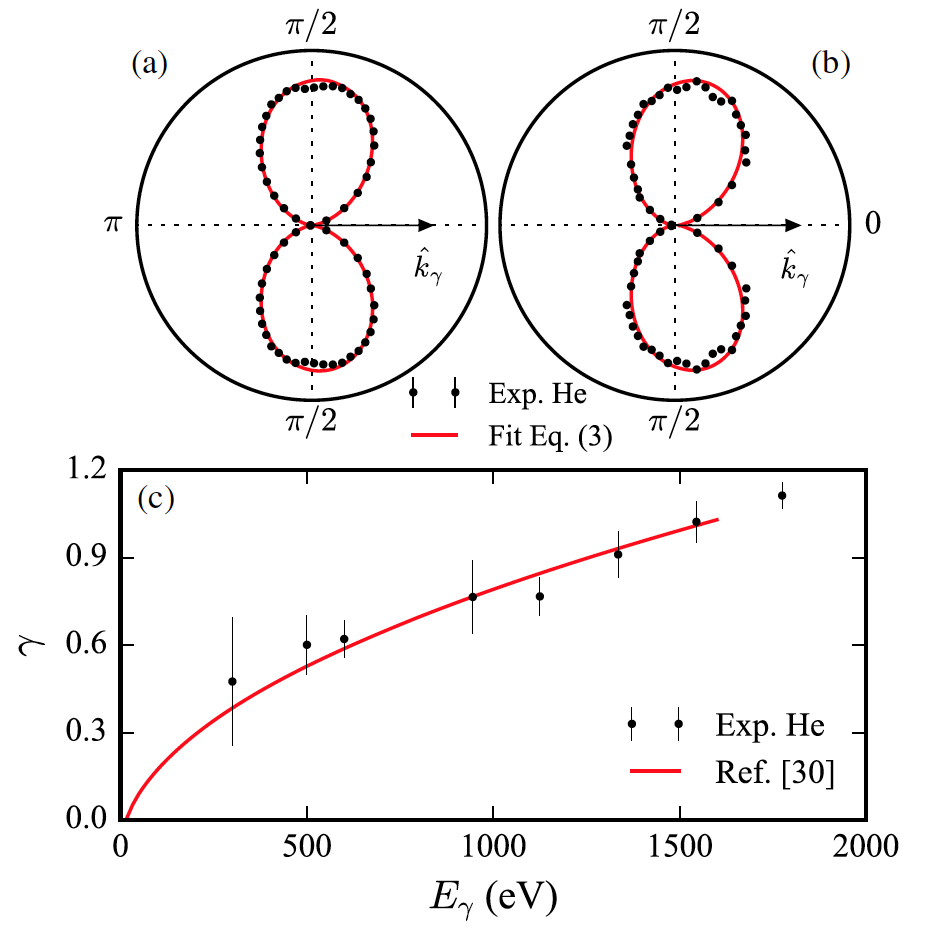
Изображение №3
Они имеют приблизительную дипольную форму, поскольку начальное состояние является He(1s), и, таким образом, главная составляющая углового момента (момента импульса) в конечном состоянии представляет собой диполь. Кроме того, эта дипольная форма отклонена вперед.
Чтобы было проще понять все нюансы, ученые предлагают вспомнить, как именно происходит передача импульса фотона при взаимодействии с электромагнитным полем. Для простоты примера была выбрана фотоионизация 1s-электрона атома водорода.
Вводя координату RH для центра масс атома и координату r′ для электрона 1s по отношению к RH, абсолютная координата электрона 1s в лабораторной системе отсчета может быть переписана как r = RH + r′. Таким образом, соответствующая фаза может быть выражена следующим образом: e ikγ·r = e ikγ·RH e ikγ·r′ .
Эта фаза, представленная полем, модифицирует элемент матрицы перехода: первый фактор из уравнения выше входит в элемент матрицы перехода ⟨π|e ikγ·RH |π 0⟩ между переходными состояниями атомного центра масс, которые описываются плоскими волнами (2π) −3/2 e iπ·RH с импульсом π. Эта амплитуда порождает закон сохранения импульса π = π0+kγ. Таким образом, поглощение фотона атомом привносит в его центр массы импульс kγ.
Второй фазовый фактор e ikγ·r′ из уравнения отвечает за мультипольные правки за пределами электрического дипольного приближения.
Выше порога ионизации в каждом событии ионизации ион получает импульс фотона и, кроме того, отдачу от фотоэлектрона. Дополнительная передача углового момента орбиты от фотона приводит к смещению вперед углового распределения электрона. Этот направленный вперед средний импульс электрона уравновешивается обратно направленной передачей импульса иону.
По результатам исследования видно, что для s-начальных состояний обратный импульс иона масштабируется -(3/5)kγ, подтверждая теорию, описанную Зоммерфельдом.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых .
Если фотон имеет низкую энергию, то при теоретическом моделировании его импульсом можно пренебрегать, говорят ученые. Однако при высоких энергиях фотона подобное пренебрежение приводит к значительным неточностям. Экспериментальные данные позволили определить порог, когда импульс фотона больше нельзя не учитывать.
В дальнейшем ученые намерены продолжить начатую работу, поскольку совершенные открытия открывают двери перед более детальным рассмотрением процессов, происходящих в момент распределения энергии между двумя или более фотонами.
Благодарю за внимание, оставайтесь любопытствующими и хорошей всем рабочей недели, ребята. 🙂
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99 , уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
17 октября 2017
- 869
- 0,7
- 2
- 3
Представление нейрона, который можно запускать и выключать с помощью света.
![]()
Виталий Шевченко

![]()
Антон Чугунов![]()
Андрей Панов






Как управлять мозгом с помощью света?
Оптогенетика — это способ управления определенными клетками с помощью света. Чаще всего речь идет о нейронах и мышечных клетках — так называемых электровозбудимых клетках. Как это работает? Для нормального существования клетка старается поддерживать определенные условия внутри и снаружи себя. По разные стороны мембраны клетки находятся ионы, но в различных концентрациях. В частности, внутри клетки преобладают ионы калия, а снаружи — ионы натрия и хлора. Причем, соотношение заряженных ионов таково, что клетка внутри заряжена более отрицательно (имеет отрицательный мембранный потенциал [2], равный примерно −70 мВ). Теперь, если мы каким-либо образом извне направим положительные заряды внутрь клетки, произойдет ее деполяризация, и клетка возбудится (возникнет потенциал действия). У нейронов в мембране находятся свои белки, которые реагируют на изменение потенциала. Именно они отвечают за распространение нервного импульса по цепочке нейронов.
В природе есть различные организмы, жизнь которых зависит от наличия света. Поглощая фотоны, естественные белки в этих организмах могут переносить различные ионы через мембрану клеток (например, протоны или ионы натрия). Если в нейрон или мышечную клетку внедрить такой белок, переносящий положительные ионы внутрь, и посветить на эту клетку, то мы искусственно запустим нервный импульс. Мы также можем и заблокировать прохождение импульса: для этого нужно, напротив, еще сильнее уменьшить мембранный потенциал — так, чтобы собственные клеточные белки не справились с повышением потенциала до нужного порога. Такое уменьшение называется гиперполяризацией. Этого можно добиться, выкачивая положительные ионы из клетки или закачивая в нее отрицательные.
Обобщая, можно сказать, что оптогенетика находит белки-транспортёры, реагирующие на свет, пытается доставить эти белки в нужные клетки и создает решения, как же посветить на эти клетки светом (это особенно сложно, когда хочется светить на клетки глубоко в мозге). Причем, это система модульная, ее составные компоненты можно заменять.
Скальпель, зажим, родопсин
Классические оптогенетические инструменты — галородопсин, археародопсин и канальный родопсин (рис. 1) [5]. Галородопсин и археародопсин используются для деактивации нейронов, они блокируют передачу нервного импульса. Под действием света галородопсин переносит отрицательно заряженные ионы хлора внутрь клетки, тем самым вызывая гиперполяризацию нейрона. Археародопсин выкачивает протоны (положительные заряды) из клетки, это, естественно, также гиперполяризует нейрон. И галородопсин, и археародопсин — это так называемые насосы, помпы. Если вокруг есть нужный им ион (а он есть почти всегда), то, поглотив квант света, эти белки активно перенесут ион с одной стороны мембраны на другую.
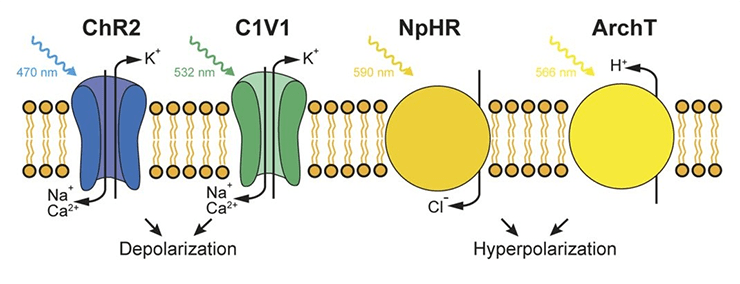
Рисунок 1. Принципы работы классических оптогенетических инструментов: канального родопсина, галородопсина и протонного насоса (археародопсина). Перенос ионов натрия внутрь клетки, обусловленный канальным родопсином 2, приводит к деполяризации, а перенос ионов хлора галородопсином или протонов археародопсином приводят к гиперполяризации клетки.
Канальный родопсин в настоящее время широко используют для активации электровозбудимых клеток. Он долгое время оставался единственным белком, способным на это. Канальный родопсин позволяет положительным ионам перетекать из области с их большей концентрацией в область с меньшей. Из внеклеточного пространства внутрь клетки идет поток ионов натрия, и нейрон возбуждается.
Несмотря на то, что три классических белка-транспортёра в принципе решают задачи активации и деактивации электровозбудимых клеток, у каждого из них есть свои преимущества и недостатки. Например, галородопсин с недостаточно высокой скоростью переносит ионы хлора, а археародопсин в своей работе использует протоны и меняет кислотность среды вокруг нейрона, отчего клетка может начать чувствовать себя неважно. Канальный родопсин — замечательный инструмент, но переносит ионы только пассивно: ему нужна разность концентраций ионов, чтобы все работало. Кроме этого, канальный родопсин не избирателен к ионам и эффективен, только если его освещать синим или фиолетовым светом, а такое коротковолноое излучение очень неглубоко проникает в ткани (рис. 2). Это ограничивает возможность применения белка, если интересующий нас участок мозга или мышцы находится глубоко.
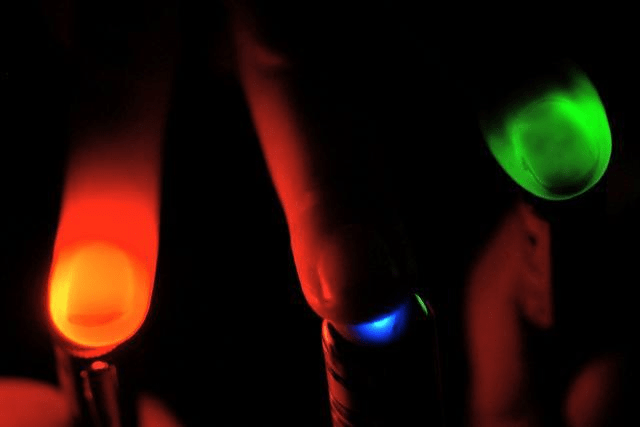
Рисунок 2. В практических применениях оптогенетики важно иметь возможность облучать клетки в глубине тканей. На рисунке видно, что красный свет может просветить ткани глубоко, в то время как синий свет не может проникнуть даже на толщину пальца.
Оптогенетические инструменты будущего
Именно поэтому не прекращается поиск, разработка и оптимизация оптогенетических инструментов нового поколения. В последнее время существенно изменили и подстроили параметры работы популярного канального родопсина: с помощью направленного введения мутаций можно получить его аналоги, более быстрые и специфичные к ионам кальция, с оптимумом работы, сдвинутым в более длинноволновую часть спектра.
Исследование баз данных белков и организмов позволяет отыскивать новые белки-транспортёры, пригодные для нужд оптогенетики. Каждая из этих находок ценна по-своему. Открытые анионные канальные родопсины [6] позволяют деактивировать нейроны, создавая поток отрицательных зарядов внутрь клетки. Химерный белок, состоящий из фоточувствительной части и вирусного калиевого канала [7], может деактивировать нейроны, пропуская ионы калия из цитоплазмы во внеклеточное пространство.
Мы с коллегами из лаборатории перспективных исследований мембранных белков МФТИ [8] внесли свой вклад в открытие и рациональный инжиниринг новых оптогенетических инструментов.
В 2013 году японские ученые открыли натриевый насос KR2 [9], выкачивающие натрий из клетки. Нам с коллегами в 2015 году удалось закристаллизовать белок и решить его структуру в различных функциональных состояниях. Полученные результаты мы опубликовали в журнале Nature Structural and Molecular Biology [10]. Анализируя структуры (рис. 3), мы разобрались в механизмах и условиях работы белка KR2. Оказалось, что у него есть полость, отвечающая за избирательность иона, который будет перенесен. И внеся в белок мутации, затрагивающие эту полость, нам удалось получить модификацию белка, которая прокачивает ионы калия [9], [10]. А ведь именно на паре ионов натрия/калия работают собственные клеточные белки, ответственные за прохождение нервного импульса. Таким образом, на основе полученных данных мы смогли создать более естественный, физиологический оптогенетический инструмент. Хотелось бы отметить, что это пример того, как человек, используя свои знания и воображение, может подстраивать под собственные нужды то, что придумала природа.
Наша группа включает в себя тех, кто может произвести белок в большом количестве, кто имеет большой опыт в кристаллизации мембранных белков (а это очень непростая задача!) и кто умеет решить структуру белка, получив данные рентгеновской дифракции. Затем мы анализируем полученную информацию и проводим дальнейшие эксперименты с модификациями белка. Новые функции мутантных форм исследуем электрофизиологически, методом локальной фиксации потенциала [1]. Финальным аккордом мы стараемся продемонстрировать применимость изученного белка в нервных клетках.
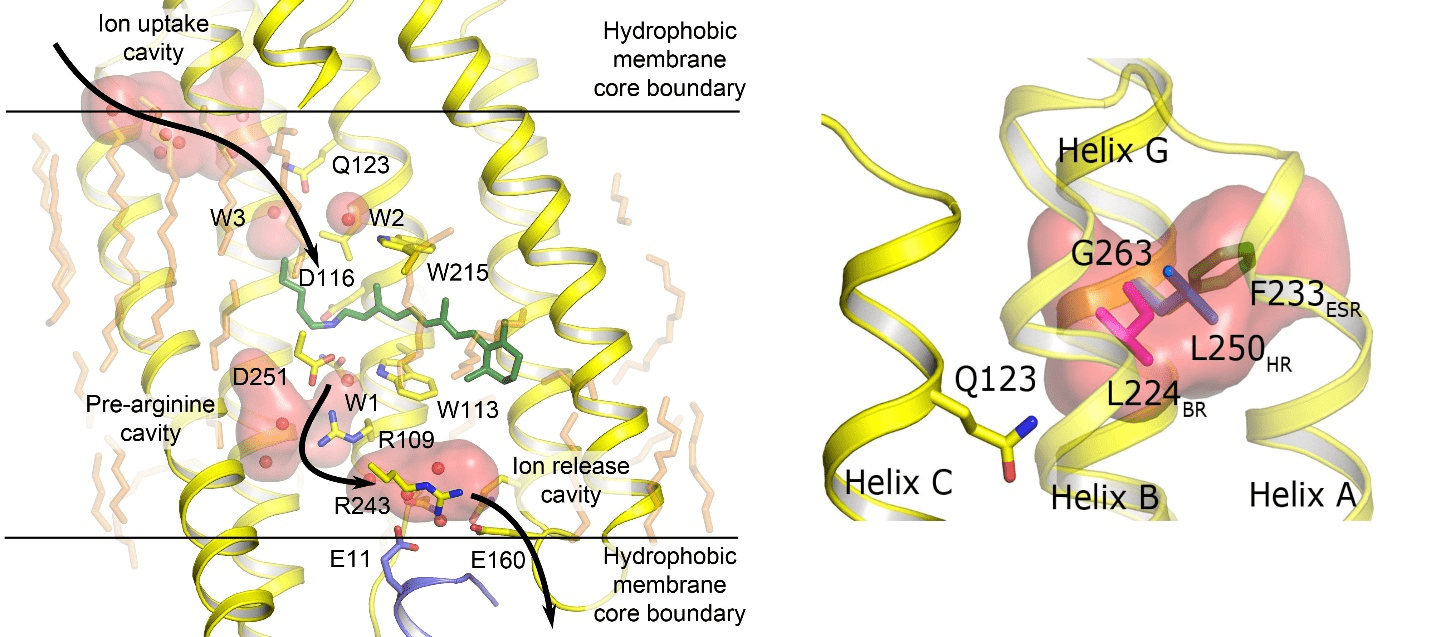
Рисунок 3. Решенная структура натриевого насоса KR2. Слева показана структура KR2, стрелками указан путь иона натрия при прокачивании. Слева сверху на структуре показана полость, где захватывается ион. Полость исполняет роль селективного фильтра. Справа эта полость показана крупно. Разными цветами показаны перепробованные нами мутации, чтобы превратить натриевый насос в калиевый.
Чтобы увидеть рисунок в полном размере, нажмите на него.
Наконец, буквально две недели назад мы опубликовали статью в журнале Science Advances [11], где фактически брошен вызов монополисту — канальному родопсину. Мы открыли протонный насос, переносящий протоны внутрь клетки. Чтобы его найти, мы исследовали широкое множество родопсинов — таких, чья функция была еще неизвестна. Буквально, мы изучали разные микроорганизмы, старались найти те, которые живут в каких-то экзотических условиях. Мы перебрали с десяток кандидатов, прежде чем нашли тот, который давал бы надежду на дальнейшее практическое применение. Ксенородопсины — так называется семейство таких насосов — обнаружены в крошечных археях (наногалоархеях), которые обитают в щелочных водоемах. Мы считаем, что эти организмы используют такой белок для поддержания гомеостаза, преодолевая негативное действие окружающей среды. Ксенородопсины эффективно работают, поглощая красный свет, что позволяет использовать их глубоко в тканях. Поэтому мы смело утверждаем, что открытые белки могут составить достойную конкуренцию канальному родопсину (рис. 4).

Рисунок 4. У исследователей, которым нужно активировать нервные клетки, теперь есть выбор: ксенородопсин (оптимум активности в красном диапазоне) или канальный родопсин (оптимум — в синем диапазоне).
Ксенородопсины переносят протон достаточно быстро, и уже через 2,5 миллисекунды готовы перенести следующий протон. Фактически, это означает, что белок может возбуждать человеческие нейроны с максимальной скоростью. Ксенородопсины способны занять нишу работы с мышечными клетками. Дело в том, что мышечные клетки очень чувствительны к транспорту кальция. Поэтому, при работе с мышцами хотелось бы избежать переноса кальция, чего не может обеспечить канальный родопсин, потому что он неселективен (рис. 1). А ксенородопсины могут.
Как и в случае с натриевым насосом, нам удалось решить структуру ксенородопсина (рис. 5, слева) и понять механизм переноса протона. Многие исследователи, которые работают в области фотоактивных белков, были очень удивлены тому, что такой белок мог найтись в природе. Широко распространено мнение, что перенос протонов через мембрану подобных микроорганизмов в первую очередь обеспечивает клетку энергией. А поведение ксенородопосина не вписывалось в эту картину. Поэтому мы специально показывали на модельных клетках HEK293, что есть ток внутрь (рис. 5, справа сверху). Причем направление тока оставалось тем же, даже если к клетке приложить удерживающее напряжение. Ксенородопсины способны насильно вкачивать протоны в цитоплазму. В этой же работе мы поставили восклицательный знак: на деле показав, что белок может возбуждать нейроны с высокой частотой (рис. 5, справа снизу).
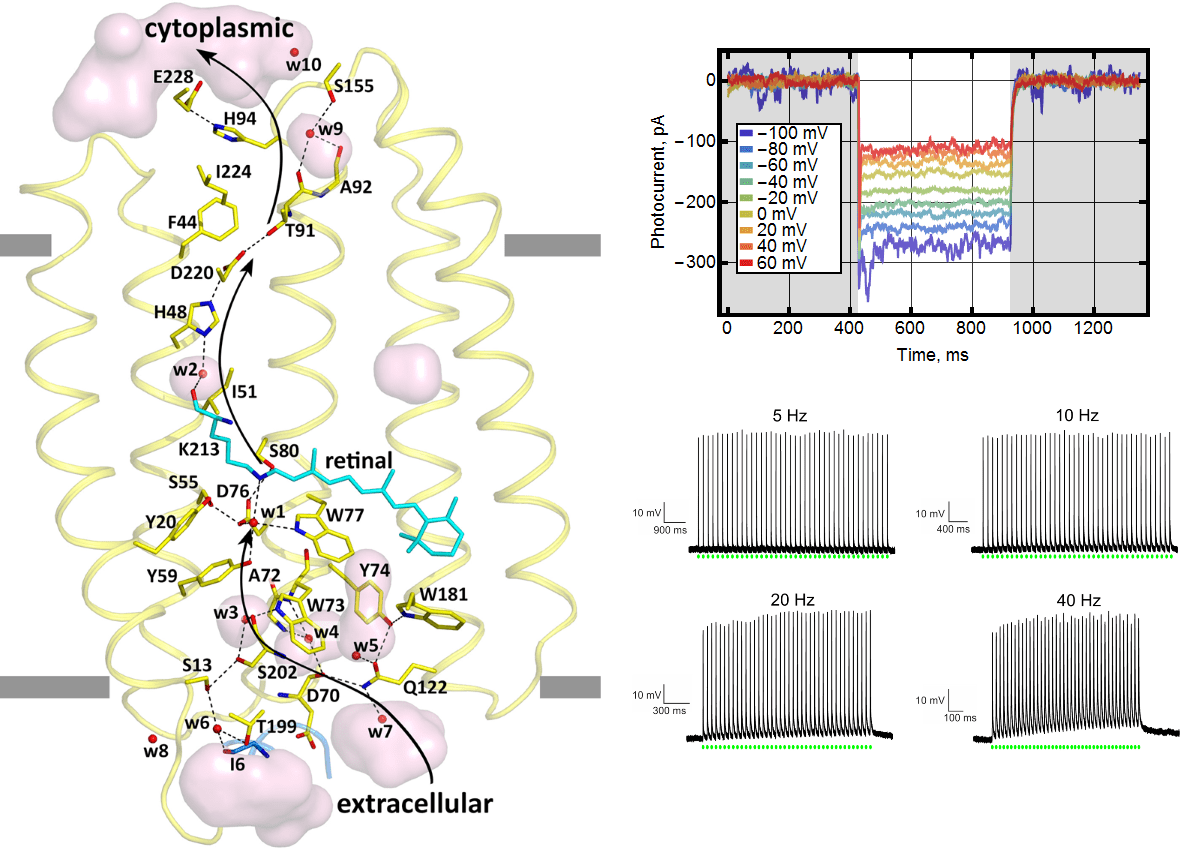
Рисунок 5. Ксенородопсин и его работа. Слева показана кристаллическая структура обратного протонного насоса — ксенородопсина. Справа сверху — наличие тока внутрь клеток HEK293 при различных напряжениях, приложенных к клетке. Справа снизу — возбуждаемые потенциалы действия при облучении нейронов световыми импульсами, повторяющимися с разной частотой.
Чтобы увидеть рисунок в полном размере, нажмите на него.
В заключение
Исследования мозга и минимально инвазивная терапия заболеваний, связанных с нервной системой, продолжают и продолжат быть актуальными в течение XXI века. Оптогенетика и ее инструменты — молекулярные машины и белки-транспортёры — дадут возможность улучшить качество жизни людей с перманентными болевыми синдромами, нейродегенеративными возрастными заболеваниями, помогут частично восстановить зрение и слух [12], [13], а возможно, и мышечные дисфункции. Фундаментальные исследования, поиск и рациональная модификация белков будут лежать в основе успеха оптогенетических подходов в терапии.
Читайте также:
