
Преобразование стимула в нервный импульс в рецепторе
Рецепцией называют процесс восприятия и трансформации (преобразования) энергии внешнего раздражителя в энергию нервного импульса или в сложную последовательность внутриклеточных процессов.
Под клеточной рецепцией понимают процесс восприятия и преобразования химического сигнала в сложную последовательность внутриклеточных химических процессов. Клеточная рецепция обеспечивает возможность обмена информации между клетками, который осуществляется при помощи биологически активных веществ (гормонов, медиаторов). Обязательным этапом такого межклеточного взаимодействия является связывание молекул вещества с соответствующей молекулой клетки-мишени, называемой клеточным рецептором . Роль клеточных рецепторов играют специфические белковые молекулы, которые могут быть расположены на поверхности клетки, в цитоплазме или в ядре. Механизмы рецепции медиаторов и гормонов подробно рассматриваются в соответствующих лекциях.
Сенсорной рецепцией называют процесс восприятия и преобразования энергии раздражителей внешней и внутренней среды организма в энергию нервных импульсов, передаваемую по чувствительным нервам в ЦНС. Сенсорный рецептор представляет собой нервную клетку или комплекс нервной и эпителиальной клетки, специально приспособленный для восприятия определенного типа раздражителей. Сенсорные рецепторы являются начальными звеньями любой рефлекторной дуги, а также участвуют в оценке параметров полезного приспособительного результата в функциональных системах организма.
По строению рецепторы подразделяют на первичные и вторичные (рис. 1).
К первичным относят такие сенсорные рецепторы, у которых действие раздражителя воспринимается непосредственно периферическими отростками чувствительного нейрона (нервными окончаниями), которые могут быть:
свободными, т. е. не имеют дополнительных образований;
инкапсулированными, т.е. окончания чувствительного нейрона заключены в особые образования, осуществляющие первичное преобразование энергии раздражителя.
К вторичным относят такие сенсорные рецепторы, у которых действие раздражителя воспринимается специализированной рецептирующей клеткой не нервного происхождения. Возбуждение, возникшее в рецептирующей клетке, передается через синапс на чувствительный нейрон.
Рис. 1. Первичные и вторичные рецепторы.
1 – тело чувствительного нейрона;
2 – периферический отросток чувствительного нейрона (дендрит);
3 – центральный отросток чувствительного нейрона (аксон);
4 – глиальная капсула;
5 – рецептирующая клетка;
6 – синапс между рецептирующей клеткой и чувствительным нейроном.
Тело чувствительного нейрона обычно располагается за пределами ЦНС: в спинномозговом или вегетативном ганглии. От такого нейрона отходят два отростка – дендрит, который следует к периферическим органам и тканям, и аксон, который направляется в спинной мозг.
По расположению сенсорные рецепторы подразделяют на:
экстерорецепторы – воспринимают раздражители из внешней среды организма;
интерорецепторы – воспринимают раздражители из внутренней среды организма;
проприорецепторы – специализированные рецепторы опорнодвигательной системы.
По разнообразию воспринимаемых раздражителей сенсорные рецепторы подразделяют на:
мономодальные – приспособлены для восприятия только одного вида раздражителя;
полимодальные – приспособлены для восприятия различных видов раздражителей.
По модальности сенсорные рецепторы подразделяют на:
хеморецепторы – воспринимают действие химических веществ;
фоторецепторы – воспринимают световые раздражители;
механорецепторы – воспринимают давление, вибрацию, перемещение, степень растяжения;
терморецепторы – чувствительны к изменениям температуры;
ноцицепторы – воспринимают болевое раздражение.
Действие раздражителя. Внешний стимул взаимодействует со специфическими мембранными структурами окончаний чувствительного нейрона (в первичном рецепторе) или рецептирующей клетке (во вторичном рецепторе), что приводит к изменению ионной проницаемости мембраны.
Генерация рецепторного потенциала. В результате изменения ионной проницаемости происходит изменение мембранного потенциала (деполяризация или гиперполяризация) чувствительного нейрона (в первичном рецепторе) или рецептирующей клетке (во вторичном рецепторе). Изменение мембранного потенциала, наступающее в результате действия раздражителя, называют рецепторным потенциалом (РП) .
Распространение рецепторного потенциала. В первичном рецепторе РП распространяется электротонически и достигает ближайшего перехвата Ранвье. Во вторичном рецепторе РП электротонически распространяется по мембране рецептирующей клетки и достигает пресинаптической мембраны, где вызывает выделение медиатора. В результате срабатывания синапса (между рецептирующей клеткой и чувствительным нейроном) происходит деполяризация постсинаптической мембраны чувствительного нейрона (ВПСП). Образовавшийся ВПСП распространяется электротонически по дендриту чувствительного нейрона и достигает ближайшего перехвата Ранвье.
В области перехвата Ранвье РП (в первичном рецепторе) или ВПСП (во вторичном рецепторе) преобразуется в серию ПД (нервных импульсов). Образовавшиеся нервные импульсы проводятся по аксону (центральному отростку) чувствительного нейрона в ЦНС. Поскольку РП генерирует образование серии ПД, его часто называют генераторным потенциалом .
чем выше сила действующего раздражителя, тем больше амплитуда РП;
чем больше амплитуда РП, тем больше частота нервных импульсов.
Чувствительность. Количественной мерой чувствительности сенсорного рецептора является абсолютный порог чувствительности – минимальная сила раздражителя, способная вызвать возбуждение рецептора.
Адаптацией называют явление ослабления возбуждения в рецепторе при действии длительного раздражителя постоянной силы.
В зависимости от скорости адаптации рецепторы подразделяют на:
тонические (пропорциональные) рецепторы генерируют нервные импульсы в течение всего времени действия раздражителя; после высокочастотного залпа в начале действия раздражителя частота нервных импульсов устанавливается на постоянном уровне (рис. 3, А);
промежуточные (фазнотонические) рецепторы генерируют нервные импульсы в течение всего времени действия раздражителя, однако их частота существенно уменьшается (рис. 3, Б);
фазные (дифференциальные) рецепторы генерируют нервные импульсы в начальный (ON-ответ) и конечный (OFF-ответ) период действия раздражителя (рис. 4, В).
Рис. 3. Адаптация тонических ( А ), промежуточных ( Б ) и фазных рецепторов ( В ) к длительно действующему раздражителю постоянной силы.
Рецептивным полем нейрона называют множество рецепторов, функционально связанных с этим нейроном. Рецептивное поле нейрона представляет собой динамическое образование – один и тот же нейрон в различные моменты времени может оказаться функционально связанным с различным количеством рецепторов. Максимальная величина рецептивного поля какого-либо нейрона соответствует количеству рецепторов, которые связаны с эти нейроном морфологически, а минимальная величина может ограничиваться всего одним рецептором (рис. 4).
Рис. 4. Рецептивные поля нейронов.
А – максимальное рецептивное поле нейрона 2;
Б – минимальное рецептивное поле нейрона 2;
В – рецептивное поле нейрона 3;
1 – рецептирующая клетка.
Перекрытие рецептивных полей. У первичных рецепторов зоны ветвления периферических отростков чувствительных нейронов могут перекрывать друг друга (рис. 5, А). У вторичных рецепторов одна рецептирующая клетка может контактировать с несколькими чувствительными нейронами, т. е. может входить в состав рецептивных полей различных нейронов (рис. 6.5, Б).
Рис. 5. Перекрытие рецептивных полей чувствительных нейронов первичных и вторичных рецепторов.
Взаимодействие рецепторов в рецептивном поле . При одновременном раздражении нескольких рецепторов, входящих в состав рецептивного поля одного нейрона, в этом нейроне можно получить ответ только от одного рецептора. В других рецептивных полях имеют место более сложные взаимодействия. Например, в сетчатке глаза существуют тормозные взаимодействия между рецепторами, расположенными в центральной и периферической части рецептивного поля.
Адекватный стимул, классификация рецепторов.Рецептор-это специализированная нервная клетка. Она сигнализирует в ЦНС о состоянии и (или) изменениях состояния среды, в которой находится. Те факторы окружающей среды и те их изменения, которые оказывают действие на рецепторы, называются стимулами. Стимулы-это величины, которые можно измерять объективными методами, например деформация кожи, температура и электромагнитное излучение (свет).
Физиологические исследования обнаружили, что каждый рецептор с особой готовностью реагирует на стимулы какого-то одного типа. Это свойство обычно называют специфичностью рецепторов; тот стимул, который является эффективным в каждом случае, иногда называют адекватным стимулом для данного рецептора. Для некоторых рецепторов адекватный стимул можно установить из повседневного опыта. Так, например, мы можем легко определить, что адекватным стимулом для рецепторов наших глаз является свет, тогда как тепловые или механические стимулы с помощью этих рецепторов мы в обычных условиях не обнаруживаем.
Следовательно, каждый рецептор способен передавать в ЦНС информацию только об одном определенном аспекте или размерности окружающей среды. Это значит, что рецептор играет роль фильтра. В связи с этим представляется полезным классифицировать рецепторы на основе их адекватных стимулов. Рецепторы млекопитающих распадаются на следующие" четыре группы: механо-, термо-, хемо-и фоторецепторы.
Внутри каждой из этих четырех групп мы можем усмотреть значительную степень специализации. Например, есть различающиеся типы фоторецепторов, по-разному отвечающие на излучения с различными длинами
волн; эти рецепторы называют чувствительными к длине во
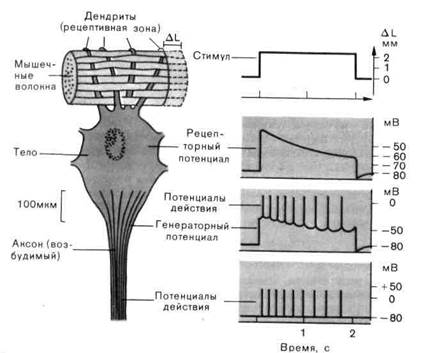
гас. г-1. схема электрофизиологических измерений, которые можно провести на рецепторе растяжения ракообразных. Во время стимуляции (изменение длины соответствующей мышцы на AL) микроэлектрод, введенный в тело клетки, регистрирует рецепторный потенциал, который возникает в рецептивной зоне (на дендритах). Являясь генераторным потенциалом, он возбуждает распространяющиеся потенциалы действия в возбудимом аксоне.
лны или цветочувствительными. Терморецепторы можно подразделить на рецепторы тепла и холода в соответствии с тем, что их возбуждает -увеличение или уменьшение температуры. У механорецепторов также можно наблюдать специализацию к различным параметрам стимулов: в коже, например, одни рецепторы чувствительны к вибрации, другие-к давлению.
Рецепторный потенциал. Вследующих параграфах мы рассмотрим процессы в рецепторе (подвергаемом действию адекватного стимула), результатом которых является потенциал действия в афферентном волокне. В качестве примера взят рецептор растяжения ракообразных-ме-ханорецептор, расположенный между определенными мышечными волокнами животного; он чувствителен к растяжению (рис. 2-1).
Когда микроэлектрод проникает в рецепторную клетку, то потенциал, который регистрируется исходно, как и во всех других нервных клетках, - это потенциал покоя (-80мВ на рис. 2-1). Под действием адекватной стимуляции - когда окружающая мышца растягивается мембранный потенциал меняется в направлении деполяризации. Эта вызываемая стимулом деполяризация-отклонение от потенциала покоя-называется рецепторным потенциалом. Рецепторный потенциал длится столько же, сколько и стимул (изменение ∆L на рис. 2-1). Однако даже тогда, когда стимул сохраняется постоянным, рецепторный потенциал снижается от своего исходного максимального значения до более низкого уровня. Такое снижение влияния стимула, поддерживаемого в течение некоторого времени на постоянном уровне, можно наблюдать практически у всех рецепторов; оно называется адаптацией.
Рецепторный потенциал в этом случае является результатом повышения проводимости мембраны, которое неспецифично и касается всех мелких ионов (Na + , K + , Са 2+ , Сl‾). В нормальных условиях единственный из этих ионов, который может обеспечить деполяризацию, это ион Na + , потому что только этот ион имеет потенциал равновесия, сдвинутый в деполяризационном направлении по отношению к потенциалу покоя. Следовательно, ионы Na + должны быть главным источником возникновения рецепторного потенциала. Повышение проводимости мембраны, описанное для рецепторов, сходно с тем, что имеет место в субсинаптической мембране возбуждающих синапсов - например в концевой пластинке мотонейрона.
Фоторецепторы позвоночных представляют исключение в том смысле, что при стимуляции светом в них возникает гиперполяризационный потенциал. Однако при синаптической передаче на нейроны высших уровней в сетчатке индуцируется деполяризация-возбуждающий пост-синаптический потенциал (ВПСП), который ведет к генерации распространяющихся потенциалов действия.
Процесс трансдукции; первичные и вторичные сенсорные клетки.Промежуточные события между поступлением стимула и появлением рецепторного потенциала пока еще мало изучены; они объединяются под общим названием процессы трансдукции. Этот термин подразумевает главным образом молекулярно-биологические процессы, имеющие место на рецептивной мембране и (или) во внутриклеточных структурах (называемых органеллами). В воспринимающей части рецептора имеется много таких органелл, как мембранные диски фоторецепторов, ко
 |
торые содержат чувствительные к свету молекулы зрительного пигмента (см. рис. 4-27) или волосковидные выросты (реснички) рецепторов внутреннего уха (см. рис. 5-3). Это вспомогательные структуры, которые делают рецептор особенно чувствительным к специфическому адекватному стимулу.
Многие рецепторы состоят из двух связанных элементов: нервного окончания и некоторой клетки ненервного происхождения типа дисков Меркеля в коже (см. рис. 3-4) или чувствительных клеток во вкусовых почках (см. рис. 7-3). Полагают, что процесс преобразования сигнала имеет место в не-нервной клетке. Такие клетки называют вторичными сенсорными клетками. На электронной микрофотографии область контакта между клетками выглядит как синапс. По этой причине предполагается, что имеется синаптическая передача от вторичной сенсорной клетки к нервному окончанию.
Напротив, первичная сенсорная клетка -это рецептор, в котором преобразование стимула происходит в нервной клетке или в нервном окончании. Примером этого являются обонятельные рецепторы (см. рис. 8-2) и свободные нервные окончания в коже.
Рецепторный потенциал как причина генерации распространяющихся потенциалов действия.Рецепторный потенциал электротонически распространяется в смежные области клетки. В результате этого аксон деполяризуется (рис. 2-1). Когда деполяризация достигает порога мембраны аксона, генерируется потенциал действия. Этот потенциал действия проводится по аксону в ЦНС. Таким образом, рецепторный потенциал действует на аксон как электрический стимул; по этой причине его также называют генераторным потенциалом.
Если генераторный потенциал не спадает после генерации первого потенциала действия, он может вызывать следующие потенциалы действия, пока стимул и соответственно генераторный потенциал не исчезнут. Это значит, что в период существования длительного генераторного потенциала афферентное волокно разряжается многократно. Процессы, происходящие между поступлением стимула и генерацией распространяющихся потенциалов действия, могут быть названы процессами трансформации стимула в нервную активность. Таким образом, эта трансформация включает следующие события: трансдукцию, увеличение проводимости рецептивной мембраны, рецепторный потенциал, синаптическую передачу (в случае вторичных сенсорных клеток), генераторный потенциал, потенциалы действия.
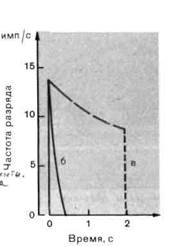
Адаптация. При рассмотрении рецепторного потенциала мы отметили, что даже если стимул поддерживается некоторое время постоянным, рецепторный потенциал может уменьшаться (рис. 2-1); этот процесс называют адаптацией. Мы также наблюдаем этот феномен на картине разряда афферентных нервных волокон: как можно видеть из рис. 2-1, при неизменном стимуле временные интервалы ∆t между двумя последовательными импульсами удлиняются, т.е. мгновенная частота разряда уменьшается (мгновенная частота = 1/∆t). Адаптация может иметь место на всех стадиях трансформации стимула: в процессе трансдукции, в механизме проведения рецепторного потенциала, при синаптической передаче от вторичных сенсорных клеток и при генерации потенциала действия. Возможно, что несколько таких механизмов работает одновременно.

Рис. 2-2. Разряды рецепторов с различными скоростями адаптации. А. Потенциалы действия очень быстро (а), быстро (б) и медленно (в) адаптирующихся рецепторов, генерируемые при длительном действии постоянного стимула. Б. Графики изменения мгновенной частоты разряда со временем для рецепторов (б) и (в). Обратите внимание на то, что разряд рецептора (в) включает как фазиче-ский, так и тонический компоненты.
Чтобы получить адаптационную характеристику рецептора, исследуют его реакцию на надпороговый ступенчатый стимул. В таких экспериментах различные рецепторы демонстрируют различные изме
нения частоты разряда во времени. На рис. 2-2, А показан разряд медленно адаптирующегося рецептора (в), умеренно адаптирующегося рецептора (б) и быстро адаптирующегося рецептора (а). На рис. 2-2, Б представлено изменение со временем мгновенной частоты для рецепторов с медленной и умеренной адаптацией.
Предложены различные функциональные объяснения адаптации. Это явление обычно рассматривают всего лишь как простое утомление рецепторов. Но во многих случаях различное развитие адаптации во времени можно интерпретировать как приспособление к передаче специфических параметров стимула. Например, при изучении трех типов ме-ханорецепторов кожи было обнаружено, что благодаря характерным временным свойствам их разряда они могут рассматриваться как пере-
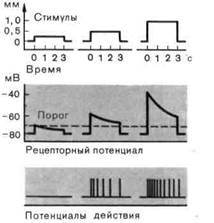 |
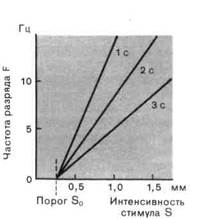 Рис. 2-3. Зависимость рецепторного потенциала и частоты разряда от интенсивности стимула. А. Рецепторные потенциалы и потенциалы действия при трех различных интенсивностях стимула. Потенциалы действия возбуждаются тогда, когда рецепторный потенциал превосходит порог (красная прерывистая линия). Б. Графики зависимости мгновенной частоты разряда F в разные моменты после включения стимула (через 1, 2 и 3 с) от интенсивности стимула S. S0 -пороговая интенсивность. |
 |
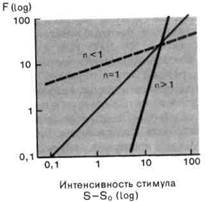 |
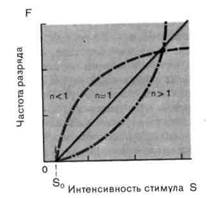
Рис. 2-4. Различные степенные функции. А. Частота разряда F как функция интенсивности стимула S. По обеим осям масштаб линейный, единицы-произвольные. Приведены три характерные степенные функции с показателями п 1 и п = 1 (красная линия). Б. Те же степенные функции изображены в логарифмическом масштабе. По абсциссе отложена разность между интенсивностью стимула S и пороговой интенсивностью S0.
датчики сведений об амплитуде, скорости и ускорении деформации кожи (см. рис. 3-5-3-8 и табл. 3-1).
Перевод интенсивности стимула в частоту импульсов в разряде.Когда сила стимула возрастает, возрастает и амплитуда рецепторного потенциала. Постепенные плавные изменения рецепторного потенциала вызывают соответствующие изменения частоты потенциалов действия. Эта ситуация представлена на рис. 2-3, А. Соотношение между интенсивностью стимула S и частотой импульсов F, часто называемое силовой функцией, изображено на рис. 2-3, £ для разных моментов времени от начала стимуляции (через 1, 2 и 3 с). В случае рецептора растяжения соотношение между F и S линейно для каждого момента времени на протяжении действия стимула.
Поскольку для того, чтобы вызвать потенциалы действия, рецепторный потенциал должен превзойти некоторое минимальное значение (порог аксона), разряд возникает только при превышении стимулом соответствующей интенсивности (S0 на рис. 2-3, Б). S0 называют пороговым стимулом для рецептора. Силовые функции рис. 2-3, Б описываются выражениями вида
т. е. линейной зависимостью для S > S0. Коэффициент к, наклон прямой, становится все меньше для последовательных интервалов времени, от-
считываемых от момента приложения стимула; это тоже является одним из выражений адаптации. '
Экспериментально было показано, что для большинства рецепторов силовая функция нелинейна. Измеренные соотношения часто можно описать различными математическими функциями, такими, как логарифмическая
Мы сталкивались с обоими этими выражениями в разделе по психофизике; они называются соответственно функциями Вебера-Фехнера иСтивенса. Кодирование интенсивности в рецепторах большинства типов лучше всего описывается степенной функцией (рис. 2-4), в которой разность- между интенсивностью стимула S и пороговой интенсивностью S0 возводится в n-ю степень. Показатель п является константой, характерной для каждого типа рецепторов. Для п 1 степенные функции (на графиках с линейным масштабом) представляются соответственно выпуклыми и вогнутыми кривыми (рис. 2-4, А), а для п = 1 силовая функция линейна, как и для рецептора растяжения. Оказалось, что для большинства рецепторов и + (значением рН);
д) медленно нарастает при действии постоянного стимула
и длится в течение всего времени стимуляции.
Верны несколько ответов.
В 2.3. В афферентных аксонах многих рецепторов частота импульсов в разряде
а) увеличивается с увеличением интенсивности стимула;
б) увеличивается со временем при действии стимула постоянной интенсивности;
в) уменьшается со временем при действии стимула постоянной
г) обычно равна нулю при действии стимулов подпороговой
д) не зависит от величины рецепторного потенциала. Верны несколько ответов.
В 2.4. В случае многих рецепторов степенная функция описывает
а) временной ход адаптации;
б) временной ход рецепторного потенциала при увеличении силы
стимула;
в) зависимость частоты импульсации F от величины превышения
стимулом порога S0;
г) связь между амплитудой потенциала действия и величиной ре
цепторного потенциала.
Дата добавления: 2015-10-05 ; просмотров: 1838 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Рецепторы – это специализированные чувствительные образования, воспринимающие раздражения из внешней и внутренней среды организма и преобразующие их в специфическую активность нервной системы
Различают клеточные и сенсорные рецепторы
Клеточные рецепторы
Расположены в наружных и внутренних мембранах различных клеток организма, осуществляют их взаимодействие с окружающей средой и обмен информацией между органеллами внутри клетки (рецепторы мембран ядра, митохондрий, рибосом, лизосом и др.)
Сенсорные рецепторы – это высокоспециализированные образования, воспринимающие раздражители внешней и внутренней среды, трансформирующие энергию раздражителя в энергию нервных импульсов (в виде рецепторного потенциала) и кодирующие свойства раздражителя
Сенсорные рецепторы классифицируются
1. По локализации в организме
Экстерорецепторы – информируют организм об изменениях во внешней среде (слуховые, зрительные, обонятельные, вкусовые, тактильные и др.)
Интерорецепторы информируют об изменениях во внутренней среде (вестибулорецепторы, проприорецепторы, хеморецепторы и др.)
2. По модальности (природе) адекватного раздражителя: механо-, фото-, хемо-, баро-, ноци- и др. рецепторы
3. По восприятию раздражителей одной или нескольких модальностей
Мономодальные рецепторы приспособлены к восприятию раздражителей одной модальности: большинство экстерорецепторов и часть интерорецепторов (например, хеморецепторы каротидной зоны)
Полимодальные рецепторы приспособлены к восприятию раздражителей нескольких модальностей (большинство интерорецепторов)
4. По скорости адаптации
Быстроадаптирующиеся: зрительные, слуховые, вкусовые, болевые и др. Медленноадаптирующиеся: проприо-, рецепторы растяжения легких, др.
5. По структурно-функциональным особенностям
Свободные нервные окончания – тонкие, безмиелиновые терминали дендритов чувствительных нейронов, свободно контактирующие с раздражителем (например, рецепторы прикосновения)
Являются полимодальными
Имеют самую низкую чувствительность среди всех рецепторов
Инкапсулированные тельца – рецепторы в виде нервного окончания, находящиеся во внутренней колбе, окруженные соединительной капсулой (например, тельца Фатера-Пачини – рецепторы давления)
Рецепторы со специальными клетками могут быть двух видов:
Первичночувствующие рецепторы (нейросенсорные и чувствительные рецепторы: рецепторы обоняния) являются мембранными белками нейрона и вступают в непосредственный контакт с сигналом (пахучим веществом).
Сама нервная клетка перемещена на периферию (например, в выстилку слизистой носа).
В этом же нейроне происходитформирование рецепторого потенциала, а затем генерация потенциала действия на аксонном холмике (первичночувствующий нейрон)
Рецепторный потенциал = генераторному потенциалу
Вторичночувствующие рецепторы (сенсороэпителиальные рецепторы: вкусовые, слуховые, зрительные, вестибулярные и др.) представлены специализированными клетками (чаще эпителиальная), которые не имеет продолжения (не являются окончаниями дендрита).
В ответ на возникший рецепторный потенциал она выделяет медиатор, который взаимодействует с окончанием дендирита нейрона. В результате возникает генераторный потенциал
Сенсорная рецептия — (от лат. recipere — принимать, получать) — процесс трансформации стимульной энергии (механической, термической, электромагнитной, химической и др.) в нервные сигналы, осуществляемый рецепторами, процесс восприятия и трансформации (преобразования) энергии внешнего раздражителя в энергию нервного импульса или в сложную последовательность внутриклеточных процессов. [1] Рецептия это:
- Клеточная и сенсорная рецепция;
- Классификация и строение сенсорных рецепторов;
- Преобразование энергии в сенсорном рецепторе;
- Свойства рецепторов;
- Рецептивное поле.
Клеточная и сенсорная рецепция Править
Клеточная рецепциия — процесс восприятия и преобразования химического сигнала в сложную последовательность внутриклеточных химических процессов. Клеточная рецепция обеспечивает возможность обмена информации между клетками, который осуществляется при помощи биологически активных веществ (гормонов, медиаторов). Обязательным этапом такого межклеточного взаимодействия является связывание молекул вещества с соответствующей молекулой клетки-мишени, называемой клеточным рецептором. Роль клеточных рецепторов играют специфические белковые молекулы, которые могут быть расположены на поверхности клетки, в цитоплазме или в ядре.
Сенсорная рецепция — процесс восприятия и преобразования энергии раздражителей внешней и внутренней среды организма в энергию нервных импульсов, передаваемую по чувствительным нервам в ЦНС. Сенсорный рецептор представляет собой нервную клетку или комплекс нервной и эпителиальной клетки, специально приспособленный для восприятия определенного типа раздражителей. Сенсорные рецепторы являются начальными звеньями любой рефлекторной дуги, а также участвуют в оценке параметров полезного приспособительного результата в функциональных системах организма.
Классификация и строение сенсорных рецепторов Править
По строению рецепторы подразделяют на первичные и вторичные (cм. рис. 1).
- К первичным рецепторам относят такие сенсорные рецепторы, у которых действие раздражителя воспринимается непосредственно периферическими отростками чувствительного нейрона (нервными окончаниями), которые могут быть:
- свободными, т. е. не имеют дополнительных образований;
- инкапсулированными, т.е. окончания чувствительного нейрона заключены в особые образования, осуществляющие первичное преобразование энергии раздражителя.
- К вторичным рецепторам относят такие сенсорные рецепторы, у которых действие раздражителя воспринимается специализированной рецептирующей клеткой не нервного происхождения. Возбуждение, возникшее в рецептирующей клетке, передается через синапс на чувствительный нейрон.
Рис.1. Первичные и вторичные рецепторы.
- 1 – тело чувствительного нейрона;
- 2 – периферический отросток чувствительного нейрона (дендрит);
- 3 – центральный отросток чувствительного нейрона (аксон);
- 4 – глиальная капсула; (Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в среднем в 10-50 раз больше, чем нейронов.)
- 5 – рецептирующая клетка;
- 6 – синапс между рецептирующей клеткой и чувствительным нейроном.
Тело чувствительного нейрона обычно располагается за пределами ЦНС: в спинномозговом или вегетативном ганглии. От такого нейрона отходят два отростка – дендрит, который следует к периферическим органам и тканям, и аксон, который направляется в спинной мозг.
По расположению сенсорные рецепторы подразделяют на:
- экстерорецепторы – воспринимают раздражители из внешней среды организма;
- интерорецепторы – воспринимают раздражители из внутренней среды организма;
- проприорецепторы – специализированные рецепторы опорнодвигательной системы.
По разнообразию воспринимаемых раздражителей сенсорные рецепторы подразделяют на:
- мономодальные – приспособлены для восприятия только одного вида раздражителя;
- полимодальные – приспособлены для восприятия различных видов раздражителей.
По модальности сенсорные рецепторы подразделяют на:
- хеморецепторы – воспринимают действие химических веществ;
- фоторецепторы – воспринимают световые раздражители;
- механорецепторы – воспринимают давление, вибрацию, перемещение, степень растяжения;
- терморецепторы – чувствительны к изменениям температуры;
- ноцицепторы – воспринимают болевое раздражение.
Преобразование энергии в сенсорном рецепторе Править
Этапы преобразования энергии внешнего раздражителя в энергию нервных импульсов.
- Действие раздражителя. Внешний стимул взаимодействует со специфическими мембранными структурами окончаний чувствительного нейрона (в первичном рецепторе) или рецептирующей клетке (во вторичном рецепторе), что приводит к изменению ионной проницаемости мембраны.
- Генерация рецепторного потенциала. В результате изменения ионной проницаемости происходит изменение мембранного потенциала (деполяризация или гиперполяризация) чувствительного нейрона (в первичном рецепторе) или рецептирующей клетке (во вторичном рецепторе). Изменение мембранного потенциала, наступающее в результате действия раздражителя, называют рецепторным потенциалом (РП) .
- Распространение рецепторного потенциала. В первичном рецепторе РП распространяется электротонически и достигает ближайшего перехвата Ранвье. Во вторичном рецепторе РП электротонически распространяется по мембране рецептирующей клетки и достигает пресинаптической мембраны, где вызывает выделение медиатора. В результате срабатывания синапса (между рецептирующей клеткой и чувствительным нейроном) происходит деполяризация постсинаптической мембраны чувствительного нейрона (ВПСП). Образовавшийся ВПСП распространяется электротонически по дендриту чувствительного нейрона и достигает ближайшего перехвата Ранвье.
- В области перехвата Ранвье РП (в первичном рецепторе) или ВПСП (во вторичном рецепторе) преобразуется в серию ПД (нервных импульсов). Образовавшиеся нервные импульсы проводятся по аксону (центральному отростку) чувствительного нейрона в ЦНС. Поскольку РП генерирует образование серии ПД, его часто называют генераторным потенциалом .
Закономерности преобразования энергии внешнего раздражителя в серию нервных импульсов (см. рис. 2):
Рис.2. Преобразование энергии внешнего раздражителя в рецепторный потенциал и серию нервных импульсов при действии слабого (А ) и сильного (Б) раздражителя.
- чем выше сила действующего раздражителя, тем больше амплитуда РП;
- чем больше амплитуда РП, тем больше частота нервных импульсов.
Свойства рецепторов Править
Специфичность. Большинство рецепторов приспособлены для восприятия только одного вида раздражителей (только одной модальности). Специфичность таких мономодальных рецепторов не является абсолютной – практически любой рецептор реагирует на разные раздражители. Однако пороговая сила того раздражителя, к восприятию которого рецептор приспособлен, значительно ниже таковой для всех прочих раздражителей. Рецепторы одной и той же модальности могут подразделяться на несколько групп в зависимости от характеристик воспринимаемого раздражителя. Например, Колбочки (сетчатка). Они отвечают за цветовое зрение и лучше функционируют в сравнительно ярком свете, в отличие от палочек, которые работают лучше при тусклом свете. Колбочки являются ответственными за цвет, цветное зрение; колбочки функционируют лучше всего в относительно ярком свете c длиной волн более 498 нм, в противоположность клеткам фоторецепторов палочек работают лучше в тусклом Свете с длиной волн менее 498 нм. Колбочки плотно упакованы в ямке фовеа "централь", с диаметром 0.3 мм стержня — свободная зона с очень тонкими, плотно упакованными колбочками, которые быстро уменьшаются количественно к периферии сетчатки глаза. [2]

Рис.1а. Схематическое изображение фоторецепторов черепахи и связанных с ними капель масла. (Нефтяные капельки). UV, UV (ультрафиолетовые) колбочки, B, синяя колбочка, DC, двойая колбочка, G, зеленая колбочка, R, красная колбочка, палочка (штанга), (без капелек масла) . [3]
Вариант колбочки с тремя пигментами опсина S,M,L) (См.рис.1a), которые часто не корректно иллюстрируют в виде трёх колбочек (как трихроматизм), отличаются тем, что колбочка содержат различные пигменты, определяющие цветовое ощущение (зрение), а именно: S-колбочки (синий цвет), M-колбочки (зелёный цвет) и L-колбочки (красный цвет). Колбочка поэтому чувствительна к видимым длинам волн света, которые соответствуют длинам волн: коротковолновой, средней длине волны и длинной волне света RGB (См. рис.1a). Откуда и пошло трихроматизм (три цвета). [4] [5]
Чувствительность. Количественной мерой чувствительности сенсорного рецептора является абсолютный порог чувствительности – минимальная сила раздражителя, способная вызвать возбуждение рецептора.
Адаптацией называют явление ослабления возбуждения в рецепторе при действии длительного раздражителя постоянной силы.
Рис.3. Адаптация тонических ( А ), промежуточных ( Б ) и фазных рецепторов ( В ) к длительно действующему раздражителю постоянной силы.
В зависимости от скорости адаптации рецепторы подразделяют на:
- тонические (пропорциональные) рецепторы генерируют нервные импульсы в течение всего времени действия раздражителя; после высокочастотного залпа в начале действия раздражителя частота нервных импульсов устанавливается на постоянном уровне (рис. 3, А);
- промежуточные (фазнотонические) рецепторы генерируют нервные импульсы в течение всего времени действия раздражителя, однако их частота существенно уменьшается (рис. 3, Б);
- фазные (дифференциальные) рецепторы генерируют нервные импульсы в начальный (ON-ответ — ДА) и конечный (OFF-ответ — НЕТ) период действия раздражителя (рис. 4, В).
Рецептивное поле Править
Рецептивным полем нейрона называют множество рецепторов, функционально связанных с этим нейроном. Рецептивное поле нейрона представляет собой динамическое образование – один и тот же нейрон в различные моменты времени может оказаться функционально связанным с различным количеством рецепторов. Максимальная величина рецептивного поля какого-либо нейрона соответствует количеству рецепторов, которые связаны с эти нейроном морфологически, а минимальная величина может ограничиваться всего одним рецептором (см. рис. 4).
Рис.4. Рецептивные поля нейронов.
- А – максимальное рецептивное поле нейрона 2;
- Б – минимальное рецептивное поле нейрона 2;
- В – рецептивное поле нейрона 3;
- 1 – рецептирующая клетка.
Перекрытие рецептивных полей. У первичных рецепторов зоны ветвления периферических отростков чувствительных нейронов могут перекрывать друг друга (рис. 5, А). У вторичных рецепторов одна рецептирующая клетка может контактировать с несколькими чувствительными нейронами, т. е. может входить в состав рецептивных полей различных нейронов (рис. 6.5, Б).
Рис.5. Рецептивные поля нейронов.
Взаимодействие рецепторов в рецептивном поле . При одновременном раздражении нескольких рецепторов, входящих в состав рецептивного поля одного нейрона, в этом нейроне можно получить ответ только от одного рецептора. В других рецептивных полях имеют место более сложные взаимодействия. Например, в сетчатке глаза существуют тормозные взаимодействия между рецепторами, расположенными в центральной и периферической части рецептивного поля. [6]
Читайте также:
