
Принципы функционирования нервных центров
Принцип доминанты сформулирован А.А Ухтомским, Под доминантой А.А.Ухтомский понимал господствующий очаг возбуждения, определяющий характер текущих реакций центров в данный момент. Этот господствующий центр может возникнуть при длительном действии гуморальных или рефлекторных раздражителей. Для него характерно: а) наличие повышенной возбудимости; б) иннерционность, обусловленная длительными следовыми процессами; в) способность к суммации и сопряженному торможению других центров, функционально несовместимых с деятельностью центров доминантного очага. Доминанта создаëт определенный вектор поведения.
Примером может служить обнимательный рефлекс у лягушки в результате гормональных воздействий. Гормоны в сочетании с раздражением кожи (рецепторы – мозоли пальцев передних конечностей самца) создают суммацию раздражителей.
А.А.Ухтомский считал, что доминанта является общим принципом работы нервных центров, что она связана с возбуждением или активацией многих нервных центров, временно кооперирующихся для выполнения биологически важной функции. Взаимодействие работающих центров на единый ритм активности (усвоение ритма) рассматривается как динамический функциональный орган.
Согласно современным представлениям (теория функциональных систем П.К.Анохина) любой поведенческий акт, в том числе условный рефлекс – результат анализа и синтеза афферентной информации, включающей, прежде всего, возбуждение центральных рецепторов латерального, переднего гипоталамуса, преоптической области, реагирующих на содержание в крови питательных веществ, половых гормонов и других факторов. Доминирующий очаг может возникнуть не только под влиянием гормональных воздействий, но и под влиянием нервных сигналов, изменяющих возбудимость центральных нейронов.
Иннерционность доминанты - это способность доминантного состояния поддерживаться довольно долго, что обусловлено длительностью следовых процессов. Следовое возбуждение в свете открытий современной физиологии может быть обусловлено следующими факторами:
Суммацией взаимодействия возбуждающих потенциалов, вызванных приходящими к нейронам подпороговыми нервными импульсами.
1.Изменением концентрации ионов К + в синаптической щели, которые как деполяризаторы, усиливают вхождение ионов Са² + в пресинаптическое окончание.
2.Следами метаболитов, связанных с медиаторами.
3.Циклическими связями в ЦНС, способными обеспечить следовую самостимуляцию центров (следовые процессы храняться различное время в гиппокампе – часы, сутки – предполагают, что здесь хранятся следовые процессы от пережитых доминат; в спинальных центрах – постсинаптический потенциал длится минуты).
Доминатное состояние как системная реакция может не только суммировать посторонние раздражения, но и может тормозить конкурирующие центры.
Феномен сопряженного торможения конкурирующих центров возможен благодаря динамическому функциональному органу как временной кооперации нервных центров.
Существуют другие теории, объясняющие сопряженное торможение. В 1957 году В.Маунткасл (в опытах использовал кошек, вводя в сенсомоторную кору электроды под углом и перпендикулярно) выдвинул гипотезу колончатой организации коры больших полушарий, согласно которой головной мозг – это совокупность нейронных модулей, связанных многократно. Доминирование одного из модулей и вызывает динамику систем.
Проведение возбуждения через нервные центрыосуществляется тольков одном направлении: с афферентного нейрона на эфферентный. Свойство объясняется строением синапсов – медиаторы выделяются только концевыми аппаратами аксонов и возбуждение распространяется к постсинаптической мембране.
Задержка проведения в синапсах и время рефлекса обусловлена более медленным проведением нервных импульсов через синапсы, т.к. необходимо время для выделения медиатора, его диффузию через синаптическую щель к постсинаптической мембране и возникновения возбуждающего постсинаптического потенциала под действием медиатора. Чем больше сила раздражителя, тем короче время рефлекса.
Иррадиация возбуждения-это способность возбуждения широкой волной разливаться по ЦНС от центра к центру во всех направлениях, что объясняется наличием огромного количества коллатералей. От каждого аксона отходят идут коллатерали к множеству нейронов, от них идет еще большее число коллатералей. Т.о. импульс, пришедший в ЦНС, может иррадиировать по многим направлениям ко многим центрам.
Конвергенции -свойство нервных центров, противоположное иррадиации. Схождение, сужение – к эфферентному нейрону возбуждение проходит по многим путям, т.к. афферентных путей в 4-5 раз больше.
Циркуляция нервных импульсов по замкнутым нейронным цепям
может длительно осуществляться в одном нервном центре благодаря тому, что импульсы, возникнув в одном из нейронов, передаются к другим нейронам, а затем по коллатералям их аксонов возвращаются на первый нейрон.
Утомляемость нервных центров обуславливается высоким обменом веществ, что приводит к нарушением проведения возбуждения в межнейронных синапсах. Уменьшаются запасы медиатора в окончаниях аксона, падает чувствительность к медиатору постсинаптической мембраны, понижаются ресурсы нервной клетки.
Пластичность нервных центров-перемена функций нервного центра в случае, если рабочий орган, с которым данный орган связан, заменить другим.
Свойства нервных центров. К ним относят:
замедленное распространение возбуждения, обусловленное наличием синапсов и многих нейронов на путях передачи возбуждения;
одностороннее распространение возбуждения, обусловленное тем, что основную роль в распространении возбуждения играют химические синапсы, в которых возбуждение передается только от пресинаптических структур на постсинап- тические структуры;
иррадиация возбуждения — распространение возбуждения от активированного нервного центра на соседние и функционально связанные нейронные структуры. Это свойство обусловлено наличием многочисленных ветвлений аксонов, заканчивающихся синаптическими контактами. В результате при активации группы нейронов от них идут возбуждающие влияния на синаптически связанные с ними другие нейронные группы. При прекращении притока возбуждающей импульсации активность нервного центра уменьшается и может наступать концентрация возбуждения — процесс, обратный иррадиации;
суммация возбуждения, проявляющаяся увеличением интенсивности рефлекторной реакции при возрастании длительности раздражения, его силы или площади раздражаемого рецепторного поля. Различают временную и пространственную суммацию. При временной суммации количество активируемых синаптических образований на эфферентном нейроне не изменяется, но увеличивается частота импульсаций, приходящих к каждому синапсу. При пространственной суммации из-за воздействия на новые рецепторы увеличивается количество одновременно активируемых синаптических входов и эфферентный нейрон быстрее и эффективнее возбуждается. В обоих случаях возрастает величина ответной реакции эффекторных структур. Например, увеличивается интенсивность сокращения мышцы, секреции слезной или слюнной железы.
трансформация ритма возбуждений, проявляющаяся изменением частоты импульсаций, генерируемых эфферентными нейронами, по отношению к частоте импульсаций, передаваемых афферентными нейронами. Такое изменение становится возможным благодаря суммации ВПСП, функционированию локальных нейронных цепей и т.д.;
явление последействия, заключающееся в способности нервных центров длительно поддерживать возбуждение после устранения раздражителя, вызвавшего его. Одним из механизмов этого служит возможность длительной циркуляции возбуждения в замкнутых нейронных цепях;
тонус нервных центров, проявляющийся длительным непрерывным возбуждением нервных центров. Такое состояние поддерживается за счет непрерывного притока афферентных импульсаций, влияния биологически активных веществ, доставляемых гуморальным путем, циркуляцией возбуждения в замкнутых нейронных цепях, спонтанной активностью отдельных нейронов;
пластичность нервных центров — способность к перестройке функциональных свойств и изменению выполняемых функций. Например, экспериментально показана способ
но ность диафрагмальных мотонейронов вместо обеспечения ритмических сокращений диафрагмы выполнять функцию регуляции шагательных движений ног;
повышенная утомляемость нервных центров по отношению к утомляемости периферических нервных структур и мышц- В системе нервный центр — нервно-мышечный препарат первое место по утомляемости занимает нервный центр, второе — нервно-мышечный синапс, третье — скелетная мышца и наименее утомляемым оказывается нервное волокно;
высокая чувствительность нервных центров к действию биологически активных веществ и ядов, нарушениям гомеостаза и недостатку кислорода. При прекращении доставки кислорода клетки коры мозга могут сохранять жизнедеятельность не более 5 мин, уменьшение содержания глюкозы в крови ниже 800 мг/л сопровождается гипогликемической комой.
Принципы функционирования, интегративной и координационной деятельности ЦНС. Интегративная деятельность ЦНС проявляется восприятием, объединением и переработкой комплекса поступающей к нервным центрам информации и принятием решений, обеспечивающих регуляцию гомеостаза, а также вегетативных и соматических реакций, направленных на достижение полезного для организма приспособительного результата. Координационной деятельностью называют регуляцию распределения возбуждения и торможения в нейронных структурах, а также взаимодействия нервных центров, обеспечивающие адекватные рефлекторные и произвольно вызываемые реакции.
Принцип рефлекторных регуляций — важнейший принцип функционирования нервной системы.
Рефлексом называют стереотипную ответную реакцию организма на действие раздражителя, осуществляющуюся благодаря регуляторной роли нервных клеток, составляющих рефлекторную дугу.
Абсолютное большинство рефлексов замыкается в головном и спинном мозге. Но есть также рефлексы, рефлекторная дуга которых замыкается вне центральной нервной системы в вегетативных внеорганных ганглиях или даже в пределах одного органа (например, сердца или кишечника).
Рефлекторная дуга включает 5 звеньев. Начальным звеном является рецептор, затем идет афферентный (чувствительный, Центростремительный) нейрон, ассоциативный (вставочный)
нейрон, эфферентный (двигательный, центробежный) нейрон и эффектор. Эффектором может быть любая структура (мышца, железа и т.д.), на которой заканчивается синапсом эфферентный нейрон. Вставочный нейрон может быть один или их может быть много. Они располагаются в нервных центрах.
Следовательно, в образовании рефлекторной дуги участвует как минимум 3 нейрона. Исключение составляет лишь один вид рефлексов так называемые сухожильные рефлексы, рефлекторная дуга которых включает только 2 нейрона: афферентный и эфферентный. Отросток афферентного нейрона входит в спинной мозг через задние корешки и, проникая в передние рога серого вещества, формирует синапс прямо на эфферентном нейроне. Примеры схем двух- и трехнейронной рефлекторной дуги соответственно сухожильного и оборонительного сгибательного рефлекса, вызываемого раздражением кожи, представлены на рис. 6.4.
Область сосредоточения рецепторов, с которых запускается определенный рефлекс, называют рецепторным полем этого рефлекса.
Разрез спинного мозга
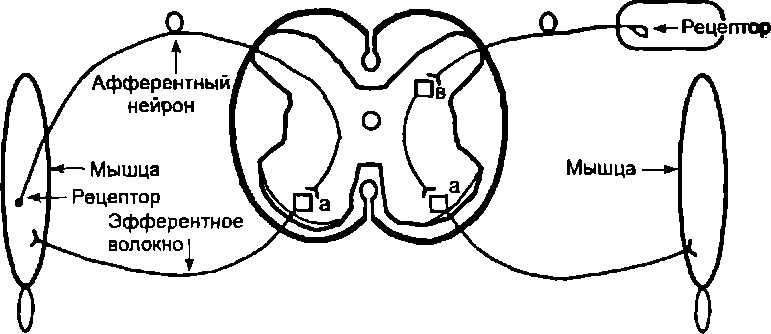
Рис. 6.4. Схема двухнейронной рефлекторной дуги сухожильного рефлекса и трехнейронной рефлекторной дуги сгибательного оборонительного
рефлекса: а — а-мотонейрон, в — вставочный нейрон
Классификация рефлексов: все рефлекторные реакции подразделяют на безусловные и условные. Безусловные врожденные и проявляются при воздействии специфического
раздражителя на строго определенное рецепторное поле. Условные — приобретаются или вырабатываются в процессе жизни. Подробная их характеристика будет дана при изучении высшей нервной деятельности. Здесь же рассмотрим наиболее употребительные варианты классификации безусловных рефлексов.
По биологической значимости рефлекторной реакции выделяют пищевые, оборонительные, половые, ориентировочные, статокинетические рефлексы.
По виду рецепторов различают эстероцептивные, инте- роцептивные, проприоцептивные рефлексы. Среди последних выделяют сухожильные и миотатические рефлексы.
По участию в рефлексе соматических или вегетативных отделов ЦНС и органов эффекторов различают соматические и вегетативные рефлексы. Если эффектор и рецепторное поле рефлекса относятся к соматическим структурам, то такие рефлексы называют соматическими. Вегетативными называют рефлексы, эффектором в которых являются внутренние органы, а эфферентная часть рефлекторной дуги образована вегетативными нейронами. Примером вегетативного рефлекса является рефлекторное замедление сердечной деятельности, вызванное воздействием на рецепторы желудка. Примером соматического рефлекса является сгибание руки в ответ на болевое раздражение кожи.
По уровню замыкания рефлекторной дуги в центральной нервной системе выделяют спинальные, бульварные (замыкающиеся в продолговатом мозге), мезенцефальные и талами- ческие рефлексы.
По количеству нейронов, участвующих в замыкании рефлекторной дуги, и числу центральных синапсов выделяют двухнейронные, трехнейронные, мультинейронные; моносинап- тические, полисинаптические рефлексы.
Благодаря принципу обратной связи (обратной афферентации) формируется замкнутый регуляторный контур. Только замкнутая регуляторная система может обеспечить устойчивое, адекватное регулировние. Благодаря обратной связи осуществляется контроль за исполнением, результатом влияния управляющего сигнала на эффекторные структуры и величиной регулируемого параметра.
Участие обратной связи даже в простейших регуляциях можно рассмотреть на примере реализации сгибательного
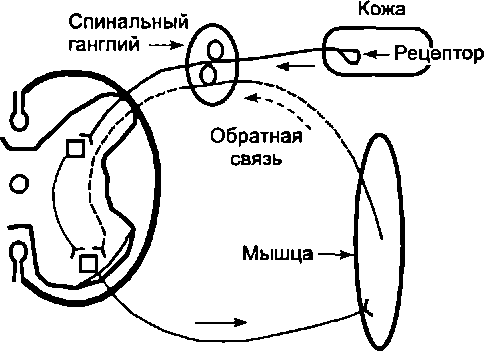
Рис. 6.5. Обратная связь в простейших рефлекторных реакциях
рефлекса, вызываемого прикосновением к коже (рис. 6.5). При рефлекторном сокращении мышцы изменяется активность находящихся в ней рецепторов и частота импульсаций в афферентных волокнах, идущих к мотонейронам. В результате формируется замкнутый контур регулирования, в котором роль канала обратной связи выполняют афферентные волокна, передающие импульсацию в нервные центры от рецепторов мышц, а роль канала прямой связи — эфферентные волокна, идущие к мышцам от мотонейронов. Таким образом, нервный центр (его мотонейроны) получает информацию об изменении состояния мышцы, вызванном передачей импульсов по двигательным волокнам. Благодаря обратной связи формируется своеобразное регуляторное кольцо. Поэтому некоторые авторы даже предлагали вместо термина "рефлекторная дуга" применять термин "рефлекторное кольцо"
Участие обратной связи в механизме регуляции кровообращения, дыхания, температуры тела и поведенческих реакций рассматривается ниже в соответствующих разделах.
Принцип реципрокной связи проявляется во взаимодействии центров-антагонистов по функциональному назначению, например группы мотонейронов, ответственных за сгибание руки, и группы, управляющей мышцами-разгибателями руки. При реципрокной связи возбуждение одного из антагонистических центров приводит к торможению другого. Это осуществляется за счет активации тормозных нейронов на конечном участке проводящих путей, идущих от возбужденного центра к антагонисту.
Принцип конвергенции заключается в схождении, поступлении к одному и тому же нейрону импульсаций от различных нервных центров или рецепторов различных модальностей (различных органов чувств). На основе конвергенции самые разные раздражители могут вызвать однотипную реакцию. Например, сторожевой рефлекс (поворот головы, насторажи- вание) может быть вызван и световым, и звуковым, и тактильным воздействием. Возможность конвергенции импульсаций с разных входов на одни и те же эфферентные нейроны называют принципом общего конечного пути.
Принцип дивергенции утверждает возможность расхождения импульсаций от одного нейрона сразу на многие нейроны. На основе дивергенции происходит иррадиация возбуждения и становится возможным быстрое вовлечение в ответную реакцию многих центров, расположенных на разных уровнях ЦНС.
Принцип доминанты характеризует особенности взаимодействия нервных центров. Доминантный очаг возбуждения обладает стойкой высокой активностью, он подавляет возбуждение в других нервных центрах, подчиняет их своему влиянию, притягивает к себе афферентные импульсации, адресуемые к другим центрам, и усиливает свою активность за счет этих импульсаций. Доминантный центр может длительно находиться в состоянии возбуждения без признаков утомления.
Примером рефлекторной реакции, вызванной доминантным очагом возбуждения, может служить обнимательный рефлекс самца лягушки, проявляющийся в период размножения. Если в этот период положить самца лягушки брюшком на палец, то у животного возникает обнимательный рефлекс. Самец обхватывает и сжимает передними лапками палец. Затем металлическим пинцетом проводят по спинке животного. Это воздействие обычно вызывает оборонительную реакцию (бегства). Но в данных условиях воздействие пинцетом лишь усиливает обнимательный рефлекс, самец сильнее сдавливает палец.
Контрольные вопросы и задания
Какова роль нервной системы в организме ?
Как подразделяют нервную систему ? Какие клетки ее образуют?
Какова роль глиальных клеток в ЦНС?
Каковы структура и функции нейронов и их частей?
Каковы особенности поляризации мембрены нейрона и возбудимости его отделов?
Каковы особенности передачи возбуждения в центральных синапсах? Что такое медиаторы?
7 Перечислите виды и механизмы торможения.
Что такое интеграция возбуждения на нейроне?
Какова классификация нейронов?
Что такое нейронные цепи?
Каковы свойства нервных центров?
Дайте характеристику рефлекса и рефлекторной дуги.
Какова классификация рефлексов?
Каковы принципы осуществления нервных регуляций?
Как соотносится длительность латентных периодов сухожильных и оборонительных рефлексов, вызываемых раздражением кожи? Почему имеются такие различия?
Зарисуйте схему нейронной сети, которая может увеличить частоту эфферентных импульсов по отношению к частоте афферентных.
Какие механизмы обусловливают способность спинальных мотонейронов генерировать частоту возбуждения до 50 имп/с, а вставочных — до 500 имп/с?
Почему при наличии обнимательного рефлекса у самца лягушки ряд раздражителей перестает вызывать оборонительную реакцию, а вместо нее лишь усиливает обнимательный рефлекс?
Рефлекс как основа деятельности ЦНС.
Свойства нервных центров.
Тормозные процессы в ЦНС.
Принципы координационной деятельности мозга.
Рефлекс как основа деятельности ЦНС. Основой деятельности ЦНС является рефлекс. Огромный вклад в разработку рефлекторной теории внесли выдающиеся российские физиологи И. М. Сеченов, И. П. Павлов, А. А. Ухтомский, П. К. Анохин и ряд других ученых.
Рефлекс(от лат. – reflexus – повернутый назад, отраженный) рассматривается как закономерная ответная реакция организма на действие раздражителя, возбуждающего сенсорные рецепторы, которая осуществляется при обязательном участии ЦНС. Рефлекс – это возникновение, изменение или прекращение функциональной активности органов, тканей или целостного организма, осуществляемое при участии ЦНС в ответ на раздражение рецепторов организма.
Осуществление любого рефлекса требует обязательного выполнения четырех основных операций:
1. прием информации от рецептора;
2. расшифровка этой информации и программирование адекватного ответа;
3. реализация ответа путем передачи сигнала к исполнительным органам (мышцам, железам);
4. контроль за правильностью осуществления программы.
Последний этап замыкает круг непрерывной циркуляции нервных импульсов, формируя рефлекторное кольцо. Рецепторный контроль за ходом реализации запрограммированного ответа позволяет ЦНС своевременно зарегистрировать отклонения от намеченного плана и внести нужные поправки.
Структурной основой рефлекса является рефлекторная дуга (рефлекторный путь) – это совокупность образований, необходимых для осуществления рефлекса, или это последовательно соединенная цепь нейронов, по которой проводится возбуждение при осуществлении рефлекса. В случае безусловных рефлексов рефлекторная дуга формируется независимо от жизненного опыта индивидуума, т. е. ее становление генетически запрограммировано. Условно-рефлекторный процесс требует создания новой рефлекторной дуги на базе безусловного рефлекса. Иначе говоря, генетически запрограммированная цепь нейронов является лишь компонентом вновь создаваемой цепи нейронов.
Элементарная рефлекторная дуга безусловного рефлекса состоит из пяти основных звеньев. Она начинается рецепторами, которые трансформируют энергию внешнего раздражения в энергию нервного импульса. Импульс с участием афферентного нейрона поступает в центральную нервную систему (нервный центр), где непосредственно или опосредованно (через вставочный нейрон) передается на эфферентный нейрон, аксон которого передает команду в виде потенциала действия эффектору (мышечному волокну, секреторной клетке, другому нейрону). Нервный центр представляет собой функциональное объединение интернейронов, участвующих в организации рефлекторного акта.
Свойства нервных центров. Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований.
1. Одностороннее проведение возбуждения. В ЦНС – в ее нервных центрах, внутри рефлекторной дуги и нейронных цепей возбуждение, как правило, идет в одном направлении – от пресинаптической мембраны к постсинаптической, т. е. вдоль рефлекторной дуги от афферентного нейрона к эфферентному. Это свойство связано со свойствами синапсов.
2. Замедление проведения возбуждения в нервных центрах, или центральная задержка. Замедление проведения возбуждения по нервным центрам получило название центральной задержки. Она обусловлена медленным проведением нервных импульсов через синапсы, так как затрачивается время на следующие выделения медиатора из пресинаптических везикул, выброс его в синаптическую щель и генерация возбуждающего постсинаптического потенциала (ВПСП).
3. Суммация возбуждения и суммация торможения. Принято выделять два вида суммации – временную и пространственную. Временная, или последовательная, суммация проявляется в том, что в области постсинаптической мембраны происходит суммация следов возбуждения во времени, т. е. на нейроне в области его аксонного холмика происходит интеграция событий, разыгрывающихся на отдельных участках мембраны нейрона на определенном отрезке времени. Пространственная суммация возбуждения проявляется в суммировании на аксонном холмике нейрона постсинаптических потенциалов, которые возникают одновременно в различных точках этого нейрона в ответ на приходящие от других нейронов потенциалы действия. Даже если каждый из нейронов в отдельности вызывает лишь подпороговые ВПСП, при синхроннном их появлении они будут способны довести мембранный потенциал в области аксонного холмика нейрона до критического уровня деполяризации и тем самым вызывать возбуждение нейрона. Все сказанное в полной мере относится и к явлению суммации торможения.
4. Явление окклюзии (или закупорки) отражает эффект взаимодействия между собой двух импульсных потоков, при котором имеет место взаимное угнетение рефлекторных реакций. Суммарная ответная реакция (рефлекс), вызываемая одновременным воздействием двух потоков, меньше, чем сумма двух реакций, возникающих при действии каждого из этих двух потоков в отдельности. Согласно Ч. Шеррингтону, явление окклюзии объясняется перекрытием синаптических полей, образуемых афферентными звеньями двух взаимодействующих рефлексов.
5. Явление облегчения, которое по своему внешнему проявлению противоположно окклюзии. 0но проявляется в том, что при совместном раздражении рецептивных полей двух рефлексов наблюдается усиление реакций организма на действие двух раздражителей одновременно.
6. Трансформация ритма возбуждения. Это одно из свойств нейрона как компонента нейронной цепи, которое обнаруживается в процессе проведения возбуждения по нейронным цепям. Трансформация ритма возбуждения заключается в способности нейрона изменять ритм приходящих импульсов. Она проявляется и в противоположном феномене – частота приходящих к нейрону импульсов выше, чем частота генерации ПД при ответе нейрона на эти импульсы.
8. Высокая утомляемость нервных центров. Это свойство характерно для нейронных цепей, в том числе для рефлекторных дуг. С одной стороны, оно проявляется в том, что в нейронных цепях, как и в других многозвеньевых системах, может развиваться утомление, которое проявляется в постепенном снижении (вплоть до полного прекращения) рефлекторного ответа при продолжительном раздражении афферентных нейронов.
9. Тонус нервных центров. Для многих нейронных объединений, или нервных центров, характерна фоновая активность, т. е. генерация нервных импульсов с определенной частотой на протяжении длительного времени. Такая активность обусловлена не наличием в составе данного объединения нейрона-пейсмкера (фоновоактивного нейрона), а постоянным возбуждением афферентного нейрона благодаря непрерывному раздражению сенсорных рецепторов. Тонус нервных центров обеспечивает постоянную импульсацию к соответствующим периферическим системам, а также постоянное межцентральное взаимодействие.
10. Пластичность нервных центров – это их способность к перестройке функциональных свойств и, в определенной степени, функций под влиянием длительных внешних воздействий или при очаговых повреждениях мозга. Посттравматическая пластичность нейронных объединений выполняет компенсаторную (восстановительную) функцию, а пластичность, вызванная длительным афферентным раздражением, – приспособительную функцию. Например, для процесса обучения пластичность нейронных объединений является необходимым условием, т. е. его рабочим механизмом. В целом благодаря свойству пластичности нервный центр может существенно модифицировать течение рефлекторных реакций. Основным фундаментом, позволяющим реализовать свойство пластичности, очевидно, следует считать наличие у каждого нейрона в отдельности огромного числа синаптических связей, а также возможность изменения синтетических процессов внутри каждого нейрона.
Организация тормозных процессов в ЦНС. Вся деятельность нервной системы строится на двух основных процессах: на процессе возбуждения и на процессе торможения. Существует несколько различных типов торможения.
1. Реципрокное торможение (от лат. reciprocus – взаимный) было открыто английским физиологом Ч. Шеррингтоном и российским физиологом Н. Е. Введенским. Этот вид торможения основан на том, что одни и те же афферентные пути, через которые осуществляется возбуждение одной группы нервных клеток, обеспечивают через вставочные нейроны торможение других групп нейронов. Например, при возбуждении болевых рецепторов кожи конечности сигнал от ноцицепторов с участием афферентного нейрона поступает в спинной мозг, где переключается на альфа-мотонейрон мышц-сгибателей и одновременно на тормозной нейрон, который тормозит активность альфа-мотонейрона мышц-разгибателей. Реципрокное торможение характерно как для спинного мозга, так и для головного.
2. Возвратное торможение наблюдается в отношении альфа-мотонейронов спинного мозга. При возбуждении альфа-мотонейрона нервный импульс направляется к мышечным волокнам, возбуждая их. Одновременно по коллатерали, идущей к тормозному нейрону (клетка Реншоу), импульс возбуждает эту тормозную клетку, которая в свою очередь вызывает торможение возбужденного ранее альфа-мотонейрона. Таким образом, альфа-мотонейрон, активируясь, через систему тормозного нейрона сам себя (возвратно, или антидромно) затормаживает.
3. Тоническоеторможение отражает наличие постоянного тормозного влияния одной структуры на другую. Примером такого постоянного тормозного влияния являются тормозные нейроны коры больших полушарий, которые угнетают нейроны ретикулярной формации ствола мозга, нейроны таламуса и лимбической системы.
4. Общее центральное торможение – это нервный процесс, который развивается при любой рефлекторной деятельности. Он захватывает почти всю ЦНС, включая центры головного мозга. Такое торможение проявляется раньше, чем возникнет какая-либо двигательная реакция.
4. Пессимальноеторможение развивается в возбуждающих синапсах в результате сильной и длительной деполяризации постсинаптической мембраны приходящими сюда высокочастотными импульсными потоками, что приводит к снижению возбудимости (повышению порога возбуждения) в постсинаптической мембране, делающим ее неактивной.
5. Торможение вслед за возбуждением развивается достаточно часто, так как возникает всякий раз на фоне следовой гиперполяризации мембраны нейрона после очередного его возбуждения. Для него характерен сравнительно кратковременный период существования, так как он определяется лабильностью нейрона, т. е. скоростью восстановления исходного уровня мембранного потенциала после генерации очередного потенциала действия.
Принципы координационной деятельности мозга. ЦНС состоит из огромного числа нейронов, которые образуют различные по уровню сложности и задачам нейронные объединения – нервные центры, нейронные цепи, рефлекторные дуги, нейронные ансамбли, нейронные сети. Часть этих объединений возникает благодаря генетической информации, а часть – в процессе индивидуального развития на основе некоторых принципов, которые также заложены в генетической программе. Эти принципы позволяют интегрировать деятельность всех отдельных нейронных объединений ради выполнения наиболее важных задач организма, обеспечивающих ему выживание в реальных условиях существования.
Принцип иррадиации, или дивергенции, возбуждения ЦНС. Иррадиация возбуждения (от лат. irradio, озарять, освещать) – это распространение процесса возбуждения из одного участка ЦНС в другой. Каждый нейрон за счет многочисленных ответвлений (дивергенции), заканчивающихся синапсами, и большого числа вставочных нейронов связан со многими другими нейронами. Поэтому нервные импульсы от одного нейрона могут быть направлены к тысяче других нейронов. Процесс иррадиации возбуждения регулируется различными механизмами. Он может быть усилен, например, за счет активации ретикулярной формации ствола мозга. С другой стороны, процесс иррадиации ограничивается, и это важное действие осуществляется с участием многочисленных тормозных нейронов. Например, в спинном мозге ограничение иррадиации осуществляется благодаря механизму возвратного торможения с помощью специальных тормозных интернейронов – клеток Реншоу. Иррадиация возбуждения играет исключительно важную роль, так как позволяет обмениваться многочисленными потоками информации различным структурам мозга. Именно за счет иррадиации происходит обмен информацией между первой и второй сигнальными системами (элективная иррадиация), что существенно увеличивает возможности высшей нервной деятельности человека. Благодаря иррадиации возбуждения осуществляется интеграция деятельности правого и левого полушарий.
Принцип концентрации возбуждения. Возбуждение и торможение могут либо иррадиировать, либо концентрироваться. Концентрация – это явление, противоположное иррадиации. Оно возникает в тот момент, когда иррадиация достигает определенной границы, после чего распространение возбуждения или торможения идет в обратном направлении, т. е. в исходный пункт. Если нейроны находятся в заторможенном состоянии, это препятствует распространению на них процесса возбуждения, и наоборот, если они находятся в состоянии возбуждения, торможением они охватываются труднее. На скорость иррадиации и концентрации нервных процессов оказывает влияние и тип нервной системы, ее индивидуальные особенности.
Принцип индукции нервных процессов. Индукция отражает проявление процессов иррадиации возбуждения и торможения в коре больших полушарий. Принято считать, что индукция – (от лат. Inductio – введение, наведение) обозначает возникновение нервного процесса, противоположного по знаку процессу, вызванному условным раздражителем (положительным или отрицательным, т. е. тормозным). Одновременная индукция заключается в том, что формирование в каком-либо центре коры больших полушарий концентрированного возбуждения вызывает в прилежащих к этому центру зонах торможение (отрицательная одновременная индукция), а такое же концентрированное торможение вызывает в этих зонах возбуждение (положительная одновременная индукция). Таким образом, при одновременной индукции нервный процесс вызывает в другом участке коры процесс, противоположный по знаку (процесс возбуждения вызывает процесс торможения и наоборот), а при последовательной индукции происходит смена противоположных нервных процессов в одном и том же участке.
Принцип субординации, или соподчинения, также относится к категории важнейших принципов организации работы мозга. Согласно этому принципу, деятельность нижележащих отделов мозга контролируется и управляется вышележащими отделами ЦНС. Например, в двигательных системах мозга и в вегетативной нервной системе имеются нейронные объединения (нервные центры), расположенные в спинном мозге или в стволе мозга, которые подчиняются деятельности нейронных объединений (нервных центров), находящихся в гипоталамусе, таламусе, мозжечке, базальных ядрах и коре больших полушарий.
Принцип обратной связи (обратной афферентации) и копий эфферентаций. Согласно этому принципу, для точной координации деятельности различных нейронных объединений (нервных центров, рефлекторных дуг) необходима оптимальная по объему информация о результатах действия. Она поступает в мозг по сенсорным каналам. Отсутствие такой информации приводит к дезинтеграции деятельности мозга. Особенно наглядна роль обратной афферентации при реализации двигательной активности – нарушение проприоцептивной чувствительности, как правило, препятствует выполнению точных движений, а также нарушает возможность формирования и сохранения адекватной для данного движения позы.
Принцип реципрокности (сопряжения) возбуждения и торможения на уровне спинного мозга реализуется с участием реципрокного торможения, благодаря которому возникают безусловные двигательные реципрокные рефлексы. Реципрокное торможение осуществляется по механизму постсинаптического торможения, которое возникает с участием специальных вставочных тормозных нейронов.
Вопросы для самоконтроля
1. Из каких звеньев состоит рефлекторная дуга?
2. Что представляет собой нервный центр?
3. Чем обусловлена центральная задержка в нервных центрах?
4. Какие существуют типы суммации возбуждения?
5. В чем заключается явление окклюзии?
6. Чем объясняется высокая утомляемость нервных центров?
7. Какую роль в функционировани ЦНС играет пластичность нервных центров?
8. Какие существуют типы тормозных процессов в ЦНС?
9. Какими механизмами регулируется процесс иррадиации возбуждения?
10. От чего зависит скорость иррадиации и концентрации нервных процессов?
11. Чем отличается одновременная индукции нервных процессов от последовательной индукции?
12. Кем было открыто явление конвергенции и в чем оно заключается?
13. Какое значение имеет доминанта в жизнедеятельности человека?
14. Какие существуют принципы кодирования информации в нервной системе?
15. В чем заключается принцип субординации?
ОСНОВНАЯ ЛИТЕРАТУРА
1. Бак З. Химическая передача нервного импульса. М.: Мир, 1977.
2. Буреш Я., Бурешова У., Хьюстон Д. П. Методики и основные эксперименты по изучению мозга и поведения. Пер. с. англ. М.: Высшая школа, 1991.
3. Недоспасов В. О. Физиология центральной нервной системы. Учебник для студентов высших учебных заведений. М.: ООО УМК Психология, 2002.
4. Основы физиологии человека. Под ред. Б. И. Ткаченко, СПБ., 1994.
5. Фабер Д. А., Семенова Л. К., Алферова В. В. Структурнофункциональная организация развивающегося мозга. Л, 1990.
6. Физиология человека. Под ред. Н. А Агаджаняна. СПб.: Сотис, 1998.
7. Физиология центральной нервной системы. Под ред. Г. А. Кураева. РостовнаДону: Феникс, 2000.
8. Физиология человека. Под ред. В. М. Смирнова, М.: Медицина, 2001.
9. Циркин В. И., Трухина С. И. Физиологические основы поведения человека. М.: Медицинская книга, 2001.
10. Шульговский В. В. Основы нейрофизиологии: Учебное пособие для студентов вузов. М.: Аспект Пресс, 2000.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
1. Батуев А. С. Высшая нервная деятельность. М.: Высшая школа, 1991.
2. Данилова Н. Н., Крылова А. Л.Физиология высшей нервной деятельности. М., 1989.
3. Кендел Э. Клеточные основы поведения, М.: Мир, 1980.
4. Физиология человека. Под ред. В. М. Покровского. Т. 2. М.: Медицина, 1998.
5. Самусев Р. П. Анатомия человека. М., 1995.
6. Симонов П. В.: Мотивированный мозг. М., 1987
7. Данилова Н. Н. Психофизиология: Учебник для вузов. М., 1998.
8. Психофизиология. Под. ред. Александрова Ю. И. СПб.: Питер, 2001.
9. Общий курс физиологии человека и животных. Под ред. А. Д. Ноздрачева. Т. 1. М.: Высшая школа, 1991.
10. Физиология. Основы и функциональные системы. Под ред. К. В. Судакова. Т. 1. М.: Медицина, 2000.
11. Физиология человека. Под ред. Г. И. Косицкого, М.: Медицина, 1985.
12. Мозг, разум и поведение. Под ред. Блума, М.1989.
Учебное издание
Пивоварчик Маргарита Владимировна
Читайте также:
