
Процесс поляризации нервных клеток
В любом учебнике, где есть раздел о нервной системе, можно посмотреть, как устроена нервная система, из каких отделов она состоит. Это объясняют и в учебниках по анатомии, и в учебниках по физиологии. Да и на этом сайте есть подобные материалы: смотри, например, раздел Спинной мозг.

Но вот как всё это работает?
Вот в чём главный вопрос!
Видеофильм о работе нейрона демонстрирует внешние события, отражающие работу нейрона.
Хорошо известно, что для своей работы нервные клетки пользуются электричеством и обмениваются именно электрическими сигналами в виде нервных импульсов. И для начала неплохо было бы узнать, откуда нервные нервные клетки берут для этого электричество!
На этот вопрос - о возникновении электрических зарядов в нервных клетках - даётся ответ в разделе: _1 Мембранный потенциал покоя
После того, как мы поняли, откуда на мембране берутся электрические потенциалы, можно попытаться понять, в чём же заключается работа нервных клеток. Как они пользуются этими электрическими потенциалами и электрически заряженными частицами - ионами?
Нервная система очень сложна. И нервные клетки, из которых она состоит, - это тоже непростые образования!
И всё же можно выделить самое главное в их работе. Мы попробуем разобраться, чем же они занимаются и что они делают главным образом.

Говоря "нервные клетки", обычно имеют в виду нейроны. И это несмотря на то, что к нервным клеткам относятся ещё и глиальные клетки, или глия (смотри: Нейроны и глия в гиппокампе), и клеточные сенсорные рецепторы. Но их всё же следует рассматривать отдельно. Так что мы тоже вслед за остальными будем называть именно нейроны "нервными клетками", используя эти термины как синонимы.
Если сформулировать задачи нейрона кратко и образно, то получится всего 3 главные задачи.
1. Восприятие возбуждения.
Происходит восприятие воздействия со стороны других нейронов или сенсорных рецепторов. Воздействие - это химический выброс медиаторов и их связывание с мембранными рецепторами нейрона. Нейрон должен воспринять подходящие (т.е. адекватные) для него химические сигналы (медиаторы) своими воспринимающими молекулярными мембранными рецепторами. Как правило, эти рецепторы сосредоточены в воспринимающих отделах его синапсов - на постсинаптических (иногда их называют также субсинаптическими) участках мембраны. Воспринимающий отдел нейрона - дендриты с синапсами, а точнее - с постсинаптическими мембранами. Результат восприятия возбуждения - возникновение на мембране воспринимающего нейрона локального электрического потенциала.
2. Проведение возбуждения.
Проведение возбуждения - это движение возбуждения от места его возникновения до места передачи, т.е. от пресинаптического окончания дендрита до постсинаптического окончания аксона. Для этого требуется рождение нервного импульса на основе локального возбуждающего потенциала и движение этого нервного импульса по мембране нейрона до постсинаптического окончания аксона, где будет происходить химический выброс медиатора. Таким образом, воздействие на нейрон происходит на одном его участке (обычно на пресинаптическом окончании дендрита), но за счёт проведения это возбуждение оказывается на другом конце нейрона - на окончаниях аксона.
3. Передача возбуждения.
Передача возбуждения - это химический выброс медиатора из окончаний аксона. За счёт выброса медиатора осуществляется воздействие со стороны одного нейрона на другой нейрон (или иные клетки). Нейрон должен оказать химическое воздействие на свои мишени: это другие нейроны, мышечные клетки или железистые клетки. Для этого он выделяет химические управляющие вещества (медиаторы или модуляторы) из окончаний своего передающего отростка - аксона. Передающие окончания заканчиваются пресинаптическими мембранами, через которые нейрон и выделяет свои управляющие вещества.
Итак, нейрон должен воспринимать химические воздействия на своей воспринимающей части, проводить возбуждение в виде нервного импульса и оказывать химическое воздействие своими управляющими веществами на свои мишени с помощью своей передающей части. Для связи между собой разных частей нейрона служат нервные импульсы, пробегающие от воспринимающей части нейрона (дендритных окончаний) к его передающей части (аксонным окончаниям).
Таким образом, нейрон должен выделять свои управляющие вещества в нужное время, в нужном месте, в нужном количестве и нужном качестве . И сам нейрон, в свою очередь, должен воспринимать управляющие воздействия, направленные на него. Если что-то из перечисленного не получится, то нужного воздействия от такого нейрона или вообще не будет, или оно будет не эффективным.
Нервные клетки имеют определённый набор ключевых реакций:
1. Формирование и поддержание мембранного потенциала покоя . Этим обеспечивается возбудимость нейронов.
2. Формирование локальных потенциалов . Этим обеспечивается подпороговое возбуждение нейронов.
3. Генерация потенциала действия и нервного импульса . Этим обеспечивается переход от аналогового к цифровому кодированию информации нейронами.
4. Секреция нейротрансмиттеров (медиаторой и модуляторов). Этим обеспечивается передача возбуждения на другие клетки.
5. Изменение своего внутриклеточного обмена и биосинтеза. Этим обеспечивается пластичность и адаптация нейронов.
6. Изменение своих клеточных структур и свойств. Этим обеспечивается пластичность нейронов.
7 Формирование новых структур, например, новых синапсов. Этим обеспечивается пластичность нервных связей.
8. Движение нервного импульса по мембране нейрона. Этим обеспечивается проведение возбуждения нейронами.
Мембранный потенциал покоя, т.е. разность электрических потенциалов между внутренней и наружной сторонами мембраны, нейроны поддерживают на уровне в среднем -70 мВ и тратят на это до 75% всей своей энергии, полученной за счёт питания.
Подпороговое изменение электрического состояния своей мембраны (локальный потенциал). Это проявляется в увеличении возбудимости или, наоборот, в уменьшении возбудимости (торможении).
Деполяризация - это уменьшение электроотрицательности нервной клетки.
Можно сказать, что электрический потенциал мембраны при этом "ползёт вверх", в сторону нуля.
Деполяризация означает возбуждение клетки, деполяризованная клетка - более возбудима, т.е. чувствительна к возбуждению.
Как правило, деполяризацию и улучшение возбудимости нервной клетки производит входящий в неё натрий (положительно заряженные ионы натрия). Хочешь вызвать деполяризацию - впусти в клетку натрий!
Важно отметить, что термин "деполяризация" означает именно утрату поляризации. При этом электрический заряд мембраны уменьшается и стремится к нулю. Клетка с "нулевым" зарядом не поляризована. Если же заряд становится вместо отрицательного положительным, то это тоже означает поляризацию клетки. Только уже положительную. К сожалению, частенько из учебников можно ошибочно сделать вывод, что деполяризация продолжается даже выше нуля, приобретая положительное значение. Это неверно!
Гиперполяризация - это увеличение электроотрицательности нервной клетки.
Термин "гиперполяризация" означает "сверхполяризацию". При этом электрический заряд мембраны "ползёт вниз", т.е. удаляется от нуля.
Гиперполяризация означает торможение клетки, она становится менее возбудимой, т.е. слабо чувствительной к возбуждению.
Как правило, гиперполяризацию можно вызвать двумя способами:
1) запустить в нервную клетку отрицательно заряженные ионы хлора,
2) выпустить из нервной клетки дополнительное количество положительно заряженных ионов калия.
Хочешь вызвать гиперполяризацию и ухудшить возбудимость нервной клетки - запусти в неё хлор или выпусти из неё калий!
Порождение нервного импульса (потенциала действия). Смотри: Потенциал действия и нервный импульс
Когда деполяризация мембраны достигает определённого критического уровня, то происходит резкий скачок электрического напряжения от отрицательного к положительному и обратно к отрицательному. В этом процессе участвуют ионные каналы мембраны и перемещение ионов натрия и калия.
Важно отметить, что после того как этот процесс начался в одном месте, он, как пожар, охватит всю мембрану нейрона и пробежит по всем его отросткам. Это и есть нервный импульс - распространяющееся возбуждение. Часто в качестве синонима для нервного импульса используют термин "потенциал действия". Однако следует помнить, что понятие "потенциал действия" относится только к электрическому процессу, т.е. отражает лишь часть более общего понятия, такого как "нервный импульс".
Нервный импульс - это волна взаимосвязанных структурных, химических и электрических процессов, пробегающая по мембране нейрона.
Потенциал действия - это резкое скачкообразное изменение электрического заряда мембраны с отрицательного на положительный и обратно на отрицательный.
Выброс специального химического вещества, влияющего на состояние других нервных клеток. Эти вещества называются нейротрансмиттерами (нейромедиаторами и нейромодуляторами). Смотри: Медиаторы и модуляторы. Пожалуй, это и есть главная функция нейрона - выбрасывать управляющие вещества для воздействия на свои мишени.
Изменение обмена веществ в данной клетке в ответ на полученный химический сигнал.
Длительное изменение свойств этой клетки или её отдельных структур. Это очень важная способность живых структур, отличающая их от технических устройств! Нейроны могут видоизменять молекулы своих рабочих белков с помощью их фосфорилирования, т.е. присоединения к ним фосфорных остатков. А также могут синтезировать новые молекулярные рецепторы и встраивать их в свою мембрану.
Это явление называется "пластическими перестройками", а сама способность к таким перестройкам - "пластичность". Именно пластичность нейронов и, в частности, пластичность синапсов, обеспечивает нервной системе такие важнейшие свойства, как память, научение, образование условных рефлексов.
Проживая свою клеточную жизнь, нейроны постоянно поддерживают себя в электрически заряженном состоянии. Как они это делают, рассматривается нами в разделе: 2_1 Мембранный потенциал покоя. И при этом время от времени они изменяют свои заряды, а также время от времени делают выбросы биологически активных веществ - трансмиттеров (медиаторов и модуляторов).
Итак, на фоне поддержания своего постоянного электрического заряда (потенциала покоя) нейроны претерпевают деполяризацию и гиперполяризацию, порождают нервные импульсы (потенциалы действия), выделяют нейротрансмиттеры (медиаторы и модуляторы), изменяют свой обмен веществ (метаболизм), или же на долгий срок изменяют свои свойства, используя свою способность к пластичности.
В этом и заключается их клеточная работа на благо всего организма.
А теперь я раскрою вам самую главную тайну о работе нейронов.
Если раньше считалось, что главная работа нейронов - это производство нервных импульсов, то теперь акценты сместились.
Главная работа нейронов - это производство и выделение химических управляющих веществ - нейротрансмиттеров (медиаторов и модуляторов). Именно с помощью своих нейротрансмиттеров нейроны управляют друг другом и всем организмом!
А электрохимические нервные импульсы лишь согласуют эту химическую активность нейронов во времени.
Вот какую великую тайну работы нервных клеток вы постигли вместе со мной.
Вся нервная деятельность успешно функционирует благодаря чередованию фаз покоя и возбудимости. Сбои в системе поляризации нарушают электрическую проводимость волокон. Но кроме нервных волокон есть и другие возбудимые ткани — эндокринная и мышечная.
Но мы рассмотрим особенности проводимых тканей, и на примере процесса возбуждения органических клеток расскажем о значении критического уровня деполяризации. Физиология нервной деятельности тесно связана с показателями электрического заряда внутри и снаружи нервной клетки.
Если один электрод присоединить к внешней оболочке аксона, а другой - к его внутренней части, то видна налицо разность потенциалов. Электрическая активность нервных проводящих путей основана на этой разности.
Что такое потенциал покоя и потенциал действия?
Все клетки нервной системы поляризованы, то есть имеют разный электрический заряд внутри и снаружи специальной мембраны. Нервная клетка всегда имеет свою липопротеиновую мембрану, имеющую функцию биоэлектрического изолятора. Благодаря мембранам создается потенциал покоя в клетке, который необходим для последующей активации.
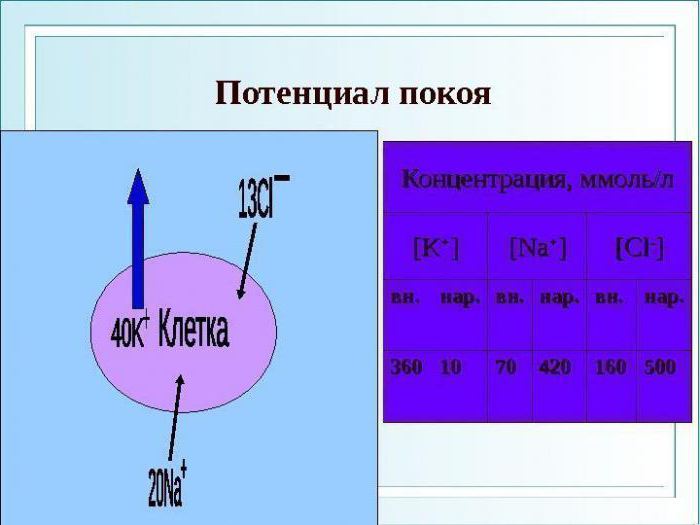
Потенциал покоя поддерживается путем переноса ионов. Выход ионов калия и вход хлора увеличивает потенциал мембранного покоя.
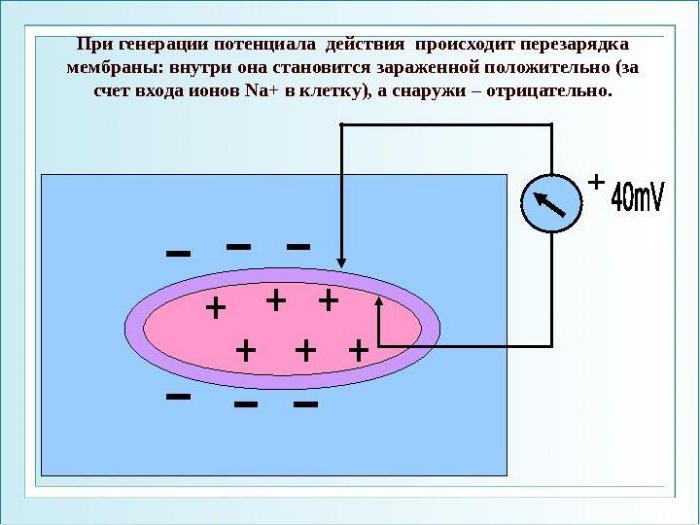
Потенциал действия накапливается в фазе деполяризации, то есть подъема электрического заряда.
Фазы потенциала действия. Физиология
Итак, деполяризация в физиологии — это снижение мембранного потенциала. Деполяризация основа возникновения возбудимости, то есть потенциала действия для нервной клетки. При достижении критического уровня деполяризации никакой, даже сильный раздражитель не способен вызвать реакции нервных клеток. Натрия при этом очень много внутри аксона.
Сразу после этой стадии следует фаза относительной возбудимости. Ответ уже возможен, но лишь на сильный сигнал-раздражитель. Относительная возбудимость медленно переходит в фазу экзальтации. Что такое экзальтация? Это пик возбудимости тканей.
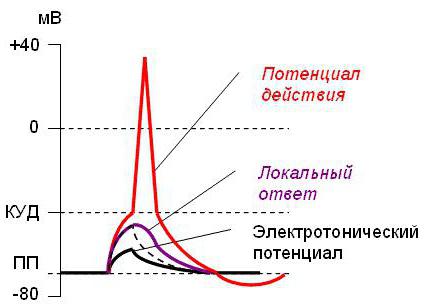
Все это время натриевые каналы активации закрыты. А их открытие произойдет, только когда нервное волокно разрядится. Реполяризация нужна для восстановления отрицательного заряда внутри волокна.
Что означает критический уровень деполяризации (КУД)?
Итак, возбудимость, это в физиологии способность клетки или ткани отреагировать на раздражитель и генерировать какой-то импульс. Как мы выяснили, для работы клеткам нужен определенный заряд — поляризация. Нарастание заряда от минуса к плюсу называется деполяризацией.
После деполяризации всегда идет реполяризация. Заряд внутри после фазы возбуждения снова должен стать отрицательным, чтобы клетка могла подготовиться к следующей реакции.
Когда показания вольтметра зафиксированы на отметке 80 - это фаза покоя. Она наступает после окончания реполяризации, а если прибор показывает положительное значение (больше 0), значит, обратная реполяризации фаза, приближается к максимальному уровню — критическому уровню деполяризации.
Как передаются импульсы от нервных клеток к мышцам?
Электрические импульсы, возникшие при возбуждении мембраны, передаются по нервным волокнам с большой скоростью. Скорость сигнала объясняется строение аксона. Аксон частично обволакивается облочкой. А между участками с миелином находятся перехваты Ранвье.
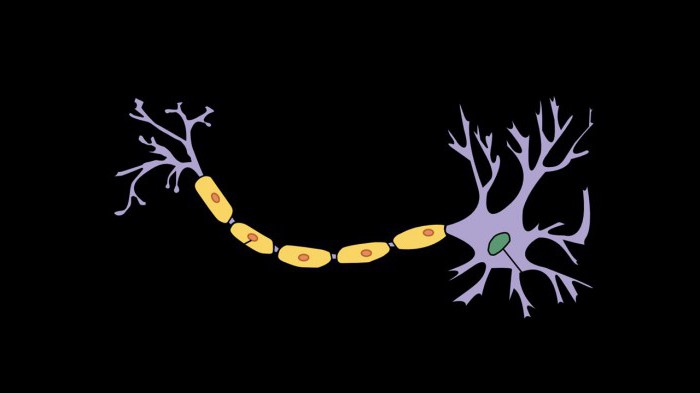
Благодаря такому устройству нервного волокна положительный заряд чередуется с отрицательным, и деполяризационный ток практически единовременно распространяется вдоль всей длины аксона. Сигнал о сокращении доходит до мышцы в доли секунды. Такой показатель, как критический уровень деполяризации мембраны означает ту отметку, при которой достигается пиковый потенциал действия. После сокращения мышцы вдоль всего аксона запускается уже реполяризация.
Что происходит при деполяризации?
Что значит такой показатель, как критический уровень деполяризации? Это в физиологии означает, что нервные клетки уже готовы к работе. Исправная работа целого органа зависит от нормальной, своевременной смены фаз потенциала действия.
Критический уровень (КУД) равен приблизительно 40–50 Мв. В это время электрическое поле вокруг мембраны уменьшается. Степень поляризации напрямую зависит от того, сколько натриевых каналов клетки открыто. Клетка в это время еще не готова к ответу, но собирает электрический потенциал. Этот период имеет название абсолютная рефрактерность. Длится фаза всего 0,004 с в нервных клетках, а в кардиомиоцитах - 0,004 с.
После прохождения критического уровня деполяризации наступает супервозбудимость. Нервные клетки могут дать ответ даже на действие подпорогового раздражителя, то есть относительно слабого воздействие среды.
Функции натриевых и калиевых каналов
Итак, важный участник процессов деполяризации и реполяризации белковый ионовый канал. Разберемся, что подразумевает под собой это понятие. Ионные каналы — это находящиеся внутри плазменной оболочки белковые макромолекулы. Когда они открыты, через них могут проходить ионны неорганического происхождения. Белковые каналы имеют фильтр. Через натриевый проток проходит только натрий, через калиевый — только этот элемент.
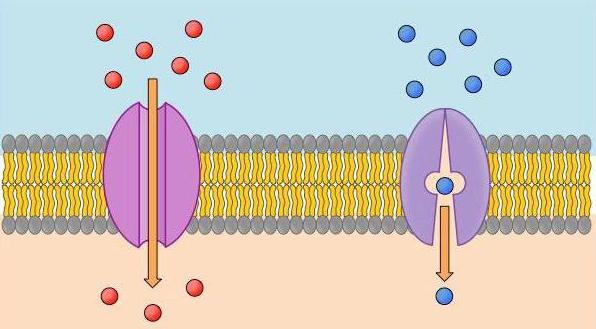
Эти электроуправляемые каналы имеют двое ворот: одни активационные, обладают свойством пропускать ионы, другие инактивационные. В то время, когда мембранный потенциал покоя равен -90 мВ, ворота закрыты, но при начале деполяризации, натриевые каналы медленно открываются. Увеличение потенциала приводит к резкому закрытию створок протока.
Фактором, который влияет на активацию каналов, является возбудимость мембраны клетки. Под действием электрической возбудимости и запускаются 2 вида ионовых рецепторов:
- запускается действие лиганд рецепторов — для хемозависимых каналов;
- электрический сигнал подается для электроуправляемых каналов.
При достижении критического уровня деполяризации мембраны клетки рецепторы дают сигнал о том, что все натриевые каналы нужно закрыть, а калиевые начинают открываться.
Натриево-калиевый насос
Процессы передачи импульса возбуждения везде проходят благодаря электрической поляризации, осуществляемой за счет движения ионов натрия и калия. Движение элементов происходит на основе принципа активного транспорта ионов - 3 Na + внутрь и 2 К + наружу. Этот механизм обмена называется натриево-калиевым насосом.
Деполяризация кардиомиоцитов. Фазы сокращения сердца
Сердечные циклы сокращений также связаны с электрической деполяризацией проводимых путей. Сигнал о сокращении всегда исходит от СА-клеток, находящихся в правом предсердии, и распространяется по проводящим путям Гисса в пучок Тореля и Бахмана в левое предсердие. Правые и левые отростки пучка Гисса передают сигнал в желудочки сердца.
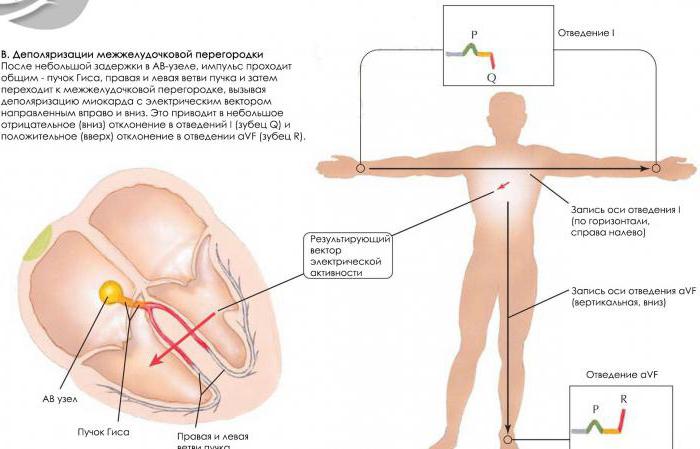
Нервные клетки быстрее деполяризуются и переносят сигнал благодаря наличию миелиновой оболочки, но мышечные ткани также постепенно деполяризуются. То есть их заряд из отрицательного превращается в положительный. Эта фаза сердечного цикла называется диастолой. Все клетки тут соединены между собой и действуют как один комплекс, поскольку работа сердца должна быть максимально скоординирована.
Когда наступает критический уровень деполяризации стенок правого и левого желудочков, генерируется выброс энергии — происходит сокращение сердца. Затем все клетки реполяризуются и готовятся к новому сокращению.
Депрессия Вериго
В 1889 году описано явление в физиологии, которое называется католической депрессией Вериго. Критический уровень деполяризации — это уровень деполяризации, при котором все натриевые каналы уже инактивированы, а вместо них работают калиевые. Если степень тока еще больше увеличивается, тогда значительно снижается возбудимость нервного волокна. А критический уровень деполяризации при действии раздражителей зашкаливает.
Во время депрессии Вериго скорость проведения возбуждения понижается, и, наконец, совсем спадает. Клетка начинает адаптироваться за счет изменения функциональных особенностей.
Адаптационный механизм
Бывает, при некоторых условиях деполяризующий ток долго не переключается. Это свойственно сенсорным волокнам. Постепенное длительное повышение такого тока сверх нормы в 50 мВ приводит к увеличению частоты электронных импульсов.
В ответ на такие сигналы повышается проводимость калиевой мембраны. Активируются более медленные каналы. В итоге возникает способность нервной ткани к повторным ответам. Это называется адаптацией нервных волокон.
При адаптации вместо большого количества коротких сигналов клетки начинают аккумулировать и отдавать одиночный сильный потенциал. А интервалы между двумя реакциями увеличиваются.
При образовании потенциалов действия (спайное) в химических синапсах между нейронами происходит высвобождение медиаторов из пузырьков пресинаптической мембраны. Медиатор проходит через синаптическую щель и активирует рецепторы, встроенные в постсинаптическую мембрану нейрона-мишени. Рецепторы, в свою очередь, активируют медиаторозависимые ионные каналы, изменяющие поляризацию нейрона-мишени. При смещении показателей мембранного потенциала от уровня потенциала покоя (-70 мВ) в отрицательную сторону (например, — 80 мВ и более) происходит гиперполяризация мембраны, а при смещении в положительную сторону — деполяризация.
а) Электротонические потенциалы. Первоначальный ответ нейрона-мишени на воздействие возбуждающих импульсов заключается в формировании локальных ступенчатых электротонических потенциалов. Положительные злектротонические потенциалы обычно возникают в мультииолярных нейронах в результате деполяризации, обусловленной активацией медиаторозависимых ионных каналов. При низкочастотном стимулирующем воздействии по дендритам распространяются небольшие волны деполяризации, угасающие в течение 2-3 мс.
При увеличении частоты воздействия происходит ступенчатая временная суммация возбуждения, в результате чего образуется прогрессивно увеличивающаяся волна, которая распространяется на поверхность сомы. Одновременное схождение в области сомы волн возбуждения, распространяющихся по разным дендритам, приводит к пространственной суммации возбуждения. При деполяризации приблизительно на 15 мВ (и значении потенциала -55 мВ) происходит открытие потенциалозависимых ионных каналов в наиболее чувствительной области нейрона — триггерной точке (триггерной области), располагающейся в начальном сегменте аксона. При достижении уровня деполяризации (генераторного потенциала) пороговых значений, т.е. потенциала, способного запустить открытие потенциалозависимых ионных каналов, происходит формирование потенциала действия.
В триггерной зоне чувствительных нейронов черешнях и спинномозговых нервов формируется так называемый рецепторный потенциал. В этой области расположено большое количество ионных каналов, активирующихся под воздействием специфических чувствительных стимулов и вызывающих ступенчатое распространение волны деполяризации.
Определение триггерной области миелинизированных нервных волокон не вызывает затруднений: у мультиполярных нейронов эта область граничит с первым миелиновым сегментом с его проксимальной стороны, а у периферических чувствительных нейронов — с последним миелиновым сегментом с его дистальной стороны.
Тормозные (гиперполяризующие) постсинаптические потенциалы возникают при открытии ионных каналов, генерирующих выходящий ток (например, калиевых каналов). Распространение тормозных потенциалов также является угасающим.
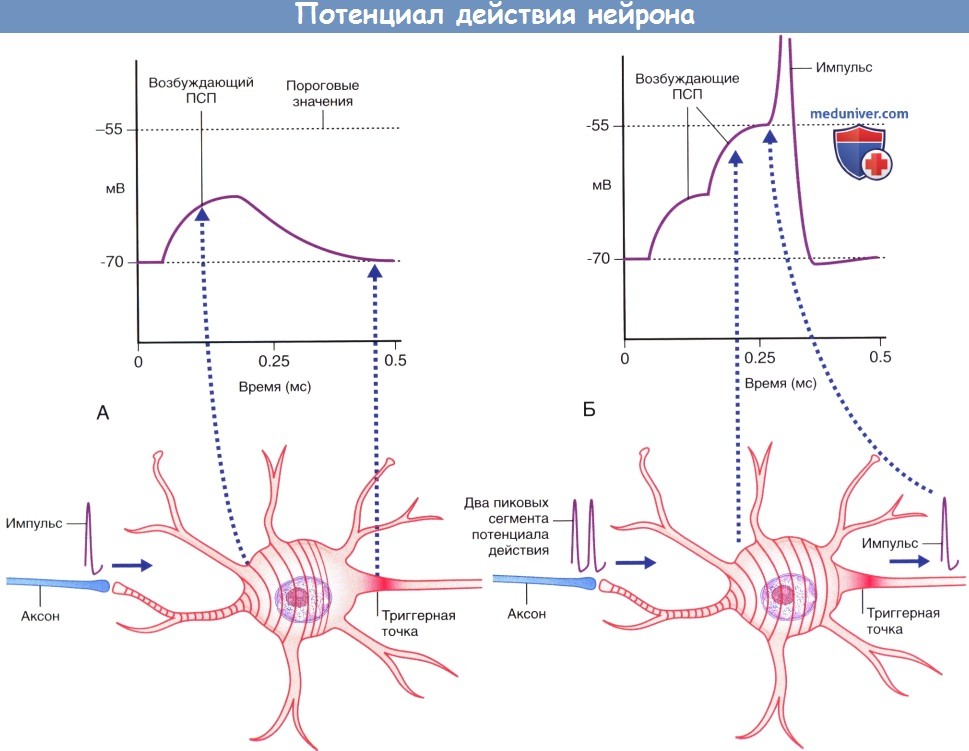
Временная суммация.
(А) Аксон чувствительного нейрона (выделен голубым цветом) передает единичный импульс двигательному нейрону,
вызывающий возбуждающему постсинаптический потенциал (ПСП), значения которого не достигают пороговых величин, вследствие чего этот потенциал затухает.
(Б) Аксон чувствительного нейрона передает два импульса.
Происходит временная суммация импульсов с достижением в начальном сегменте аксона пороговых значений возбуждения,
что приводит к формированию потенциала действия, который в дальнейшем распространяется по аксону двигательного нейрона. 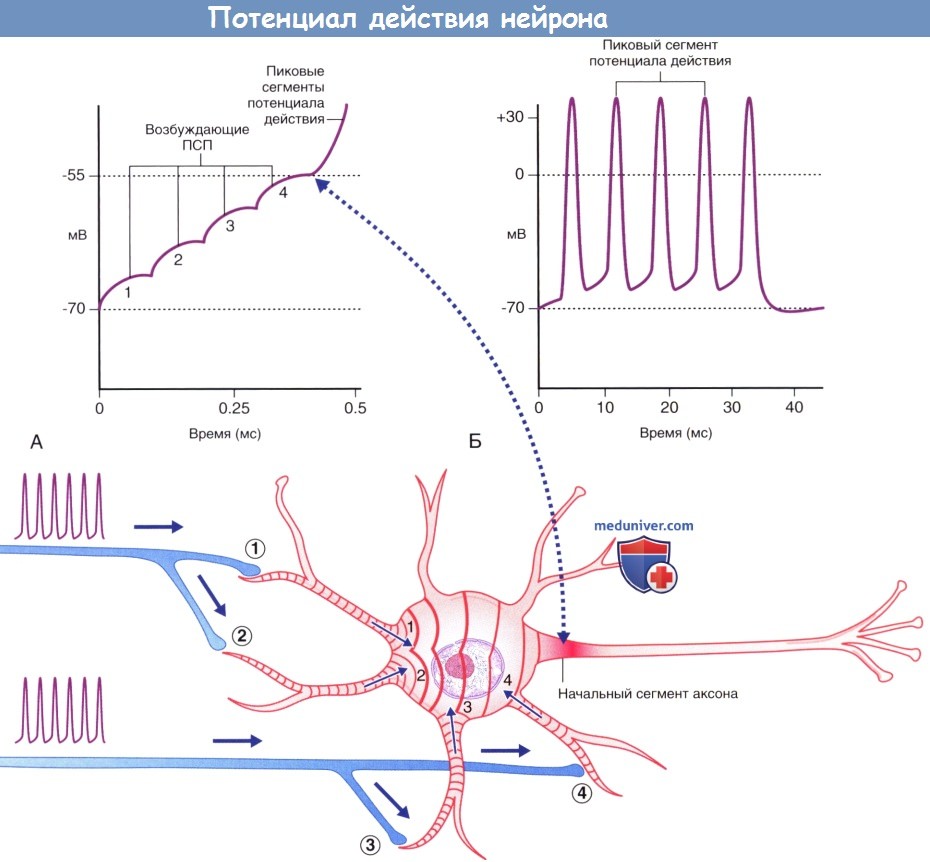
(А) Ступенчатая суммация возбуждающих постсинаптических потенциалов (ПСП), за счет которой происходит формирование пикового потенциала. Пунктирной линией указана область, соответствующая графику А.
(Б) Генераторные потенциалы достаточной силы формируют множественные пиковые потенциалы. 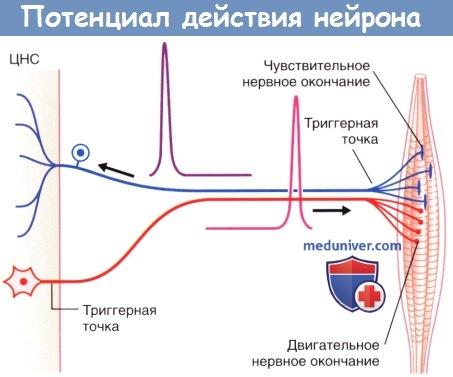
Потенциалы действия двигательных и чувствительных нейронов скелетной мускулатуры. ЦНС — центральная нервная система. 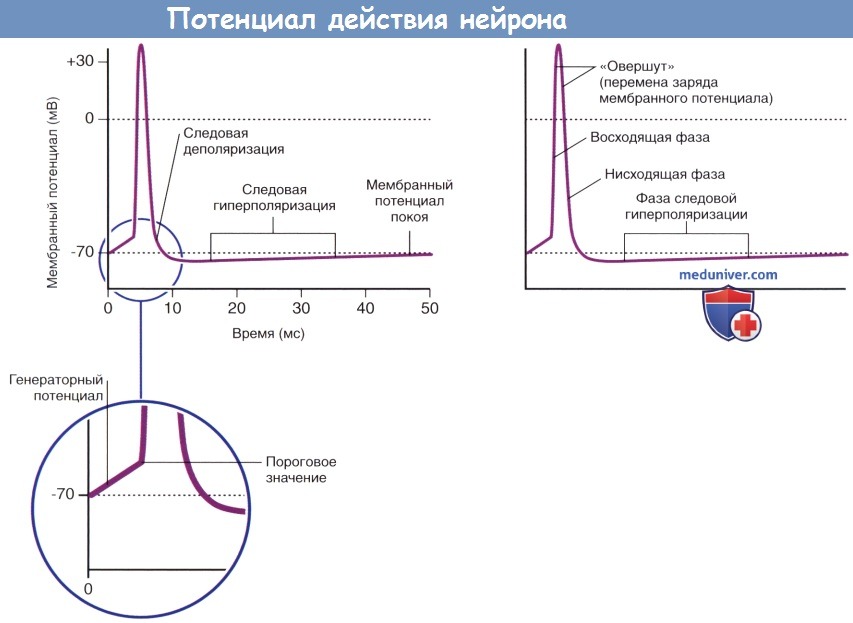
Основные характеристики потенциала действия.
Возникновение фазы следовой гиперполяризации объясняют тем, что после полной инактивации потенциалозависимых натриевых каналов многие калиевые каналы остаются открытыми. Восстановление ионных градиентов обеспечивает натрий-калиевый насос.
В ходе подробного изучения натриевых каналов было обнаружено, что их регуляция осуществляется двумя путями. В состоянии покоя (-70 мВ) активационные ворота, расположенные в средней части натриевых и калиевых каналов, закрыты. Первыми на пороговое раздражение реагируют натриевые каналы: происходят открытие активационных ворот и быстрый выход ионов Na + в клетку по концентрационному и электрическому градиентам. При достижении потенциалом действия пиковых значений (+35 мВ) происходит открытие калиевых каналов, а натриевые каналы закрываются инактивационными воротами, представленными глобулярным белком. При реполяризации и восстановлении мембранного потенциала (-70 мВ) активационные натриевые ворота закрываются, а затем вновь открываются инактивационные натриевые ворота, тем самым приводя натриевые каналы в исходное состояние.
Работа потенциалозависимых калиевых каналов регулирует одноворотный механизм: этими каналами управляют только активационные ворота.
Во время фазы нарастания и в начале фазы реполяризации потенциала действия нейрон находится в абсолютно рефрактерном состоянии и не может воспринимать следующие импульсы в результате инактивации потенциалозависимых натриевых каналов. Вслед за этим периодом наступает интервал относительной рефрактерности, во время которого стимул, превышающий исходный более чем на 15 мВ, способен вызвать формирование повторного потенциала действия. Часто для достижения показателя 35 мВ генераторный потенциал производит от 50 до 100 импульсов в секунду (частота составляет от 50 до 100 Гц).
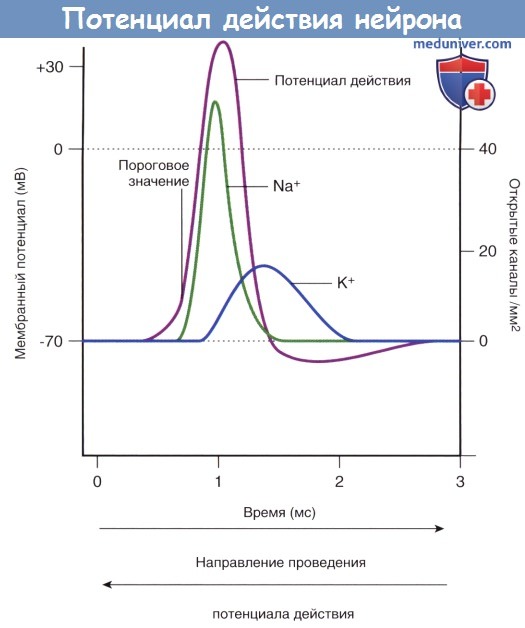
Изменения проницаемости потенциалозависимых натриевых и калиевых каналов, обеспечивающих развитие потенциала действия. 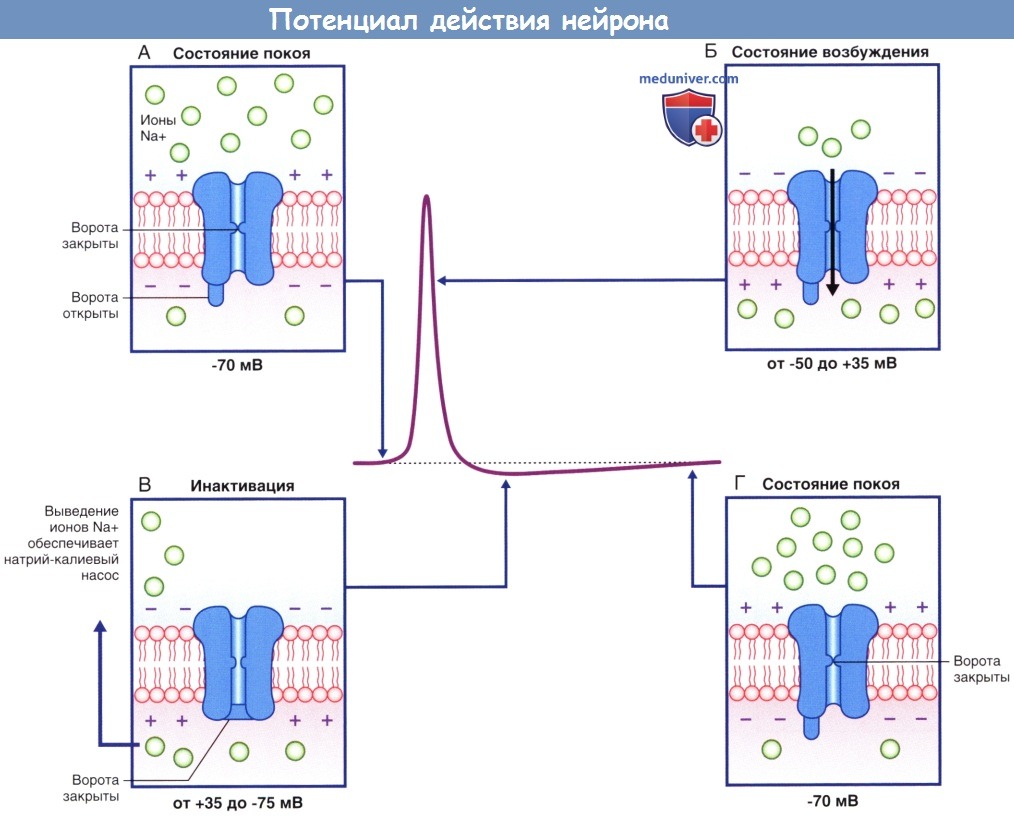
Потенциалозависимые натриевые каналы в разные фазы потенциала действия.
(А) В состоянии покоя, предшествующем возбуждению, активационные ворота находятся в закрытом состоянии, а инактивационные — в открытом.
(Б) При преодолении порога возбуждения происходит открытие активационных ворот, что приводит к полному открытию ионного канала.
(В) Инактивационные ворота закрываются, что приводит к закрытию ионного канала.
(Г) При восстановлении потенциала покоя активационные ворота закрываются, а инактивационные — открываются.
Ионный канал возвращается в исходное состояние. 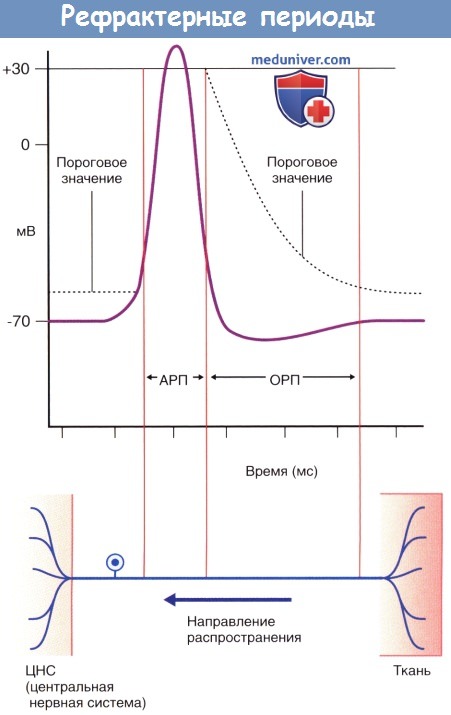
Рефрактерные периоды.
АРП — абсолютный рефрактерный период;
ОРП — относительный рефрактерный период.
в) Передача нервных импульсов. Изменение потенциала триггерной зоны распространяется вдоль аксона в соответствии с электротонической траекторией. Положительные заряды внутренней стороны мембраны распространяются в обоих направлениях через аксоплазму. Положительные заряды внешней стороны мембраны аналогичным образом проходят через внеклеточную жидкость и нейтрализуют отрицательный заряд на внешней стороне мембраны. Прилежащий проксимальный участок мембраны достаточно рефрактерен и способен противостоять деполяризации. Одновременно с этим в прилежащем дистальном сегменте возникает локальный ответ (деполяризация), который достигает пороговых величин и распространяется далее вдоль ствола и ветвей аксона к нервным окончаниям, обеспечивая проведение потенциала действия.
Проведение импульса по немиелинизированному нервному волокну происходит непрерывно, а по миелинизированному — сальтаторно (скачкообразно). Миелиновая оболочка выполняет функцию изолятора и располагается между перехватами Ранвье, где сконцентрированы натриевые каналы. В связи с этим в каждом перехвате Ранвье последовательно возникают потенциалы действия за счет распространения положительного тока вдоль аксоплазмы миелинизированного участка аксона и его выхода в области следующего перехвата Ранвье. При распространении тока в обратном направлении по внеклеточной жидкости для повторного заряда деполяризованного участка мембраны снятие положительного заряда приводит к деполяризации следующего перехвата Ранвье.
г) Скорость проведения нервных импульсов. Скорость проведения нервного импульса по немиелинизированным нервным волокнам прямо пропорциональна диаметру аксона. В первую очередь это связано с тем, что больший объем аксоплазмы обеспечивает более высокую скорость продольного распространения возбуждения. Кроме того, чем больше диаметр аксона, тем больше площадь поверхности его плазматической мембраны, следовательно, на ней расположено большее количество ионных каналов, а деполяризация и восстановление заряда мембраны происходят быстрее. Диаметр аксонов варьирует от 0,2 до 2 мкм, а скорость проведения импульсов — от 2 до 25 м/с.
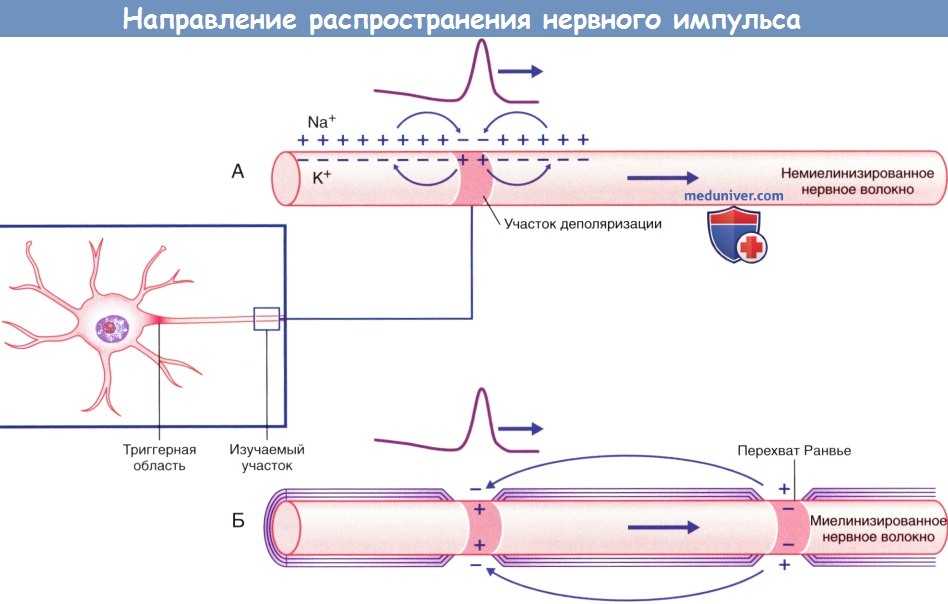
Направление распространения импульса, показанное в виде прохождения положительного заряда.
(А) Непрерывное проведение импульса по немиелинизированному нервному волокну.
(Б) Скачкообразное проведение импульса по миелинизированному нервному волокну.
д) Механизм действия местных анестетиков. При применении местных анестетиков происходит обратимая блокировка проведения нервных импульсов за счет инактивации натриевых каналов, особенно потенциалозависимых, вследствие чего деполяризация мембраны становится невозможной. Молекулы местных анестетиков связывают белок, окружающий ионные каналы. В течение времени действия анестетика происходит связывание белка, окружающего ионные каналы. Для доступа к этим белкам местные анестетики должны быть хорошо растворимы в жирах. При введении анестетика вблизи периферического нерва в первую очередь происходит инактивация немиелинизированных и тонко миелинизированных Аб-волокон. При воздействии на смешанный нерв может возникнуть временный двигательный паралич.
Большинство местных анестетиков — производные амидов (бупивакаин, лидокаин) или сложных эфиров (бензокаин, новокаин). И те, и другие вызывают местное расширение сосудов за счет прямого расслабления гладкой мускулатуры артериол, что приводит к ускорению их выведения. В связи с этим в растворы для местной анестезии часто добавляют адреналин, вызывающий сокращение гладкой мускулатуры и сужение сосудов.
е) Резюме. Электрически заряженные атомы или группы атомов называют ионами. Ионы Na + и К + являются катионами, а ионы Cl + и отрицательно заряженные белки — анионами. Клеточные мембраны представляют собой заряженные конденсаторы с потенциалом покоя величиной -70 мВ.
Неуправляемые натриевые, калиевые и хлорные каналы открыты постоянно, и движение соответствующих ионов через них происходит по градиенту концентрации. Натриевые каналы относительно малочисленны, а калиевые и кальциевые каналы представлены в большом количестве. Ионы К + сконцентрированы в цитозоле за счет притягивающего влияния белков-анионов цитоскелета и отталкивающего действия ионов Na + , расположенных на внешней части плазматической мембраны. Натрий-калиевый насос обеспечивает поддержание мембранного потенциала.
Первоначальная реакция мультиполярного нейрона на возбуждающий стимул заключается в формировании угасающих волн положительного электротонуса. В результате их временной и/или пространственной суммации происходит образование генераторного потенциала в начальном сегменте аксона. При достижении пороговых значений (-55 мВ) потенциалозависимые ионные каналы генерируют потенциал действия, распространяющийся по нервному волокну. Тормозные стимулы вызывают формирование волн отрицательного электротонуса, суммация которых приводит к гиперполяризации мембраны, тем самым отдаляя величины ее потенциала от пороговых значений.
Потенциал действия (спайк) проходит фазу нарастания (деполяризации) от начальных значений до +35 мВ, фазу убывания (реполяризации), в которую возвращается к исходным показателям, и фазу следовой гиперполяризации, в которую понижается до -75 мВ, а затем вновь возвращается к первоначальным значениям. Деполяризация мембраны происходит в результате активации потенциалозависимых натриевых каналов, в ходе которой происходит их стремительное (со скоростью + в клетку и деполяризации мембранного потенциала. При достижении мембранного потенциала +35 мВ происходит закрытие натриевых каналов при помощи инактивационных ворот.
При достижении потенциала действия максимальных значений происходит открытие потенциалозависимых калиевых каналов, и поток ионов Na + в клетку сменяется потоком ионов К + из клетки, что приводит к реполяризации и следовой гиперполяризации мембранного потенциала.
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
Читайте также:
