
Проведение по демиелинизированным нервным волокнам
15. Классификация нервных волокон.Факторы, определяющие скорость проведения возбуждения по аксонам.
Все нервные волокна, во-первых, делятся на миелинизированные (покрытые миелиновой оболочкой) и немиелинизированные (не покрытые миелиновой оболочкой). Также аксоны разделяют по их диаметру и скорости проведения возбуждения.
Скорость проведения возбуждения по нервному волокну зависит от диаметра волокна, от наличия или отсутствия миелиновой оболочки (по миелинизированным аксонам возбуждение проводится быстрее), а также от свойств мембраны нервного волокна (плотности натриевых каналов, емкости мембраны и т.д.).
Классификация нервных волокон по Эрлагеру – Гассеру
Однако по миелинизированным участкам мембраны тоже должна происходить передача возбуждения. В этих участках деполяризация распространяется по мембране пассивным (электротоническим) способом. Внутренняя часть нерва заполнена внутриклеточной средой, которая является достаточно хорошим проводником, т.к. содержит свободные заряженные частицы, в частности положительно заряженные ионы металлов. В результате возникновения на мембране в области перехвата Ранвье зоны возбуждения (ПД) и, соответственно, разности потенциалов на соседних участках мембраны возникают токи заряженных частиц, и эти токи, распространяясь по волокну, несут с собой деполяризацию. Однако эти токи по мере удаления от источника тока (в данном случае им является участок возбужденной мембраны, где наблюдается ПД и перезарядка мембраны) довольно быстро угасают (тем быстрее, чем выше сопротивление внутриклеточной среды), поэтому электротонически возбуждение может распространяться только на очень короткие расстояния. Однако, несмотря на то, что электротон угасает по мере удаления от перехвата Ранвье, на мембрану следующего перехвата все же приходит достаточная деполяризация для того, чтобы возник ПД. Дальше уже этот перехват Ранвье выступает как источник тока, электротон распространяется, угасая, по следующему миелинизированному участку и т.д.
Таким образом, успешное распространение импульса по миелинизированному волокну определяется расстоянием между перехватами Ранвье (они не должны быть слишком большими, чтобы деполяризация не угасла до подпорогового уровня) и амплитудой ПД. Если по какой-то причине амплитуда ПД уменьшается (например, выключается часть натриевых каналов или изменяется состояние мембраны), а все остальное в нерве (сопротивление внутриклеточной среды, расстояние между перехватами Ранвье) остается прежним, то в следующем перехвате Ранвье ПД уже может не возникнуть, т.к. деполяризация мембраны может оказаться ниже пороговой. И, таким образом, дальнейшее проведение импульса по волокну прекратится.
Роль возбудимости в жизнедеятельности клеток гораздо шире, чем просто проведение сигналов на большие расстояния. Она важна для многих общих процессов, протекающих не только в нервных, но и других клетках организма.
Роль возбудимости в жизнедеятельности клеток (по: Шеперд Г., 1987).
| Развитие клеток: оплодотворение клеточное деление морфогенез |
| Перенос ионов через мембрану |
| Биолюминесценция |
| Секреция: гормонов продуктов экзокринных желез медиаторов |
| Движение: ресничек сосудов мышц |
| Передача информации в нервной системе: проведение импульса |
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Поенциал действия может возникнуть в любой точке возбудимой мембраны. Механизм распространения потенциала действия по нервным волокнам зависит от их типа, т.е. немиелинизированный или миелинизированный. Возбуждение способно проводиться, распространяться по мембране нейрона.
Немиелинизированные волокна. При распространении потенциала действия по немиелинизированным волокнам в месте раздражения (электрическое или механическое раздражение мембраны)активируются натриевые каналы, повышается проницаемость мембраны для ионов натрия. Возникают локальные электрические токи, которые вызывают деполяризацию участков мембраны, находящихся в состоянии покоя, т.е. в этих участках мгновенно открываются потенциал-зависимые натриевые каналы. Процесс возбуждения распространяется по всей длине нервного волокна от места возникновения потенциала действия в двух направлениях. Поскольку в фазу деполяризации нервного волокна его возбудимость полностью отсутствует (фаза абсолютной рефрактерности), то потенциал действия не может распространяться в обратном направлении.
Потенциальная амплитуда максимальна – закон "все или ничего", изменение знака с "-" на "+" – принцип овершута; сила ЭДС такова, что вызывает возбуждению на соседних участках мембраны. Дальше этот процесс переходит на соседние участки.
Такое волокно покрыто мембраной швановской клетки (вспомогательные клетки нервной ткани, которые формируются вдоль аксонов периферических нервных волокон. Создают, а иногда и разрушают, электроизолирующую миелиновую оболочку нейронов) на протяжении 1-3 мм. Цитоплазма швановской клетки содержит липид – сфингомиелин, который уменьшает поток ионов через мембрану примерно в 5000 раз и снижает ее емкость в 50 раз. Между швановскими клетками мембрана аксона открыта на расстоянии 2-3 мкм (перехват Ранвье). В зоне перехвата Ранвье мембрана аксона способна генерировать потенциалы действия, которые будут распространяться от перехвата к перехвату, т.е. сальтаторно. Сальтаторный механизм повышает скорость распространения потенциалов действия от 5 до 50 рази экономит энергию нервного волокна на процесс передачи возбуждения. Кроме того, в таких волокнах реполяризация мембраны после потенциала действия происходит с очень высокой скоростью. При этом большинство калиевых каналов остаются закрытыми. Сальтаторное проведение возбуждения осуществляется преимущественно за счет вольт-зависимых натриевых каналов.
Действие порогового раздражителя, возникает возбуждение, следующее возбуждение возникает в перехватах Ранвье, увеличивается скорость возбуждения – миелинизация.
Возбудимость нервного волокна имеет обратную зависимость с диаметром нервного волокна. Скорость проведения возбуждения пропорциональна диаметру нервного волокна. Эволюционно на фоне уменьшения диаметра нервных волокон повышение скорости проведения возбуждения было достигнуто за счет миелинизации волокон и появления сальтаторного механизма проведения нервного импульса.
Законы проведения возбуждения по нервному волокну.
1. Возбуждение проводится в обе стороны.
2. Потенциал действия максимален.
3. Для проведения необходима анатомическая и физиологическая целостность нервного волокна.
Потенциал действия нерва.
В состав нерва входят нервные волокна с разной скоростью проведения возбуждения.
Впервые эту особенность изучили Эрлангер и Гассер. Они предложили свою классификацию. 3 типа нервных волокон:
1. А-тип – миелиновые нервные волокна, скорость= от 50 до 100 м/с; афферентные и эфферентные соматические нервные волокна.
2. В-тип – миелиновые, но меньшего диаметра, скорость от 20 до 40 м/с; преганглионарные нервные волокна ВНС.
3. С-тип – безмиелиновые, скорость 1-2 м/с; постганглионарные нервные волокна.
Законы проведения возбуждения по нервам.
1. Возбуждение проводится в обе стороны.
2. По нерву закон "все или ничего" не выполняется (ПД не максимален).
3. Требуется анатомическая и физиологическая целостность.
4. Нерв обладает капельными свойствами.
Потенциал действия нервного ствола. Если на нерв нанести пороговое раздражение, то ПП быстро уменьшится, и кратковременно (миллисекунда) внутренняя сторона мембраны станет электроположительной относительно наружной. Это кратковременное изменение ПП при возбуждении клетки называется потенциалом действия. ПД в возбудимых тканях возникает при снижении величины ПП (деполяризации мембраны) до так называемого порога генерации ПД, который в среднем равен 15-20 мВ. По своей амплитуде(100-120 мВ0 ПД на 20-25 мВ превышает величину ПП. Это значит, что внутренняя сторона мембраны на короткое время становится электроположительной по отношению к наружной – "овершут" или реверсия заряда. При ПД мембрана избирательно проницаема для ионов натрия. При этом ионы натрия через ионный канал устремляются по концентрационному градиенту внутрь клетки. Это связано с открытием потенциал-зависимых натриевых каналов. При раздражении мембрана деполяризуется, что открывает натриевые каналы и создает поток ионов натрия внутрь клетки. Это дополнительно усиливает деполяризацию, увеличивает число открытых натриевых каналов и повышает натриевую проницаемость. В результате потенциал внутренней стороны мембраны стремится достичь величины натриевого равновесного потенциала. Причиной прекращения роста ПД и начала
реполяризации мембраны клетки являются, во-первых, увеличение деполяризации мембраны, во-вторых, происходит инактивация натриевых каналов в результате деполяризации мембраны и торможение ее натриевой проницаемости; в-третьих, увеличивается проницаемость мембраны для ионов калия в результате все той же деполяризации мембраны. Выходящий калиевый ток стремится сместить мембранный потенциал в сторону калиевого равновесного потенциала. В определенный момент времени величина входящего тока натрия сравнивается с возросшим выходящим током калия – рост амплитуды ПД прекарщается. Когда суммарный выходящий ток превышает входящий, начинается реполяризация мембраны. Реполяризация закрывает натриевые каналы, что снижает натриевую проницаемость мембраны, ускоряет реполяризацию и восстанавливает ПП.
В нервных волокнах сигналы передаются с помощью потенциалов действия, которые представляют собой быстрые изменения мембранного потенциала, быстро распространяющиеся вдоль мембраны нервного волокна. Каждый потенциал действия начинается со стремительного сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу. При проведении нервного сигнала потенциал действия движется вдоль нервного волокна вплоть до его окончания. Нервный ствол, или нерв, представляет собой пучок множества нервных волокон, покрытых у позвоночных общими эпителиальной и соединительнотканной оболочками. При регистрации электрической активности нервного ствола Джозеф Эрлангер и Герберт Гассер в 1937 г. обнаружили составной характер тока действия нервного ствола. На основании полученных данных (диаметр, скорость проведения, функция) разработана классификация, в соответствии с которой нервные волокна подразделены на группы А, В и С.
- Виртуальная лаборатория
- Мочевыводящая система
- Сердечно-сосудистая система
- Влияние давления и вязкости жидкости, а также радиуса и длины сосуда на движение жидкости по сосуду
- Воздействие адреналина, ацетилхолина, атропина и адреналина на основе атропина на артериальное давление
- Воздействие медикаментов и химических медиаторов на деятельность сердца
- Воздействие электрических стимулов на сердечную деятельность
- Воздействие возбуждения блуждающего нерва на сердечную деятельность
- Наложение лигатур Станниуса
- Дыхательная система
- Влияние давления в плевральной полости на вентиляцию легких
- Влияние сурфактанта на вентиляцию легких
- Механизм дыхания. Объемы и емкости легких. Влияние радиуса просвета дыхательных путей
- Внутренняя среда организма. Кровь
- Определение групп крови системы ABO с использованием стандартных сывороток
- Обмен веществ и энергии. Питание
- Физиология человека и животных
- ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА. КРОВЬ. ИММУНИТЕТ
- План. Внутренняя среда организма. Кровь. Иммунитет
- 1. Функции крови. Состав и физико-химические свойства крови. Плазма крови
- 2. Механизмы поддержания кислотно-основного равновесия. Буферные системы крови
- 3. Лимфа. Образование лимфы. Ликвор
- 4. Эритроциты: строение и функции. Гемолиз. Гемоглобин. Эритропоэз. Анемии
- 5. Группы крови. Агглютиногены (антигены) и агглютинины (антитела). Резус-фактор. Правила переливания крови
- 6. Строение и функции лейкоцитов. Лейкоцитарная формула
- 7. Регуляторная функция лейкоцитов (цитокины)
- 8. Защитная функция крови. Понятие о клеточном и гуморальном иммунитете
- 9. Тромбоциты, их строение и функции
- 10. Свертывающая и противосвертывающая системы крови. Плазменные и клеточные факторы свертывания
- ФИЗИОЛОГИЯ СЕРДЕЧНО–СОСУДИСТОЙ СИСТЕМЫ
- План. Физиология сердечно-сосудистой системы
- 1. Строение и функции сердечно-сосудистой системы
- 2. Строение сердца. Общие свойства сердечной мышцы: автоматия, проводимость, возбудимость и сократимость. Типичные (рабочие) и атипичные кардиомиоциты
- 3. Механизм автоматии миокарда. Градиент автоматии
- 4. Особенности кровоснабжения и энергетического обеспечения сердца. Функциональная роль предсердий и желудочков, клапанного аппарата
- 5. Сердечный цикл. Понятие о систолическом и минутном объемах крови
- 6. Методы изучения сердечной деятельности
- 7. Миогенная, нейрогенная и гуморальная регуляция деятельности сердца
- 8. Гемодинамика. Функциональные особенности различных отделов сосудистого русла. Линейная и объемная скорость движения крови. Факторы, обеспечивающие непрерывность кровотока
- 9. Капиллярное кровообращение и его особенности. Микроциркуляция. Представление о тонусе сосудов. Регуляция тонусов сосудов. Сосудодвигательный центр
- 10. Строение лимфатической системы и ее функции. Транспорт лимфы
- ФИЗИОЛОГИЯ ДЫХАНИЯ
- План. Физиология дыхания
- Дыхание у высших позвоночных: внешнее дыхание, газообмен в легких и тканях, транспорт газов кровью, тканевое дыхание
- Вентиляция легких. Механика и динамика дыхательных движений. Внутриплевральное давление и его значение. Роль сурфактанта. Аэрогематический барьер
- Показатели внешнего дыхания. Понятие о легочных объемах и емкостях. Состав вдыхаемого, выдыхаемого и альвеолярного воздуха
- Газообмен в легких и тканях. Парциальное давление кислорода и диоксида углерода в альвеолярном воздухе, венозной и артериальной крови. Транспорт кислорода кровью
- Механизм переноса диоксида углерода, роль карбоангидразы. Взаимосвязь между дыханием и поддержанием кислотно-щелочного равновесия крови
- Нервные механизмы регуляции дыхания. Защитные и регуляторные дыхательные рефлексы. Хеморецепторы. Дыхание при физической нагрузке, при повышенном и пониженном атмосферном давлении
- ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ. ПИТАНИЕ. ТЕРМОРЕГУЛЯЦИЯ
- План. Обмен веществ и энергии. Питание. Терморегуляция
- Обмен веществ и энергии как основное условие гомеостазиса. Анаболические и катаболические процессы. Обмен веществ как источник образования тепла
- Основной и общий обмен. Рабочая прибавка. Общие энергозатраты людей различных профессиональных групп
- Характеристика продуктов питания. Пищевые и питательные вещества. Нормы питания
- Значение воды для организма. Витамины и их роль в обмене веществ
- Принципы составления пищевых рационов
- Терморегуляция. Механизмы теплопродукции и теплоотдачи. Суточные изменения температуры тела у человека
- Лихорадка
- ФИЗИОЛОГИЯ МОЧЕВЫДЕЛЕНИЯ
- План. Физиология мочевыделения
- Строение и функции почек. Нефрон как функциональная единица почки. Особенности кровоснабжения почек
- Механизм образования мочи
- Реабсорбция в нефроне и ее механизмы. Поворотно-противоточная система. Механизмы осмотического концентрирования и разведения мочи
- Процессы секреции и экскреции в почечных канальцах. Образование конечной мочи, ее состав и свойства
- Гуморальная и гормональная регуляции почечной функции
- ФИЗИОЛОГИЯ СИСТЕМЫ ПИЩЕВАРЕНИЯ
- План. Физиология системы пищеварения
- Строение и функции пищеварительной системы. Типы пищеварения
- Пищеварение в ротовой полости. Функции слюнных желез, состав и свойства слюны. Регуляция слюноотделения
- Пищеварение в желудке. Механизмы желудочной секреции ферментов и соляной кислоты. Фазы желудочной секреции. Состав и свойства желудочного сока
- Пищеварение в тонком кишечнике. Пристеночное (мембранное) пищеварение. Всасывание в пищеварительной системе
- Роль печени и поджелудочной железы в пищеварении
- Пищеварение в толстом кишечнике
- Регуляция деятельности органов пищеварения
- Аппетит, голод, насыщение
- ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
- План. Физиология возбудимых тканей
- 1. Раздражимость и возбудимость как способность живых систем реагировать на действие факторов внешней среды. Ионные насосы и их функции (калий-натриевый, кальциевый, хлорный насос)
- 2. Ионные каналы, классификация, строение и функции. Потенциал действия и его фазы. Вклад потенциалзависимых ионных каналов в формирование потенциала действия
- 3. Причины существования относительной и абсолютной рефрактерности. Функциональная лабильность и ее проявления
- 4. Возбудимые клетки как проводники электричества. Распространение электротонического потенциала. Константа длины. Механизм проведения возбуждения по немиелинизированным и миелинизированным нервным волокнам
- 5. Классификация и свойства нервных волокон. Законы проведения возбуждения по нервным волокнам
- 6. Основы межклеточного взаимодействия в организме. Физиология синапсов
- 7. Химические синапсы. Механизм возникновения возбуждающего постсинаптического потенциала. Тормозной постсинаптический потенциал
- 8. Общая характеристика и классификация нейромедиаторов. Нейромодуляторы
- 9. Ионотропные и метаботропные рецепторы. Основные группы синаптически активных веществ (лиганды, агонисты, антагонисты)
- 10. Физиология мышц. Классификация мышц по структурным, биохимическим и функциональным критериям
- 11. Виды и режимы мышечных сокращений. Работа и сила мышц. Типы нервных волокон
- 12. Механизм мышечного сокращения. Роль АТФ, кальция и ионных насосов в мышечном сокращении. Энергетическое обеспечение мышечного сокращения
- ОБЩАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
- План. Общая физиология центральной нервной системы
- 1. Структурно-функциональная организация нейронов и глиальных клеток. Представления о функциональной роли глии
- 2. Строение и свойства нейронов
- 3. Понятие о нервном центре, нервной сети. Свойства нервных центров
- 4. Принципы координации работы нервных центров
- 5. Торможение в ЦНС и его виды. Интегративная деятельность нейрона
- 6. Рефлекторная теория. Компоненты рефлекторной дуги. Классификация рефлексов
- 7. Классификация врожденных форм поведения. Принцип обратной связи
- 8. Теория функциональных систем П.К. Анохина
- ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
- План. Частная физиология центральной нервной системы
- 1. Функциональная организация спинного мозга и его роль в координации рефлекторной деятельности центральной нервной системы
- 2. Функции спинного мозга. Особенности управления движениями на уровне спинного мозга. Спинальный контроль мышечных сокращений и управление мышечным тонусом
- 3. Спинальные соматические и вегетативные рефлексы. Сегментарный и межсегментарный принцип работы спинного мозга
- 4. Структурно-функциональная характеристика основных отделов головного мозга
- 5. Функции продолговатого мозга, топография ядер, регуляция витальных функций
- 6. Роль ретикулярной формации мозгового ствола в регуляции функций. Варолиев мост
- 7. Средний мозг
- 8. Мозжечок: строение, связи и регулирующие влияния
- 9. Промежуточный мозг. Роль ядер таламуса в передаче сигналов с периферии в кору больших полушарий. Гипоталамус как интегративно координирующий вегетативный центр мозга
- 10. Подкорковые ганглии, их строение и функции. Экстрапирамидная двигательная система
- 11. Гиппокамп и лимбическая система, их взаимодействие при интеграции сигналов в мозге
- 12. Физиология вегетативной нервной системы. Современные представления о нейрогуморальной регуляции висцеральных функций организма. Вегетативная рефлекторная дуга
- 13. Функциональные особенности и взаимодействие отделов вегетативной нервной системы в управлении функциями организма
- 14. Основы физиологии коры больших полушарий. Представление о кортикализации функций. Цитоархитектоника коры больших полушарий
- 15. Понятие о полях и зонах коры больших полушарий
- 16. Электрическая активность коры головного мозга. Электроэнцефалограмма
- ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ФУНКЦИЙ
- План. Гормональная регуляция функций
- 1. Эндокринный контроль функций и его регуляторная роль. Участие желез внутренней секреции в интегративной регуляции деятельности организма
- 2. Роль эндокринной системы в регуляции процессов роста, развития, размножения, разных форм адаптации, поведения. Классификация гормонов
- 3. Современные взгляды на механизмы влияния гормонов: рецепторы и вторые посредники
- 4. Гипоталамо-нейрогипофизарная и гипоталамо-аденогипофизарная системы. Гипоталамические рилизинг – факторы (либерины и статины)
- 5. Структура и функции долей гипофиза, секретируемые тропные и эффекторные гормоны, их роль в организме
- 6. Эпифиз и роль мелатонина у животных и человека
- 7. Щитовидная железа. Тиреоидные гормоны (трииодтиронин и тироксин) и кальцитонин
- 8. Паращитовидные железы
- 9. Эндокринная функция поджелудочной железы и ее гормоны (инсулин, глюкагон, соматостатин)
- 10. Гормоны коркового и мозгового слоя надпочечников
- 11. Роль надпочечных желез в реализации адаптационно-приспособительной деятельности организма (стресс). Фазы стресса
- 12. Половые железы и их внутренняя секреция. Гормональная функция семенников. Гормональная функция яичников. Половые циклы
- 13. Эндокринная функция почек, желудочно-кишечного тракта, сердца. Гормональная функция эндотелия
- ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ
- План. Физиология сенсорных систем
- 1. Принципы строения сенсорных систем. Понятие о рецепторах
- 2. Классификация рецепторов. Механизм возбуждения рецепторов. Рецепторный и генераторный потенциалы. Адаптация рецепторов
- 3. Кодирование сенсорной информации. Проводниковые структуры сенсорных систем
- 4. Зрительный анализатор
- 4. I. Преломление света в оптических средах глаза. Построение изображения на сетчатке
- 4. II. Типы движений глаз
- 4. III. Строение сетчатки. Фоторецепторы и преобразование световой энергии в электрический сигнал
- 4. IV. Представления о механизмах цветного зрения и обработке зрительной информации в ЦНС
- 4. V. Проводниковый и корковый отделы зрительного анализатора
- 5. Слуховой анализатор
- 5. I. Строение слухового анализатора
- 5. II. Строение улитки. Орган Корти. Фонорецепторы. Звуковосприятие
- 5. III. Проводящие пути и корковое представительство слухового анализатора. Представления о механизме восприятия частоты и интенсивности звука
- 6. Вестибулярный анализатор
- 7. Вкусовой анализатор: структурные и функциональные особенности. Клеточные механизмы вкусовой рецепции
- 8. Обонятельный анализатор: структурные и функциональные особенности обонятельного анализатора. Клеточные механизмы обонятельной рецепции
- 9. Соматосенсорная система. Кожные рецепторы: тактильные, температурные, болевые. Мышечно-суставная рецепция (проприорецепция)
- 10. Болевые рецепторы. Защитные (ноцицептивные) рефлексы. Современные теории боли
- ФИЗИОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
- План. Физиология высшей нервной деятельности
- 1. Понятие о ВНД. Современные подходы к нейробиологическим механизмам поведения
- 2. Роль эмоций и мотиваций в организации поведения
- 3. Условный рефлекс как приспособительный механизм в животном мире. Классические и инструментальные условные рефлексы. Классификация условных рефлексов
- 4. Нейрофизиологическая сущность торможения. Характеристика внешнего безусловного торможения. Запредельное торможение, его биологическое значение. Основные виды условного торможения
- 5. Нейрофизиологическая сущность сна. Электроэнцефалографическая характеристика медленноволнового сна и парадоксального сна
- 6. Память как свойство центральной нервной системы воспринимать, хранить и воспроизводить информацию
- НЕЙРОГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
- План. Нейрогуморальная регуляция физиологических функций
- 1. Общее понятие о процессах регуляции функций. Гомеостазис. Принципы регуляции функций
- 2. Механизмы регуляции жизнедеятельности организма. Нервная регуляция как высший этап развития приспособлений организма к меняющимся условиям существования
- 3. Единство нервных, гуморальных и иммунных механизмов регуляции
- ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА. КРОВЬ. ИММУНИТЕТ
4. Возбудимые клетки как проводники электричества. Распространение электротонического потенциала. Константа длины. Механизм проведения возбуждения по немиелинизированным и миелинизированным нервным волокнам
Мембрана нейрона и его отростков (нервных волокон) обладает способностью не только генерировать потенциал действия, но и передавать го на относительно большие расстояния без уменьшения его амплитуды и расходования энергии, поэтому нервные импульсы распространяются без затухания (бездекрементно). Способность к распространению возбуждения связана с тем, что во время потенциала действия происходит изменение знака заряда в возбужденном участке мембраны (инверсия заряда). Между ним и невозбужденными соседними участками мембраны возникают локальные электрические токи, под действием которых происходит деполяризация новых соседних участков, что приводит к формированию в них потенциала действия. За счет последовательного охвата возбуждением все новых участков нервного волокна возбуждение перемещается вдоль дендритов и аксонов. Чем больше диаметр волокна, тем больше скорость распространения возбуждения.
Особенности проведения возбуждения в нервных волокнах определяются наличием или отсутствием у них оболочки. Нервные волокна делятся на миелиновые и безмиелиновые, то есть покрытые особой миелиновой оболочкой или не покрытые. Эта оболочка образуется на периферии шванновскими клетками, а в головном мозге – клетками олигодендроглии.
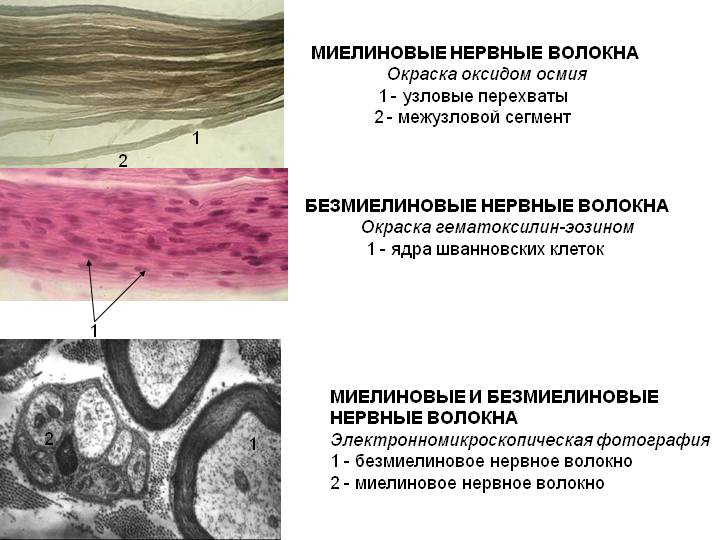

Механизм проведения импульса по нервным волокнам объясняется теорией “местных токов” Тасаки (рисунок 3 и рисунок 4). В безмиелиновом нервном волокне ПД генерируется каждой точкой поверхности мембраны – так называемое непрерывное проведение возбуждения. Во время пикового ПД на наружной поверхности волокна происходит перезарядка, в результате чего возникает местный ток от соседнего положительно заряженного участка к возбужденному, а внутри – наоборот. Таким образом образуется кольцевой ток.
В миелиновых волокнах слой миелина не целиком охватывает волокно, а имеет промежутки – перехваты Ранвье. Сам миелин обладает высоким сопротивлением электрическому току, поэтому в участках, покрытых миелином, потенциал действия генерироваться не может. Перехваты Ранвье располагаются примерно через каждые 0,5-1,5 мм (в зависимости от толщины оболочки), в них обнаружено очень много (около 12 тыс/мкм 2 ) натриевых каналов, поэтому в миелиновых волокнах проведение возбуждения происходит скачкообразно (сальтаторно).
Между соседними участками возникает большой по величине ток (ПД = 100-120 мВ), который в 3-5 раз превышает порог раздражения и способен возбудить соседние участки, что составляет фактор надежности.
Скорость проведения возбуждения пропорциональна корню квадратному от диаметра волокна. Самые тонкие волокна – безмиелиновые, а в миелиновых волокнах чем толще слой миелина, тем больше расстояние между перехватами Ранвье. В безмиелиновых волокнах местные кольцевые токи последовательно распространяются “маленькими шажочками” непрерывно, скорость проведения возбуждения в безмиелиновых волокнах гораздо ниже, чем в миелиновых. Такая передача импульса является медленной и неэкономичной в плане энергозатрат. Поэтому безмиелиновыми являются только самые тонкие волокна (менее 1 мкм в диаметре), проводящие возбуждение к медленно работающим органам – кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.
Константа длины – это расстояние, на котором местный ток сохраняет пороговое значение, то есть способен вызывать образование ПД на следующем участке нервного волокна.
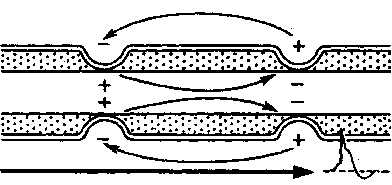
Рисунок 3 – Передача возбуждения в миелинизированном нервном волокне

Рисунок 4 – Передача возбуждения в немиелинизированном нервном волокне
Такая передача импульса является медленной и неэкономичной в плане.
Прямые электрофизиологические исследования, проведенные Като (1924), а затем Тасаки (1953) на одиночных миелинизированных нервных волокнах лягушки, показали, что потенциалы действия в этих волокнах возникают только в перехватах, а участки между, перехватами, покрытые миелином, являются практически невозбудимыми.
В состоянии покоя наружная поверхность возбудимой мембраны всех перехватов (перехватов Ранвье) заряжена положительно. Разности потенциалов между со седними перехватами не существует. В момент возбуждения поверхность мембраны перехвата А становится заряженной электроотрицательно по отношению к поверхности мембраны соседнего перехвата Б. Это приводит к возникновению местного (локального) электрического тока, который идет через окружающую волокно межтканевую жидкость, мембрану и аксоплазму в направлении, показанном на рис. 46 стрелкой. Выходящий через перехват Б ток возбуждает его, вызывая перезарядку мембраны. В перехвате А возбуждение еще продолжается, и он на время становится рефрактерным. Поэтому перехват Б способен привести в состояние возбуждения только следующий пере хват В и т. д.
Время, необходимое для передачи возбуждения от одного перехвата другому, примерно одинаковое у волокон различного диаметра (при температуре 24 °С оно составляет около 0,07 мс). Длина межперехватных участков, как отмечалось, пропорциональна диаметру нераного волокна. Отсюда следует, что в миелинизированных
| 2. | |
| 3 д '-Л | |
| 4 Д | |
| ■J | \ |
| '.Л | |
| ^----------- | |
| ШМ | 1ШШШ |
Рис. 47. Ответы седалищного нерва лягушки на электрические стимулы возрастающей силы.
Отведение потенциалов действия двухфазное. В начале каждой записи виден артефакт раздражения. Сила тока увеличивается постепенно от I к 6. Отметка времени —1 мс.
Рис. 46. Сальтаторное распространение возбуждения в мнкотном нервном волокне от перехвата к перехвату.
Стрелками показано направление тока, возникающего между возбужденным (А) и соседним покоящимся (Б) перехватом.
волокнах скорость проведения нервного импульса примерно пропорциональна их диаметру. В этом отношении миелинизи- рованные волокна отличаются от безмякотных, у которых скорость проведения пропорциональна не диаметру, а корню квадратному из его величины.
Проведение возбуждения по миелинизированному нервному волокну часто сравнивают с передачей сигналов по электрическому кабелю с ретранслирующими генераторами (например, трансатлантическому кабелю). Действительно, участки нервного волокна между перехватами по своим электрическим свойствам подобны кабелю, погруженному в жидкость, обладающую высокой электропроводностью. Внутренним проводником является аксоплазма, внешним — межклеточная жидкость, а изолятором — жировая миелиновая оболочка. Импульс, проходящий между перехватами, представляет собой импульс электрического тока. Перехваты Ранвье играют роль ретранслирующих генераторов, т. е. промежуточных усилительных станций линии связи. При передаче сигнала каждый следующий перехват возбуждается импульсом, генерируемым предыдущим, вырабатывает новый импульс и передает его по волокну. Поскольку сопротивление внутреннего проводника на единицу длины очень велико (в 10 6 раз больше, чем медной проволоки того же диаметра), ретранслирующие генераторы должны быть расположены'близко друг к другу, иначе импульс угаснет.
 |
составной характер потенциала действия нервного ствола и
классификации нервных волокон
Амплитуда электрических импульсов, отводимых от целого нервного ствола, зависит от силы приложенного раздражителя. Раздражителю слабой силы соответствует
 j—1 I L |
100 - 75 -60 -  |
| i I I I i III |
25 50 75 100 125 150 175 200 225 250 275 300
Рис. 48. Комплекс составного потенциала действия смешанного нервного ствола лягушки (по Эрлангеру и Гассеру).
Рис. 49. Схематическоое изображение всех составных частей потенциала действия смешанного нерва (Объяснение в тексте).
Такую картину можно наблюдать, если отводящие электроды расположить на нерве вблизи раздражающих электродов. При увеличении расстояния между этими двумя парами электродов суммарный потенциал действия начинает расчленяться на несколько отдельных колебаний, которые становятся наиболее отчетливо выраженными при удалении отводящих электродов на 10—15 см от места раздражения (рис. 48). Впервые это явление было подробно изучено Эрлангером и Гассером (1937). Они показали, что причиной расчленения суммарного потенциала действия на компоненты является неодинаковая скорость проведения возбуждения по разным волокнам, вследствие чего к отводящим электродам нервные импульсы поступают по этим волокнам неодновременно.
В настоящее время нервные, волокна по скорости проведения возбуждения, длительности различных фаз потенциала действия и строению принято подразделять на три основных типа, обозначаемых буквами А, В и С (рис. 49).
Волокна типа А делятся на четыре подгруппы: ос, (5, у, б. Они покрыты миелиновой оболочкой. Наиболее толстые их них а-волокна (Аа), у теплокровных животных и человека они имеют диаметр 12—22 мкм и характеризуются значительной скоростью проведения возбуждения — 70—120 м/с. Такие волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от определенных рецепторов мышц к соответствующим нервным центрам. Пик потенциала действия волокна Аа у теплокровных длится 0,4—0,5 мс. После его окончания развивается следовая деполяризация, которая продолжается 15—20 мс и переходит в следовую гиперполяризацию длительностью около 40—60 мс.
Три другие группы волокон типа А: Ар, Ау и А5 — имеют меньший диаметр, меньшую скорость проведения и более длительный потенциал действия. Это преимущественно чувствительные волокна, проводящие возбуждение от различных рецепторов (тактильных, некоторых болевых, температурных и рецепторов внутренних органов) в ЦНС.
Исключение составляют лишь у-волокна, значительная часть которых проводит возбуждение в центробежном направлении от клеток спинного мозга к так называемым интрафузальным мышечным волокнам, входящим в состав рецепторов мышц — мышечных веретен (см. рис. 51, А).
К волокнам типа В относятся миелинизированные, преимущественно преганглио- нарные, волокна вегетативной нервной системы. Скорость проведения возбуждения в этих волокнах у теплокровных животных составляет 3—18 м/с. Продолжительность потенциала действия волокон типа В (1—2 мс) примерно в 3 раза превышает длительность потенциала действия волокон типа А. Отличительной особенностью этих волокон является то, что в них не обнаруживается фаза следовой деполяризации: нисходящее колено пика непосредственно переходит в следовую гиперполяризацию, которая в ряде случаев продолжается более 100 мс.
К волокнам типа С относят безмякотные нервные волокна очень малого диаметра (примерно 1 мкм). Скорость проведения возбуждения в этих волокнах не более 3 м/с. Большинство волокон типа С — это постганглионарные волокна симпатической нервной системы.
К волокнам типа С относят также те нервные волокна, которые участвуют в проведении возбуждения от болевых рецепторов и некоторых рецепторов холода, тепла и давления.
Потенциалы действия волокон этого типа характеризуются наибольшей продолжительностью (2 мс у теплокровных животных)., Они имеют длительную фазу следовой деполяризации (50—80 мс), сопровождающуюся еще более продолжительной (300 — 1000 мс) следовой гиперполяризацией. Все эти данные суммированы в табл. 3.
Читайте также:
