
Проведение возбуждения по нерву и от нерва к мышцам
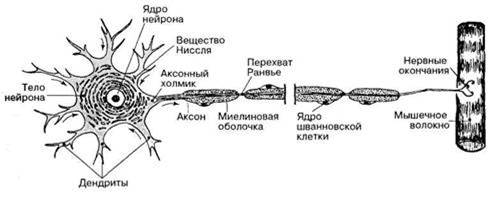
Функцию быстрой передачи возбуждения к нервной клетке и от нее выполняют ее отростки - дендриты и аксоны, т.е. нервные волокна. В зависимости от структуры их делят на мякотные, имеющие миелиновую оболочку, и безмякотные. Эта оболочка формируется шванновскими клетками, являющиеся видоизмененными глиальными клетками. Они содержат миелин, который в основном состоит из липидов. Он выполняет изолирующую и трофическую функции. Одна шванновская клетка образует оболочку на 1 мм нервного волокна. Участки, где оболочка прерывается, т.е. не покрыты миелином называют перехватами Ранвье. Ширина перехвата 1 мкм (рис.).
Функционально все нервные волокна делят на три группы:
1.Волокна типа А - это толстые волокна, имеющие миелиновую оболочку. В эту группу входят 4 подтипа:
1.1.Аa - к ним относятся двигательные волокна скелетных мышц и афферентные нервы, идущие от мышечных веретен (рецепторов растяжения). Скорость проведения по ним максимальна - 70-120 м/сек
1.2.Аb - афферентные волокна, идущие от рецепторов давления и прикосновения кожи. 30 - 70 м/сек
1.3.Аg - эфферентные волокна, идущие к мышечным веретенам (15 - 30 м/сек).
1.4.Аd - афферентные волокна от температурных и болевых рецепторов кожи (12 - 30 м/сек).
2. Волокна группы В - тонкие миелинизированные волокна, являющиеся преганглионарными волокнами вегетативных эфферентных путей. Скорость проведения - 3-18 м/сек
3.Волокна группы С, безмиелиновые постганглионарные волокна вегетативной нервной системы. Скорость 0,5 -3 м/сек.
Проведение возбуждения по нервам подчиняется следующим законам:
1.Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезке, вторая - действии веществ блокирующих проведение, например новокаина.
2.Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаще всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным - от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное или антидромное распространение возбуждения.
3.Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола.
4.Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются проходя по ним.
5.Скорость проведения прямо пропорциональна диаметру нерва.
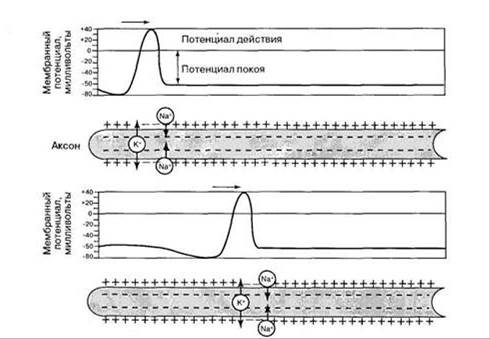
Нервные волокна обладают свойствами электрического кабеля, у которого не очень хорошая изоляция. В основе механизма проведения возбуждения лежит возникновение местных токов. В результате генерации ПД в аксонном холмике и реверсии мембранного потенциала, мембрана аксона приобретает противоположный заряд. Снаружи она становится отрицательной, внутри положительной. Мембрана нижележащего, невозбужденного участка аксона заряжена противоположным образом. Поэтому между этими участками, по наружной и внутренней поверхностям мембраны начинают проходить местные токи. Эти токи деполяризуют мембрану нижележащего невозбужденного участка нерва до критического уровня и в нем также генерируется ПД. Затем процесс повторяется и возбуждается более отдаленный участок нерва и т.д. (рис.). Т.к. по мембране безмякотного волокна местные токи текут не прерываясь, поэтому такое проведение называется непрерывным. При непрерывном проведении местные токи захватывают большую поверхность волокна, поэтому им требуется. длительное время для прохождения по участку волокна. В результате дальность и скорость проведения возбуждения по безмякотным волокнам небольшая.
В мякотных волокнах, участки покрытые миелином обладают большим электрическим сопротивлением. Поэтому непрерывное проведение ПД невозможно. При генерации ПД местные токи текут лишь между соседними перехватами. По закону "все или ничего" возбуждается ближайший к аксонному холмику перехват Ранвье, затем соседний нижележащий перехват и т.д. (рис.). Такое проведение называется сальтаторным (прыжком). При этом механизме ослабления местных токов не происходит и нервные импульсы распространяются на большое расстояние и с большой скоростью.
| | | следующая лекция ==> | |
| Физиология гладких мышц | | | Синаптическая передача |
Дата добавления: 2017-11-04 ; просмотров: 540 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Возбуждение — это очень сложный биологический процесс, который лежит в основе деятельности органов, тканей и клеток организм.а. Возбуждаясь, каждый орган выполняет свою специфическую функцию: например, железы пищеварительного тракта вырабатывают ферменты, железы внутренней секреции-—гормоны, мышцы производят сократительный акт. Наряду с такими совершенно различными специфическими реакциями имеются и общие черты в возбуждении различных органов. Это относится прежде всего к электрическим явлениям — первым и основным процессам, в которых проявляется возбуждение.
Электрические свойства нервных и мышечных клеток в состоянии покоя. Мембранный потенциал. В покое клетка имеет определенный электрический заряд. Снаружи сарколемма заряжена положительно, а изнутри — отрицательно. Возникновение этого двойного электрического заряда связано с особыми свойствами мембраны. Она обладает избирательной проницаемостью для различных ионов. Так, она относительно легко пропускает положительно заряженные ионы (катионы) калия (К+) и почти не пропускает катионы натрия (Nа+). Не могут пройти через мембрану и крупные молекулы белковых анионов. Если бы она была проницаема для всех этих веществ, то содержание их внутри и вне клетки стало бы одинаковым. В связи с избирательной и ограниченной проницаемостью мембраны поддерживается разная концентрация различных ионов внутри клетки и в окружающей ее среде — в межтканевой жидкости. Калия внутри клетки содержится в 30—40 раз больше, чем снаружи, а натрия —в 10—12 раз меньше. В силу разности концентраций катионы К + выходят из клетки наружу (мембрана для них проницаема), ионы же Nа + проникнуть внутрь не могут (мембрана в состоянии покоя для них почти непроницаема). Не могут выйти наружу через мембрану и анионы. В связи с диффузией К+ наружу и под влиянием электростатических сил притяжения между противоположно заряженными ионами анионы концентрируются у поверхности мембраны изнутри, а катионы — снаружи, образуя, таким образом, на мембране двойной электрический слой, т. е. поляризуя ее. Разность потенциалов между наружной и внутренней поверхностями мембраны, называемая потенциалом покоя (ПП), или мембранным потенциалом (МП), равна 70—90 мв.
Изменения электрического состояния клетки при возбуждении. Потенциал действия. При раздражении клетки происходит изменение мембранного потенциала покоя. Возбужденный участок мембраны оказывается снаружи заряженным отрицательно по отношению к своей внутренней поверхности. Иными словами, происходит перезарядка мембраны, смена знаков зарядов — инверсия потенциала покоя. Этот процесс обусловлен изменениями проницаемости мембраны под влиянием раздражения. Она на какое-то короткое время становится значительно более проницаемой для ионов Na+, чем для ионов К + . Ионы Na + , которых в межтканевой жидкости, как уже указывалось, в 10—12 раз больше, чем внутри клетки, начинают проникать внутрь. Нейтрализуя отрицательный заряд внутренней поверхно-сти мембраны в раздражаемом участке, они снижают тем самым существовавшую в покое разность потенциалов, т. е. приводят к деполяризации мембраны. Интересно, что этот процесс сам себя подкрепляет: начинающаяся деполяризация повышает проницаемость мембраны для ионов Nа + ; диффузия катионов Na + углубляет деполяризацию; в связи с этим проницаемость для этих ионов становится еще более значительной и т. д. В результате этого процесса происходит не только деполяризация мембраны, но и ее перезарядка: внутренняя поверхность ее в раздражаемом участке становится заряженной положительно, а наружная по отношению к ней — отрицательно. При измерении разности потенциалов между наружной и внутренней поверхностями клетки оказывается, что вместо заряда — 90 мв внутри, который отмечался в состоянии покоя, там обнаруживается заряд + 30— 40 мв. Проникновение положительно заряженных ионов Na + внутрь клетки привело к развитию электрического процесса, характеризующегося напряжением 120—130 мв <от—90 до +30 мв). Этот процесс — колебание потенциала покоя — получил название потенциала действия (ПД). Он характеризует возникновение возбуждения в нервной или мышечной клетке.
Проведение возбуждения по нервным и мышечным волокнам. С потенциалом действия связано проведение возбуждения по нервным и мышечным волокнам. При возникновении потенциала действия между возбужденным участком и соседними, находящимися в состоянии покоя, возникает разность потенциалов. Наружная поверхность возбужденного участка мембраны, как отмечалось выше, оказывается отрицательно заряженной, а соседнего с ним — находящегося в состоянии покоя — положительно заряженной. В связи с разностью потенциалов между этими соседними участками возникает электрический ток — так называемый местный ток действия. Этот ток является раздражителем участка волокна, находившегося до этого времени в состоянии покоя. Под влиянием раздражения в этом участке начинаются описанные выше процессы — деполяризация, повышение натриевой проницаемости и т. д., т. е. возникает потенциал действия. Затем возбуждается следующий участок волокна и т. д. Таким образом, проведение импульса заключается, по существу, в последовательном, один за другим, возбуждении участков волокна.
Передача возбуждения через синапсы происходит, как уже говорилось выше, через посредство химических веществ — медиаторов, вырабатываемых концевыми веточками аксонов. Химическим путем передается возбуждение как в синапсах центральной нервной системы, где разветвления одних аксонов образуют синапсы на теле и дендритах других, так и в нервномышечном, или мионевральном, синапсе. Медиатором окончаний мотонейрона в мышечных волокнах является ацетилхолин. При возбуждении нервных окончаний синаптические пузырьки, в которых содержится медиатор, лопаются, ацетилхолин проникает через пресинаптическую мембрану в синаптическую. щель и вызывает возбуждение постсинаптической мембраны. Последняя обладает высокой чувствительностью к этому воздействию. Под влиянием ацетилхолина повышается ее проницаемость для ионов Na+ и К + , происходит деполяризация и возникает постсинаптический потенциал. С постсинаптической мембраны возбуждение передается на другие (внесинаптические) участки мембраны мышечного волокна снова электрическим путем (см. рис. 4).
6. Значение функционального состояния нервно-мышечного аппарата для развития процесса возбуждения. Возбудимость. Способность живой ткани развивать возбуждение в ответ на раздражение называется возбудимостью. Таким образом, возбудимость является одним из основных свойств живой ткани, обеспечивающих взаимодействие организма со средой. Разные ткани обладают различной возбудимостью. Уровень возбудимости одной и той же ткани тоже изменчив. Умеренные воздействия на ткань повышают ее возбудимость, чрезмерные по силе или длительности понижают. Так, под влиянием разминки возбудимость центральной нервной, системы и нервно-мышечного аппарата повышается, при утомительной работе понижается.
Изменение возбудимости происходит закономерно во время протекания каждой волны возбуждения. Когда возникает потенциал действия (в течение фазы деполяризации), ткань становится невозбудимой: она не способна ответить на новое раздражение. Это так называемая абсолютная рефракторная фаза. Постепенно возбудимость ткани восстанавливается до исходного уровня, а затем становится на некоторое время даже выше его.
Возбудимость можно измерять. Чем выше возбудимость ткани, тем легче вызвать ее возбуждение — ответную реакцию. Минимальная сила раздражения, которая нужна, чтобы вызвать возбуждение ткани, характеризует так называемый порог возбудимости данной ткани и называется пороговой силой. Уровень возбудимости служит важным показателем функционального состояния ткани.
Функциональная подвижность (лабильность). Одним из важных факторов, от которых зависит деятельность возбудимых тканей (таких, как нервные клетки, синапсы, нервно-мышечный аппарат), является скорость протекания возбуждения, получившая название лабильности (Н. Е. Введенский). В одних образованиях волна возбуждения развивается и затухает с большой скоростью, в других значительно медленнее. От скорости возбуждения зависит частота импульсов, которую ткань может развить в единицу времени. Наиболее высока лабильность нервных волокон, значительно ниже лабильность мышечных волокон, нервных клеток и особенно синапсов.
Лабильность, как и возбудимость ткани, не постоянна. Умеренные воздействия увеличивают скорость протекания волны возбуждения, чрезмерные — ее уменьшают. Под влиянием разминки, например, лабильность центрально-нервных образований и нервно-мышечного аппарата повышается, при утомлении понижается.
Об уровне лабильности можно судить по разным показателям. Н.Е.Введенский предложил измерять ее максимальным числом волн возбуждения, которое может возникнуть в ткани в единицу времени (в 1 сек).
Работа мышц. В процессе мышечного сокращения потенциальная химическая энергия переходит в потенциальную механическую энергию напряжения и кинетическую энергию движения. Различают внутреннюю и внешнюю работу. Внутренняя работа связана с трением в мышечном волокне при его сокращении. Внешняя работа проявляется при перемещении собственного тела, груза, отдельных частей организма (динамическая работа) в пространстве. Она характеризуется коэффициентом полезного действия (КПД) мышечной системы, т.е. отношением производимой работы к общим энергетическим затратам (для мышц человека кпд составляет 15—20%, у физически развитых тренированных людей этот показатель несколько выше).
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Нерву присущи два физиологических свойства - возбудимость и проводимость, т. е. способность на раздражение отвечать возбуждением и проводить его. Проведение возбуждения является единственной функцией нервов. От рецепторов они проводят возбуждение к центральной нервной системе, а от нее - к рабочим органам.
С физической точки зрения нерв очень плохой проводник. Его сопротивление в 100 млн. раз больше, чем у медной проволоки того же диаметра, однако нерв отлично выполняет свою функцию, проводя импульсы без затухания на большое расстояние.
Как осуществляется проведение нервного импульса?
Согласно мембранной теории, каждый возбужденный участок приобретает отрицательный заряд, а так как соседний невозбужденный участок имеет положительный заряд, то два участка оказываются противоположно заряженными. При создавшихся условиях между ними потечет электрический ток. Этот местный ток является раздражителем для покоящегося участка, он вызывает его возбуждение и изменяет заряд на отрицательный. Как только это произойдет, между вновь возбужденным и соседним покоящимся участками потечет электрический ток и все повторится.
Так распространяется возбуждение в тонких, безмиелиновых нервных волокнах. Там, где есть миелиновая оболочка, возбуждение может возникать только в узлах нервного волокна (перехватах Ранвье), т. е. в точках, где волокно оголено. Поэтому в миелиновых волокнах возбуждение распространяется скачками от одного перехвата к другому и движется гораздо быстрее, чем в тонких безмиелиновых волокнах (рис. 16).

Рис. 16. Проведение возбуждения в миелиновом нервном волокне. Стрелками показано направление тока, возникающего между возбужденным (А) и соседним покоящимся (Б) перехватами
Следовательно, в каждом участке волокна возбуждение генерируется заново и распространяется не электрический ток, а возбуждение. Этим объясняется способность нерва проводить импульс без затухания (без декремента). Нервный импульс остается постоянным по величине в начале и в конце своего пути и распространяется с постоянной скоростью. Кроме того, все импульсы, которые проходят по нерву, совершенно одинаковы по величине и не отражают качества раздражения. Меняться может только их частота, которая зависит от силы раздражителя.
Величина и длительность импульса возбуждения определяются свойствами нервного волокна, по которому оно распространяется.
Скорость проведения импульса зависит от диаметра волокна: чем оно толще, тем быстрее распространяется возбуждение. Наибольшей скоростью проведения (до 120 м/с) отличаются миелиновые двигательные и чувствительные волокна, управляющие функцией скелетных мышц, поддерживающих равновесие тела и выполняющие быстрые рефлекторные движения. Наиболее медленно (0,5 - 15 м/с) проводят импульсы безмиелиновые волокна, иннервирующие внутренние органы, и некоторые тонкие чувствительные волокна.
Законы проведения возбуждения по нерву
Доказательством того, что проведение по нерву - процесс физиологический, а не физический, служит опыт с перевязкой нерва. Если нерв туго перетянуть лигатурой, то проведение возбуждения прекращается - закон физиологической целостности.
Нервное волокно может проводить импульс в обе стороны. Если нерв раздражать посредине, то на двух его концах осциллограф зарегистрирует потенциалы действия - закон двустороннего проведения возбуждения.
В нервах импульсы проводятся по отдельным нервным волокнам изолированно. Поэтому один и тот же нерв в одном направлении проводит чувствительные (афферентные), а в другом - двигательные (эфферентные) импульсы - закон изолированного проведения.
Основы эмбриологии человека
Зародыш (эмбрион) - это организм, развивающийся под покровом яйцевых оболочек или внутри материнского организма. Под зародышевым, или эмбриональным, развитием понимают ранний период индивидуального развития - от момента оплодотворения (зачатия) до рождения или вылупления из яйцевых оболочек.
У человека внутриутробный период длится в среднем 280 дней, или 10 лунных месяцев. В акушерской практике зародышем (эмбрионом) называют развившийся организм в течение первых 2 мес внутриутробной жизни, а с III по X месяц - плодом; этот период развития называют плодным, или фетальным.

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим.
ФУНКЦИЮ быстрой передачи возбуждения к нервной клетке и от нее выполняют ее отростки - дендриты и аксоны, т.е нервные волокна. В зависимости от структуры их делят на мякотные, имеющие миелиновую оболочку, и безмякотные. Эта оболочка формируется шванновскими клетками, являющиеся видоизмененными глиальными клетками. Они содержат миелин, который в основном состоит из липидов. Он выполняет изолирующую и трофическую функции. Одна шванновская клетка образует оболочку на 1 мм нервного волокна. Участки, где оболочка прерывается, т.е. 1:е покрыты миелином называют перехватами Ранвье. Ширина перехвата 1 мкм (.рис.). Функционально все нервные волокна делят на три группы: Скорость передачи нервных импульсов по миелинизированным волокнам значительно превышает таковую по волокнам лишенным миелиновой оболочки.
1 Волокна типа А - это толстые волокна, имеющие миелиновую оболочку. В эту группу входят 4 подтипа: двигательные волокна скелетных мышц и афферентные нервы (рецепторов растяжения). Скорость проведения по ним максимальна - 70-120 м,'сек
2 ß (бета) - афферентные волокна, идущие от рецепторов давления и прикосновения кожи. 30-70 м/сек
3 (гамма)- эфферентные волокна, идущие к мышечным веретенам (15-30 м/сек).
4 (сигма) - афферентные волокна от температурных и болевых рецепторов кожи (12-30 м/сек).
Волокна группы В - тонкие миелинизированные волокна, являющиеся преганглионарными волокнами вегетативных эфферентных путей. Скорость проведения - 3-18 м/сек
Волокна группы С, безмиелиновые постганглионарные волокна вегетативной нервной системы. Скорость 0,5 –3 м сек. проведение возбуждения по нервам подчиняется следующим законам:
1. Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезе, вторая - действии веществ блокирующих проведение, например новокаина.
2. Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаше всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным - от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное или антидромное распространение возбуждения.
3. Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола.
4. Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания.
Следовательно, нервные импульсы не ослабляются, проходя по ним.
5. Скорость проведения прямо-пропорциональна диаметру нерва. (Нервные волокна обладают свойствами центрического кабеля, у которого не очень хорошая изоляция. В основе механизма проведения возбуждения лежит возникновение местных токов: В результате генерации ПД в аксоном холмике и реверсии мембранного потенциала, мембрана аксона приобретает противоположный заряд. Снаружи она становится отрицательной, ^внутри Положительной. Мембрана нижележащего, невозбужденного участка аксона заряжена противоположным образом. ''Поэтому между этими участками, по наружной и внутренней поверхностям мембраны начинают проходить местные, токи. Эти токи деполяризуют мембрану нижележащего невозбужденного участка нерва до критического уровня к в нем также генерируется ПД. Затем процесс повторяется и возбуждается более отдаленный участок нерва и т.д. (рис.). Т.к. по мембране безмякотного волокна местные токи текут не прерываясь, поэтому такое проведение называется непрерывным. При непрерывном проведении местные токи захватывают большую поверхность волокна, поэтому им " требуется, длительное время для прохождения по участку волокна. В результате дальность, и. скорость проведения возбуждения по безмякотным волокнам небольшая. В мякотных волокнах участки, покрытые миелином обладают большим электрическим сопротивлением. Поэтому непрерывное проведение ПД Невозможно. При генерации ПД местные токи текут лишь между соседними, перехватами По закону "все или ничего" возбуждается ближайший к аксонному холмику перехват Ранвье, затем соседний нижележащий перехват и т.д. (рис.). Такое проведение называется сальтаторным (прыжком). При этом
механизме ослабления местных токов не происходит, и нервные импульсы распространяются на большое расстояние и с большой скоростью.
Нервные центры

Аванесян
Свойства нервных центров
Нервным центром (НЦ) называется совокупность нейронов в различных отделах ЦНС, обеспечивающих регуляцию какой-либо функции организма. Например, бульбарный дыхательный центр.
Для проведения возбуждения через нервные центры характерны следующие, особенности:
1. однострочное проведение, оно идет от афферентного, через вставочный к эфферентному нейрону. Это обусловлено наличием межнейронных синапсов.
2. Центральная задержка проведения возбуждения т.е по НЦ возбуждения идет значительно медленнее, чем по нервному волокну. Это объясняется синаптической задержкой т.к больше всего синапсов в центральном звене рефлекторной дуги, там скорость проведения наименьшая. Исходя из этого, время рефлекса - это время от начала воздействия раздражителя до появления ответной реакции. Чем длительнее центральная задержка, тем больше время рефлекса. Вместе с тем оно зависит от силы раздражителя. Чем она больше, тем время рефлекса короче и наоборот. Эго объясняется явлением суммации возбуждений в синапсах. Кроме того, оно определяется и функциональным состоянием ЦНС. Например, при утомлении НЦ длительность рефлекторной реакции увеличивается.
3. Пространственная и временная суммация. Временная суммация возникает, как и в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда ВПСП. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторов нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются 11 и мембране нейрона генерируется распространяющийся ПД.
4. Трансформация ритма возбуждения - изменение частоты нервных импульсов при прохождении через нервный центр. Частота может понижаться или повышаться. Например, повышающая трансформация (увеличение частоты) обусловлено дисперсией и мультипликацией возбуждения в нейронах. Первое явление возникает в результате разделения нервных импульсов на несколько нейронов, аксоны которых образуют затем синапсы на одном нейроне. Второе, генерацией нескольких нервных импульсов при развитии возбуждающего постсинаптического потенциала на мембране одного нейрона. Понижающая трансформация объясняется суммацией нескольких ВПСП и возникновением одного ПД в нейроне.
5. Посттетаническая потенциация, это усиление рефлекторной реакции в результате длительного возбуждения
нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы, выделяется большое количество нейромедиатора в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
6. Последействие- это запаздывание окончания рефлекторного ответа после прекращения действия раздражителя. Связано с циркуляцией нервных импульсов по замкнутым цепям нейронов.
7. Тонус нервных центров - состояние постоянной повышенной активности. Он обусловлен постоянным поступлением к НЦ нервных импульсов от периферических рецепторов, возбуждающим влиянием на нейроны продуктов метаболизма и других гуморальных факторов. Например, проявлением тонуса соответствующих центров является тонус определенной группы мышц.
8. автоматия или спонтанная активность нервных центров. Периодическая или постоянная генерация нейронами нервных ИМПУЛЬСОВ, которые возникают в них самопроизвольно, т.е. в отсутствии сигналов от других нейронов или рецепторов. Обусловлена колебаниями процессор метаболизма в нейронах и действием на них гуморальных факторов.
9. Пластичность нервных центров. Это их способность изменять функциональные свойства. При этом центр приобретает возможность выполнять новые функции или восстанавливать старые после повреждения. В основе пластичности Н.Ц. лежит пластичность синапсов и мембран нейронов, которые могут изменять свою молекулярную структуру.
10. Низкая физиологическая лабильность и быстрая утомляемость.
Н.Ц. могут проводить импульсы лишь ограниченной частоты. Их утомление объясняется утомлением синапсов и ухудшением метаболизма нейронов. Утомляемость нервных центров - постепенное снижение и полное прекращение ответа при продолжительном раздражении афферентных нервных волокон. Утомление нервных центров вызывается прежде всего нарушением проведения возбуждения в межнейронных синапсах. Тот факт, что утомление сначала возникает в синапсе доказывается простым опытом. В то время, как раздражение афферентного нервного волокна спинальной лягушки не вызывает сокращения мышцы, стимуляция эфферентного волокна приводит к мышечной реакции.
В настоящее время считают, что утомление синапса обусловлено резким снижением запаса медиатора в пресинаптической мембране (истощение), уменьшением чувствительности постсинаптической мембраны (десенсетизация) и уменьшением энергетических ресурсов нейрона. Не все рефлекторные реакции одинаково быстро приводят к развитию утомления. Некоторые рефлексы могут в течение длительного времени протекать без развития утомления. К таким рефлексам относятся проприорецептивные тонические рефлексы
Простейшим нервным центром является нервная цепь, состоящая из трех последовательно соединенных нейронов (рис). Нейроны сложных нервных центров имеют многочисленные связи между собой, образуя нервные сети трех типов: иерархические локальные и дивергентные с одним входом
ИВАНЬКО
Иерархические сети.
Наиболее распространенный тип межнейронных связей можно увидеть в главных сенсорных и двигательных путях.
В сенсорных системах иерархическая организация носит восходящий характер; в нее включаются различные клеточные уровни, по которым информация поступает в высшие центры – от первичных рецепторов к вторичным вставочным нейронам, затем к третичным и т.д.
Двигательные системы организованы по принципу нисходящей иерархии, где команды “спускаются” от нервной системы к мышцам: клетки, расположенные, фигурально говоря, “наверху”, передают информацию специфическим моторным клеткам спинного мозга, а те в свою очередь – определенным группам мышечных клеток.
Ниже лежащий текст прочитать и рассказать о том, как организуются нервные сплетения и что они снабжают на примере шейного узла.




Галетина
Последнее изменение этой страницы: 2016-09-19; Нарушение авторского права страницы
Читайте также:
